
Assessment of Pb(II), Cd(II), and Al(III) Removal Capacity of Bacteria from Food and Gut Ecological Niches: Insights into Biodiversity to Limit Intestinal Biodisponibility of Toxic Metals

 , , , and
, , , and 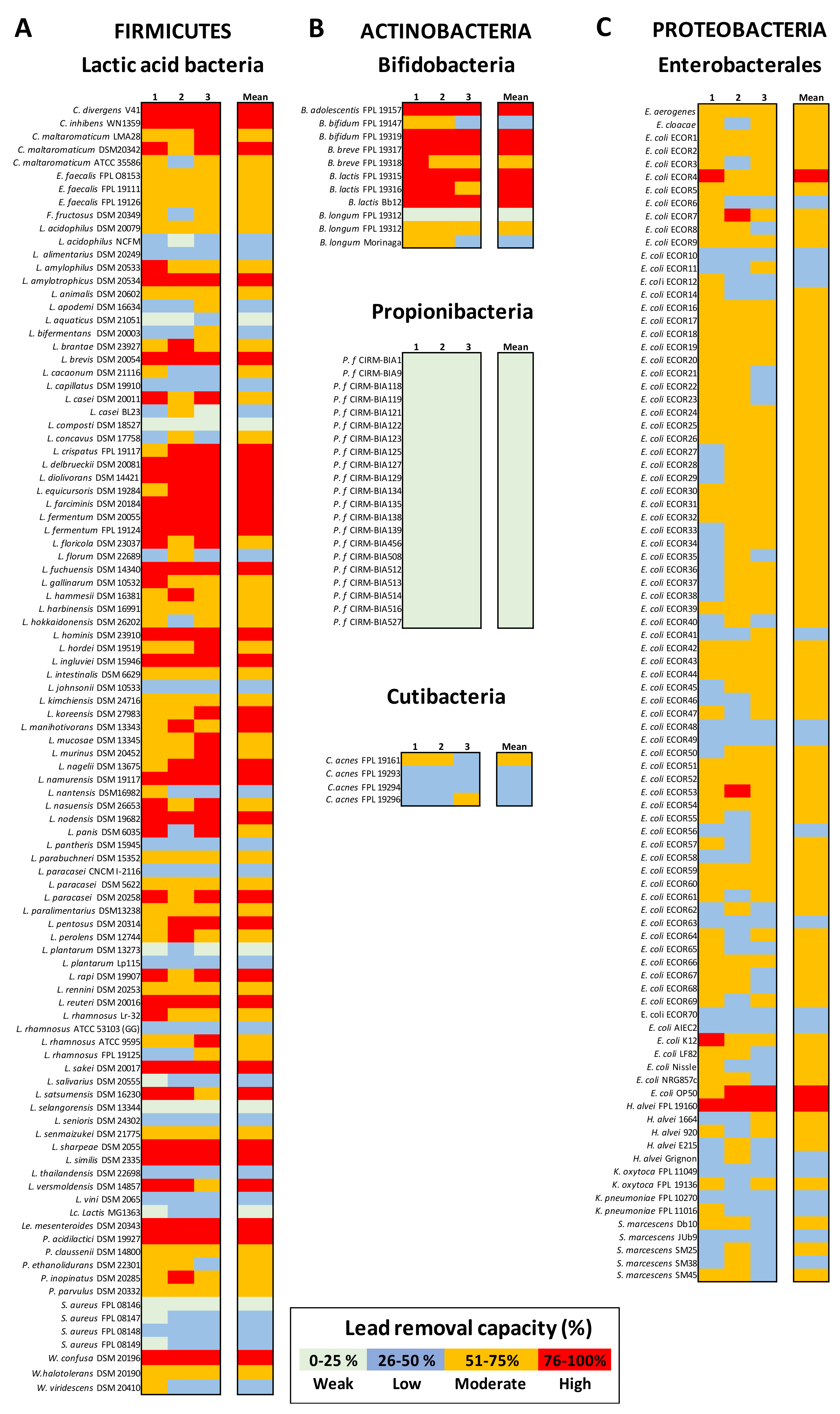{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Reagents and Instruments
2.2. Bacterial Strains Collections and Culture Conditions
2.3. Metal Removal Assays
2.4. Statistical Analyses
3. Results
3.1. Lactic Acid Bacteria (LAB) Exhibit Variable Pb Removal Capacities
3.2. Actinobacteria Strains Differ in Their Pb Biosorption Potential
3.3. Enterobacterales Have a Moderate Pb Biosorption Potential
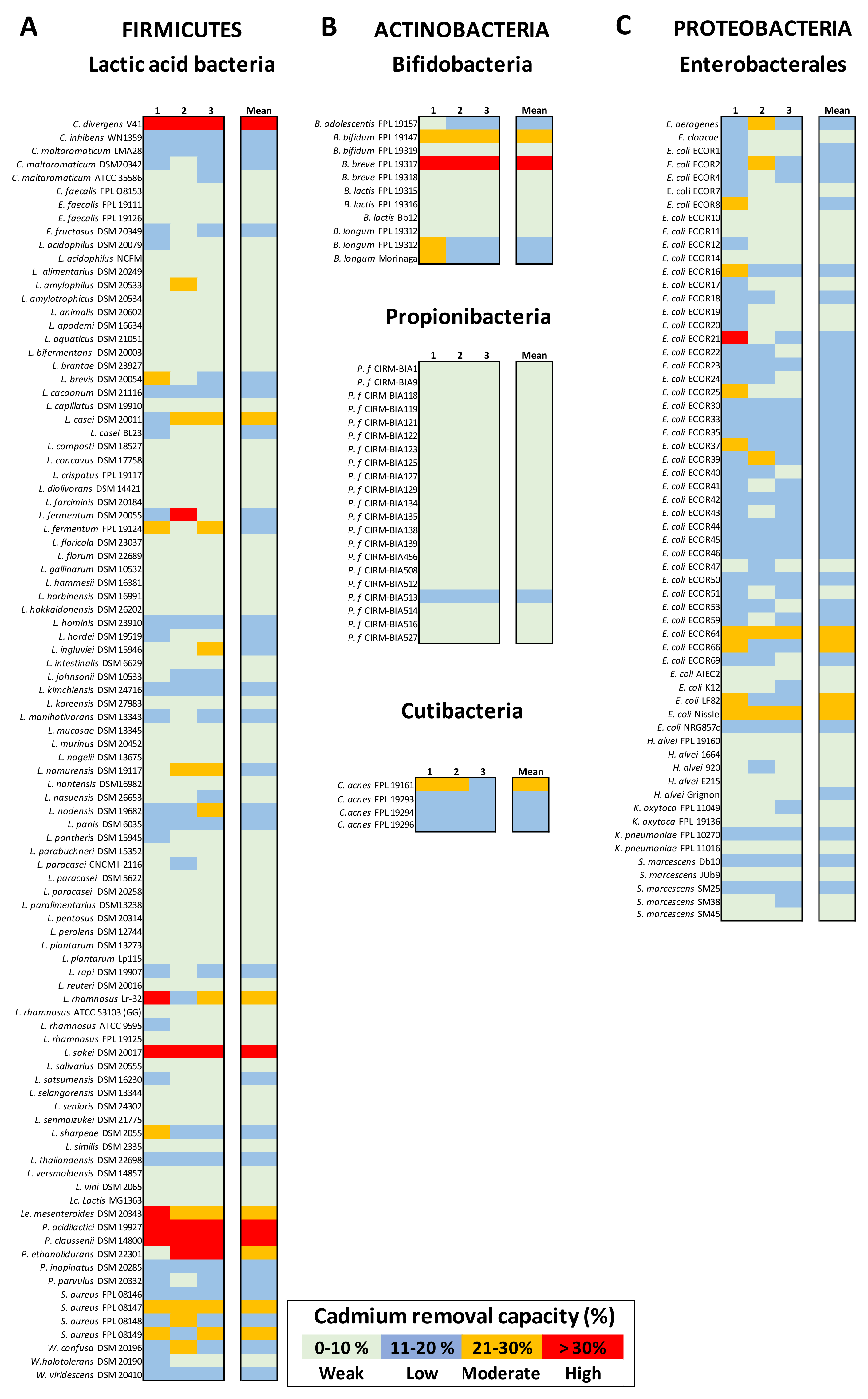
3.4. Bacterium-Mediated Cd Removal Capacity Is Phylum-, Genus-, and Strain-Specific
3.5. Bacteria-Mediated Al Removal Capacity Is Also Genus- and Strain-Dependent
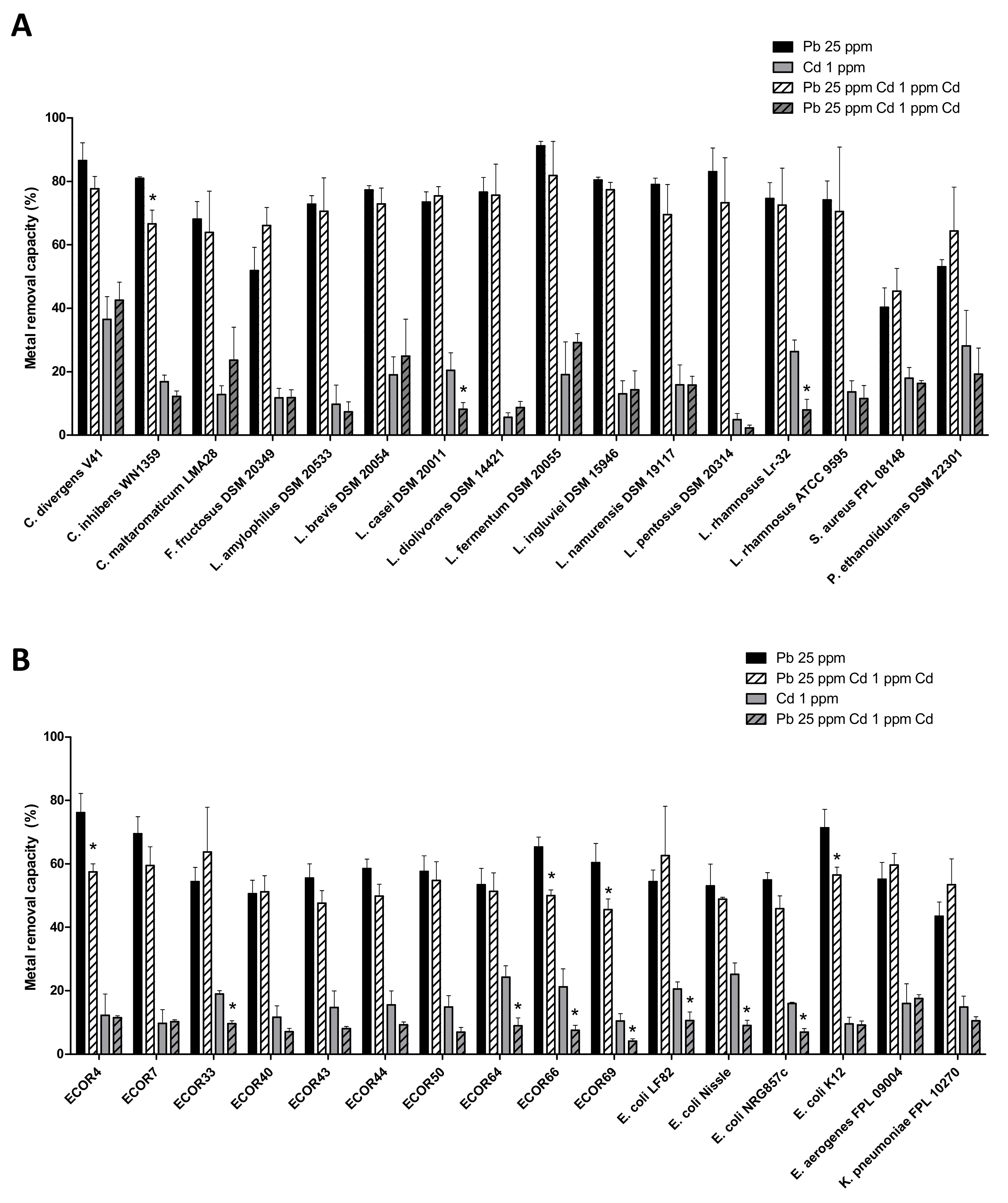
3.6. The Ability of Bacteria to Remove Pb and Cd Is Not Greatly Affected When Both Metals Are Present
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Breton, J.; Le Clère, K.; Daniel, C.; Sauty, M.; Nakab, L.; Chassat, T.; Dewulf, J.; Penet, S.; Carnoy, C.; Thomas, P.; et al. Chronic Ingestion of Cadmium and Lead Alters the Bioavailability of Essential and Heavy Metals, Gene Expression Pathways and Genotoxicity in Mouse Intestine. Arch. Toxicol. 2013, 87, 1787–1795. [Google Scholar] [CrossRef]
- Breton, J.; Daniel, C.; Dewulf, J.; Pothion, S.; Froux, N.; Sauty, M.; Thomas, P.; Pot, B.; Foligné, B. Gut Microbiota Limits Heavy Metals Burden Caused by Chronic Oral Exposure. Toxicol. Lett. 2013, 222, 132–138. [Google Scholar] [CrossRef]
- Flora, G.; Gupta, D.; Tiwari, A. Toxicity of Lead: A Review with Recent Updates. Interdiscip. Toxicol. 2012, 5, 47–58. [Google Scholar] [CrossRef]
- Wani, A.L.; Ara, A.; Usmani, J.A. Lead Toxicity: A Review. Interdiscip. Toxicol. 2015, 8, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Lanphear, B.P.; Hornung, R.; Khoury, J.; Yolton, K.; Baghurst, P.; Bellinger, D.C.; Canfield, R.L.; Dietrich, K.N.; Bornschein, R.; Greene, T.; et al. Low-Level Environmental Lead Exposure and Children’s Intellectual Function: An International Pooled Analysis. Environ. Health Perspect. 2005, 113, 894–899. [Google Scholar] [CrossRef]
