
Shift from Carbon Flow through the Microbial Loop to the Viral Shunt in Coastal Antarctic Waters during Austral Summer

 , ,
, , _Brussaard.png)
Abstract
:1. Introduction
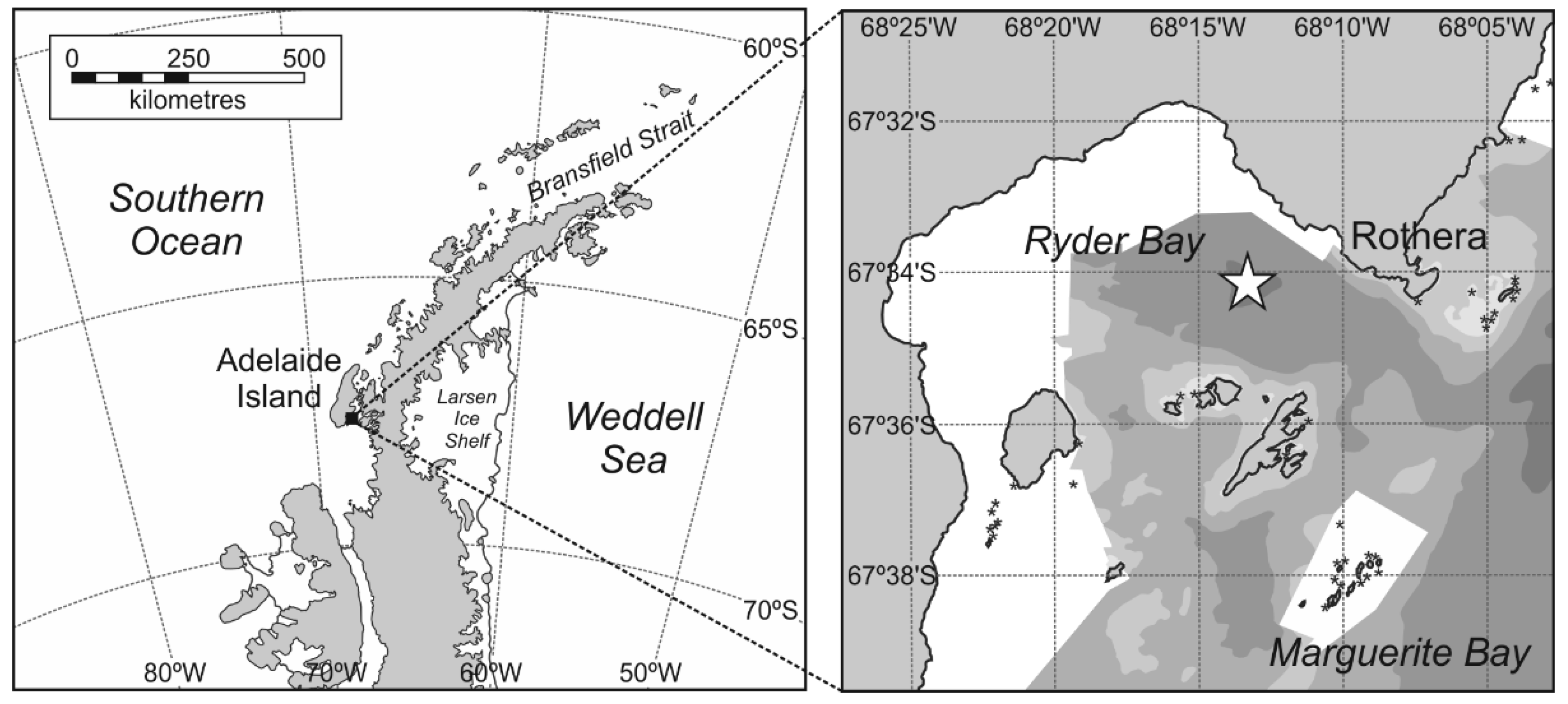