- Reuben, A. Childhood Lead Exposure and Adult Neurodegenerative Disease. J. Alzheimers Dis. JAD 2018, 64, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Watt, G.C.; Britton, A.; Gilmour, H.G.; Moore, M.R.; Murray, G.D.; Robertson, S.J. Public Health Implications of New Guidelines for Lead in Drinking Water: A Case Study in an Area with Historically High Water Lead Levels. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2000, 38, 73–79. [Google Scholar] [CrossRef]
- Iternational Agency for Research on Cancer. Cadmium and Cadmium Compounds. IARC Monogr. Eval. Carcinog. Risks Hum. 1993, 58, 119–237. [Google Scholar]
- Järup, L.; Akesson, A. Current Status of Cadmium as an Environmental Health Problem. Toxicol. Appl. Pharmacol. 2009, 238, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, M.; Micali, A.; Marini, H.; Adamo, E.B.; Puzzolo, D.; Pisani, A.; Trichilo, V.; Altavilla, D.; Squadrito, F.; Minutoli, L. Cadmium, Organ Toxicity and Therapeutic Approaches: A Review on Brain, Kidney and Testis Damage. Curr. Med. Chem. 2017, 24, 3879–3893. [Google Scholar] [CrossRef]
- Satarug, S.; Garrett, S.H.; Sens, M.A.; Sens, D.A. Cadmium, Environmental Exposure, and Health Outcomes. Environ. Health Perspect. 2010, 118, 182–190. [Google Scholar] [CrossRef]
- Buha, A.; Đukić-Ćosić, D.; Ćurčić, M.; Bulat, Z.; Antonijević, B.; Moulis, J.-M.; Goumenou, M.; Wallace, D. Emerging Links between Cadmium Exposure and Insulin Resistance: Human, Animal, and Cell Study Data. Toxics 2020, 8, 63. [Google Scholar] [CrossRef]
- Skalny, A.V.; Lima, T.R.R.; Ke, T.; Zhou, J.-C.; Bornhorst, J.; Alekseenko, S.I.; Aaseth, J.; Anesti, O.; Sarigiannis, D.A.; Tsatsakis, A.; et al. Toxic Metal Exposure as a Possible Risk Factor for COVID-19 and Other Respiratory Infectious Diseases. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2020, 146, 111809. [Google Scholar] [CrossRef]
- Satarug, S.; Gobe, G.C.; Vesey, D.A.; Phelps, K.R. Cadmium and Lead Exposure, Nephrotoxicity, and Mortality. Toxics 2020, 8. [Google Scholar] [CrossRef]
- Hossein-Khannazer, N.; Azizi, G.; Eslami, S.; Alhassan Mohammed, H.; Fayyaz, F.; Hosseinzadeh, R.; Usman, A.B.; Kamali, A.N.; Mohammadi, H.; Jadidi-Niaragh, F.; et al. The Effects of Cadmium Exposure in the Induction of Inflammation. Immunopharmacol. Immunotoxicol. 2020, 42, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Breton, J.; Daniel, C.; Vignal, C.; Body-Malapel, M.; Garat, A.; Plé, C.; Foligné, B. Does Oral Exposure to Cadmium and Lead Mediate Susceptibility to Colitis? The Dark-and-Bright Sides of Heavy Metals in Gut Ecology. Sci. Rep. 2016, 6, 19200. [Google Scholar] [CrossRef] [Green Version]
- Sanders, A.P.; Claus Henn, B.; Wright, R.O. Perinatal and Childhood Exposure to Cadmium, Manganese, and Metal Mixtures and Effects on Cognition and Behavior: A Review of Recent Literature. Curr. Environ. Health Rep. 2015, 2, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Becaria, A.; Campbell, A.; Bondy, S.C. Aluminum as a Toxicant. Toxicol. Ind. Health 2002, 18, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Igbokwe, I.O.; Igwenagu, E.; Igbokwe, N.A. Aluminium Toxicosis: A Review of Toxic Actions and Effects. Interdiscip. Toxicol. 2019, 12, 45–70. [Google Scholar] [CrossRef] [Green Version]
- Exley, C.; Clarkson, E. Aluminium in Human Brain Tissue from Donors without Neurodegenerative Disease: A Comparison with Alzheimer’s Disease, Multiple Sclerosis and Autism. Sci. Rep. 2020, 10, 7770. [Google Scholar] [CrossRef]
- Jeong, C.H.; Kwon, H.C.; Kim, D.H.; Cheng, W.N.; Kang, S.; Shin, D.-M.; Yune, J.H.; Yoon, J.E.; Chang, Y.H.; Sohn, H.; et al. Effects of Aluminum on the Integrity of the Intestinal Epithelium: An in Vitro and in Vivo Study. Environ. Health Perspect. 2020, 128, 17013. [Google Scholar] [CrossRef] [Green Version]
- Esquerre, N.; Basso, L.; Dubuquoy, C.; Djouina, M.; Chappard, D.; Blanpied, C.; Desreumaux, P.; Vergnolle, N.; Vignal, C.; Body-Malapel, M. Aluminum Ingestion Promotes Colorectal Hypersensitivity in Rodents. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A. Aluminum Is a Potential Environmental Factor for Crohn’s Disease Induction: Extended Hypothesis. Ann. N. Y. Acad. Sci. 2007, 1107, 329–345. [Google Scholar] [CrossRef]
- Pineton de Chambrun, G.; Body-Malapel, M.; Frey-Wagner, I.; Djouina, M.; Deknuydt, F.; Atrott, K.; Esquerre, N.; Altare, F.; Neut, C.; Arrieta, M.C.; et al. Aluminum Enhances Inflammation and Decreases Mucosal Healing in Experimental Colitis in Mice. Mucosal Immunol. 2014, 7, 589–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vignal, C.; Desreumaux, P.; Body-Malapel, M. Gut: An Underestimated Target Organ for Aluminum. Morphol. Bull. Assoc. Anat. 2016, 100, 75–84. [Google Scholar] [CrossRef]
- Röllin, H.B.; Nogueira, C.; Olutola, B.; Channa, K.; Odland, J.Ø. Prenatal Exposure to Aluminum and Status of Selected Essential Trace Elements in Rural South African Women at Delivery. Int. J. Environ. Res. Public. Health 2018, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, H.; Yu, L.; Tian, F.; Zhai, Q.; Fan, L.; Chen, W. Gut Microbiota: A Target for Heavy Metal Toxicity and a Probiotic Protective Strategy. Sci. Total Environ. 2020, 742, 140429. [Google Scholar] [CrossRef] [PubMed]
- Assefa, S.; Köhler, G. Intestinal Microbiome and Metal Toxicity. Curr. Opin. Toxicol. 2020, 19, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Tsiaoussis, J.; Antoniou, M.N.; Koliarakis, I.; Mesnage, R.; Vardavas, C.I.; Izotov, B.N.; Psaroulaki, A.; Tsatsakis, A. Effects of Single and Combined Toxic Exposures on the Gut Microbiome: Current Knowledge and Future Directions. Toxicol. Lett. 2019, 312, 72–97. [Google Scholar] [CrossRef]
- Bolan, S.; Seshadri, B.; Grainge, I.; Talley, N.J.; Naidu, R. Gut Microbes Modulate Bioaccessibility of Lead in Soil. Chemosphere 2020, 128657. [Google Scholar] [CrossRef]
- Nakamura, I.; Hosokawa, K.; Tamura, H.; Miura, T. Reduced Mercury Excretion with Feces in Germfree Mice after Oral Administration of Methyl Mercury Chloride. Bull. Environ. Contam. Toxicol. 1977, 17, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Qu, D.; Feng, S.; Yu, Y.; Yu, L.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Oral Supplementation of Lead-Intolerant Intestinal Microbes Protects Against Lead (Pb) Toxicity in Mice. Front. Microbiol. 2019, 10, 3161. [Google Scholar] [CrossRef]
- Gupta, A.; Joia, J. Microbes as Potential Tool for Remediation of Heavy Metals: A Review. J. Microb. Biochem. Technol. 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Monachese, M.; Burton, J.P.; Reid, G. Bioremediation and Tolerance of Humans to Heavy Metals through Microbial Processes: A Potential Role for Probiotics? Appl. Environ. Microbiol. 2012, 78, 6397–6404. [Google Scholar] [CrossRef] [Green Version]
- George, F.; Daniel, C.; Thomas, M.; Singer, E.; Guilbaud, A.; Tessier, F.J.; Revol-Junelles, A.-M.; Borges, F.; Foligné, B. Occurrence and Dynamism of Lactic Acid Bacteria in Distinct Ecological Niches: A Multifaceted Functional Health Perspective. Front. Microbiol. 2018, 9, 2899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, F.; Halttunen, T.; Tahvonen, R.; Salminen, S. Probiotic Bacteria as Potential Detoxification Tools: Assessing Their Heavy Metal Binding Isotherms. Can. J. Microbiol. 2006, 52, 877–885. [Google Scholar] [CrossRef]
- Halttunen, T.; Salminen, S.; Tahvonen, R. Rapid Removal of Lead and Cadmium from Water by Specific Lactic Acid Bacteria. Int. J. Food Microbiol. 2007, 114, 30–35. [Google Scholar] [CrossRef]
- Teemu, H.; Seppo, S.; Jussi, M.; Raija, T.; Kalle, L. Reversible Surface Binding of Cadmium and Lead by Lactic Acid and Bifidobacteria. Int. J. Food Microbiol. 2008, 125, 170–175. [Google Scholar] [CrossRef]
- Bhakta, J.N.; Ohnishi, K.; Munekage, Y.; Iwasaki, K.; Wei, M.Q. Characterization of Lactic Acid Bacteria-Based Probiotics as Potential Heavy Metal Sorbents. J. Appl. Microbiol. 2012, 112, 1193–1206. [Google Scholar] [CrossRef]
- Kinoshita, H.; Sohma, Y.; Ohtake, F.; Ishida, M.; Kawai, Y.; Kitazawa, H.; Saito, T.; Kimura, K. Biosorption of Heavy Metals by Lactic Acid Bacteria and Identification of Mercury Binding Protein. Res. Microbiol. 2013, 164, 701–709. [Google Scholar] [CrossRef]
- Topcu, A.; Bulat, T. Removal of Cadmium and Lead from Aqueous Solution by Enterococcus Faecium Strains. J. Food Sci. 2010, 75, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Halttunen, T.; Collado, M.C.; El-Nezami, H.; Meriluoto, J.; Salminen, S. Combining Strains of Lactic Acid Bacteria May Reduce Their Toxin and Heavy Metal Removal Efficiency from Aqueous Solution. Lett. Appl. Microbiol. 2008, 46, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Daisley, B.A.; Monachese, M.; Trinder, M.; Bisanz, J.E.; Chmiel, J.A.; Burton, J.P.; Reid, G. Immobilization of Cadmium and Lead by Lactobacillus rhamnosus GR-1 Mitigates Apical-to-Basolateral Heavy Metal Translocation in a Caco-2 Model of the Intestinal Epithelium. Gut Microbes 2019, 10, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Zhai, Q.; Zhao, J.; Liu, X.; Wang, G.; Zhang, H.; Zhang, H.; Chen, W. Lactobacillus plantarum CCFM8661 Alleviates Lead Toxicity in Mice. Biol. Trace Elem. Res. 2012, 150, 264–271. [Google Scholar] [CrossRef]
- Li, B.; Jin, D.; Yu, S.; Etareri Evivie, S.; Muhammad, Z.; Huo, G.; Liu, F. In Vitro and In Vivo Evaluation of Lactobacillus delbrueckii Subsp. bulgaricus KLDS1.0207 for the Alleviative Effect on Lead Toxicity. Nutrients 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Q.; Wang, G.; Zhao, J.; Liu, X.; Narbad, A.; Chen, Y.Q.; Zhang, H.; Tian, F.; Chen, W. Protective Effects of Lactobacillus Plantarum CCFM8610 against Chronic Cadmium Toxicity in Mice Indicate Routes of Protection besides Intestinal Sequestration. Appl. Environ. Microbiol. 2014, 80, 4063–4071. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Q.; Wang, G.; Zhao, J.; Liu, X.; Tian, F.; Zhang, H.; Chen, W. Protective Effects of Lactobacillus plantarum CCFM8610 against Acute Cadmium Toxicity in Mice. Appl. Environ. Microbiol. 2013, 79, 1508–1515. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Zhai, Q.; Yin, R.; Li, P.; Tian, F.; Liu, X.; Zhao, J.; Gong, J.; Zhang, H.; Chen, W. Lactobacillus plantarum CCFM639 Alleviate Trace Element Imbalance-Related Oxidative Stress in Liver and Kidney of Chronic Aluminum Exposure Mice. Biol. Trace Elem. Res. 2017, 176, 342–349. [Google Scholar] [CrossRef]
- Yu, L.; Zhai, Q.; Liu, X.; Wang, G.; Zhang, Q.; Zhao, J.; Narbad, A.; Zhang, H.; Tian, F.; Chen, W. Lactobacillus plantarum CCFM639 Alleviates Aluminium Toxicity. Appl. Microbiol. Biotechnol. 2016, 100, 1891–1900. [Google Scholar] [CrossRef]
- Bron, P.A.; Tomita, S.; Mercenier, A.; Kleerebezem, M. Cell Surface-Associated Compounds of Probiotic Lactobacilli Sustain the Strain-Specificity Dogma. Curr. Opin. Microbiol. 2013, 16, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Kumari, V.; Ram, C.; Thakur, K.; Tomar, S.K. Bio-Prospectus of Cadmium Bioadsorption by Lactic Acid Bacteria to Mitigate Health and Environmental Impacts. Appl. Microbiol. Biotechnol. 2018, 102, 1599–1615. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Harris, H.M.B.; McCann, A.; Guo, C.; Argimón, S.; Zhang, W.; Yang, X.; Jeffery, I.B.; Cooney, J.C.; Kagawa, T.F.; et al. Expanding the Biotechnology Potential of Lactobacilli through Comparative Genomics of 213 Strains and Associated Genera. Nat. Commun. 2015, 6, 8322. [Google Scholar] [CrossRef]
- Deutsch, S.-M.; Mariadassou, M.; Nicolas, P.; Parayre, S.; Le Guellec, R.; Chuat, V.; Peton, V.; Le Maréchal, C.; Burati, J.; Loux, V.; et al. Identification of Proteins Involved in the Anti-Inflammatory Properties of Propionibacterium freudenreichii by Means of a Multi-Strain Study. Sci. Rep. 2017, 7, 46409. [Google Scholar] [CrossRef]
- Ochman, H.; Selander, R.K. Standard Reference Strains of Escherichia coli from Natural Populations. J. Bacteriol. 1984, 157, 690–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmouni, O.; Vignal, C.; Titécat, M.; Foligné, B.; Pariente, B.; Dubuquoy, L.; Desreumaux, P.; Neut, C. High Carriage of Adherent Invasive E. coli in Wildlife and Healthy Individuals. Gut Pathog. 2018, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Pradel, E.; Zhang, Y.; Pujol, N.; Matsuyama, T.; Bargmann, C.I.; Ewbank, J.J. Detection and Avoidance of a Natural Product from the Pathogenic Bacterium Serratia marcescens by Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2007, 104, 2295–2300. [Google Scholar] [CrossRef] [Green Version]
- Adouard, N.; Foligné, B.; Dewulf, J.; Bouix, M.; Picque, D.; Bonnarme, P. In Vitro Characterization of the Digestive Stress Response and Immunomodulatory Properties of Microorganisms Isolated from Smear-Ripened Cheese. Int. J. Food Microbiol. 2015, 197, 98–107. [Google Scholar] [CrossRef]
- Malik, A.C.; Reinbold, G.W.; Vedamuthu, E.R. An Evaluation of the Taxonomy of Propionibacterium. Can. J. Microbiol. 1968, 14, 1185–1191. [Google Scholar] [CrossRef]
- Ameen, F.A.; Hamdan, A.M.; El-Naggar, M.Y. Assessment of the Heavy Metal Bioremediation Efficiency of the Novel Marine Lactic Acid Bacterium, Lactobacillus plantarum MF042018. Sci. Rep. 2020, 10, 314. [Google Scholar] [CrossRef]
- Lin, D.; Ji, R.; Wang, D.; Xiao, M.; Zhao, J.; Zou, J.; Li, Y.; Qin, T.; Xing, B.; Chen, Y.; et al. The Research Progress in Mechanism and Influence of Biosorption between Lactic Acid Bacteria and Pb(II): A Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 395–410. [Google Scholar] [CrossRef]
- Muhammad, Z.; Ramzan, R.; Zhang, S.; Hu, H.; Hameed, A.; Bakry, A.M.; Dong, Y.; Wang, L.; Pan, S. Comparative Assessment of the Bioremedial Potentials of Potato Resistant Starch-Based Microencapsulated and Non-Encapsulated Lactobacillus plantarum to Alleviate the Effects of Chronic Lead Toxicity. Front. Microbiol. 2018, 9, 1306. [Google Scholar] [CrossRef] [Green Version]
- Mohapatra, R.K.; Parhi, P.K.; Pandey, S.; Bindhani, B.K.; Thatoi, H.; Panda, C.R. Active and Passive Biosorption of Pb(II)Using Live and Dead Biomass of Marine Bacterium Bacillus xiamenensis PbRPSD202: Kinetics and Isotherm Studies. J. Environ. Manag. 2019, 247, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Kirillova, A.V.; Danilushkina, A.A.; Irisov, D.S.; Bruslik, N.L.; Fakhrullin, R.F.; Zakharov, Y.A.; Bukhmin, V.S.; Yarullina, D.R. Assessment of Resistance and Bioremediation Ability of Lactobacillus Strains to Lead and Cadmium. Int. J. Microbiol. 2017, 2017, 9869145. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.; Cao, H.; Zhong, Y.; Huang, Y.; Zou, J.; He, Q.; Ji, R.; Qin, T.; Chen, Y.; Wang, D.; et al. Screening and Identification of Lactic Acid Bacteria from Ya’an Pickle Water to Effectively Remove Pb2. AMB Express 2019, 9, 10. [Google Scholar] [CrossRef]
- Foligné, B.; Breton, J.; Mater, D.; Jan, G. Tracking the Microbiome Functionality: Focus on Propionibacterium Species. Gut 2013, 62, 1227–1228. [Google Scholar] [CrossRef]
- Djurasevic, S.; Jama, A.; Jasnic, N.; Vujovic, P.; Jovanovic, M.; Mitic-Culafic, D.; Knezevic-Vukcevic, J.; Cakic-Milosevic, M.; Ilijevic, K.; Djordjevic, J. The Protective Effects of Probiotic Bacteria on Cadmium Toxicity in Rats. J. Med. Food 2017, 20, 189–196. [Google Scholar] [CrossRef]
- Bisanz, J.E.; Enos, M.K.; Mwanga, J.R.; Changalucha, J.; Burton, J.P.; Gloor, G.B.; Reid, G. Randomized Open-Label Pilot Study of the Influence of Probiotics and the Gut Microbiome on Toxic Metal Levels in Tanzanian Pregnant Women and School Children. mBio 2014, 5, e01580-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Q.; Wang, J.; Cen, S.; Zhao, J.; Zhang, H.; Tian, F.; Chen, W. Modulation of the Gut Microbiota by a Galactooligosaccharide Protects against Heavy Metal Lead Accumulation in Mice. Food Funct. 2019, 10, 3768–3781. [Google Scholar] [CrossRef]
- Bhattacharya, S. The Role of Probiotics in the Amelioration of Cadmium Toxicity. Biol. Trace Elem. Res. 2020, 197, 440–444. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, F.; Mahieux, S.; Daniel, C.; Titécat, M.; Beauval, N.; Houcke, I.; Neut, C.; Allorge, D.; Borges, F.; Jan, G.; et al. Assessment of Pb(II), Cd(II), and Al(III) Removal Capacity of Bacteria from Food and Gut Ecological Niches: Insights into Biodiversity to Limit Intestinal Biodisponibility of Toxic Metals. Microorganisms 2021, 9, 456. https://doi.org/10.3390/microorganisms9020456
George F, Mahieux S, Daniel C, Titécat M, Beauval N, Houcke I, Neut C, Allorge D, Borges F, Jan G, et al. Assessment of Pb(II), Cd(II), and Al(III) Removal Capacity of Bacteria from Food and Gut Ecological Niches: Insights into Biodiversity to Limit Intestinal Biodisponibility of Toxic Metals. Microorganisms. 2021; 9(2):456. https://doi.org/10.3390/microorganisms9020456
Chicago/Turabian StyleGeorge, Fanny, Séverine Mahieux, Catherine Daniel, Marie Titécat, Nicolas Beauval, Isabelle Houcke, Christel Neut, Delphine Allorge, Frédéric Borges, Gwénaël Jan, and et al. 2021. "Assessment of Pb(II), Cd(II), and Al(III) Removal Capacity of Bacteria from Food and Gut Ecological Niches: Insights into Biodiversity to Limit Intestinal Biodisponibility of Toxic Metals" Microorganisms 9, no. 2: 456. https://doi.org/10.3390/microorganisms9020456
APA StyleGeorge, F., Mahieux, S., Daniel, C., Titécat, M., Beauval, N., Houcke, I., Neut, C., Allorge, D., Borges, F., Jan, G., Foligné, B., & Garat, A. (2021). Assessment of Pb(II), Cd(II), and Al(III) Removal Capacity of Bacteria from Food and Gut Ecological Niches: Insights into Biodiversity to Limit Intestinal Biodisponibility of Toxic Metals. Microorganisms, 9(2), 456. https://doi.org/10.3390/microorganisms9020456