2. Materials and Methods
3. Results
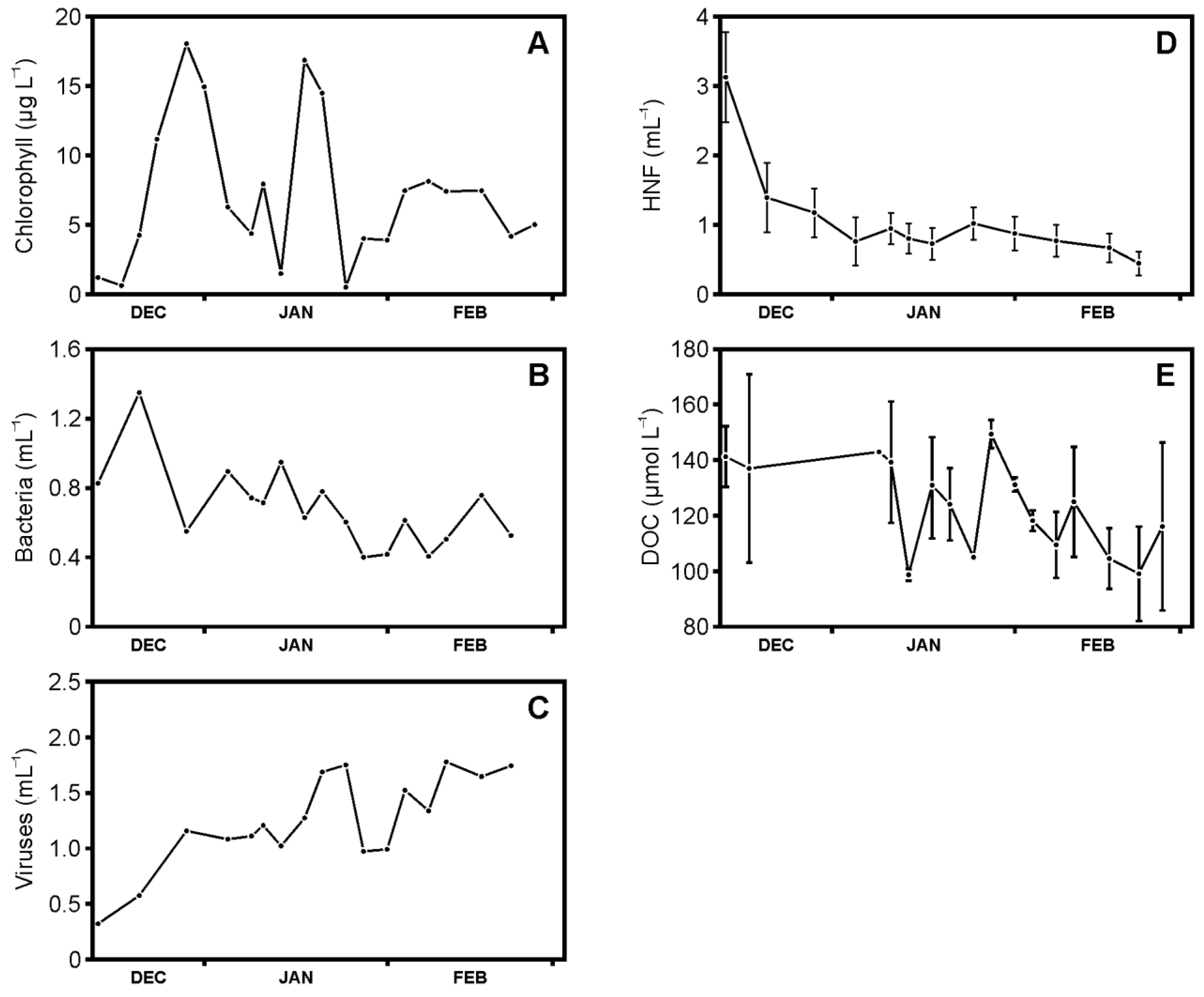
3.1. Abundances of Phytoplankton, Bacteria, Viruses, and Heterotrophic Nanoflagellates, and Concentrations of Dissolved Organic Carbon
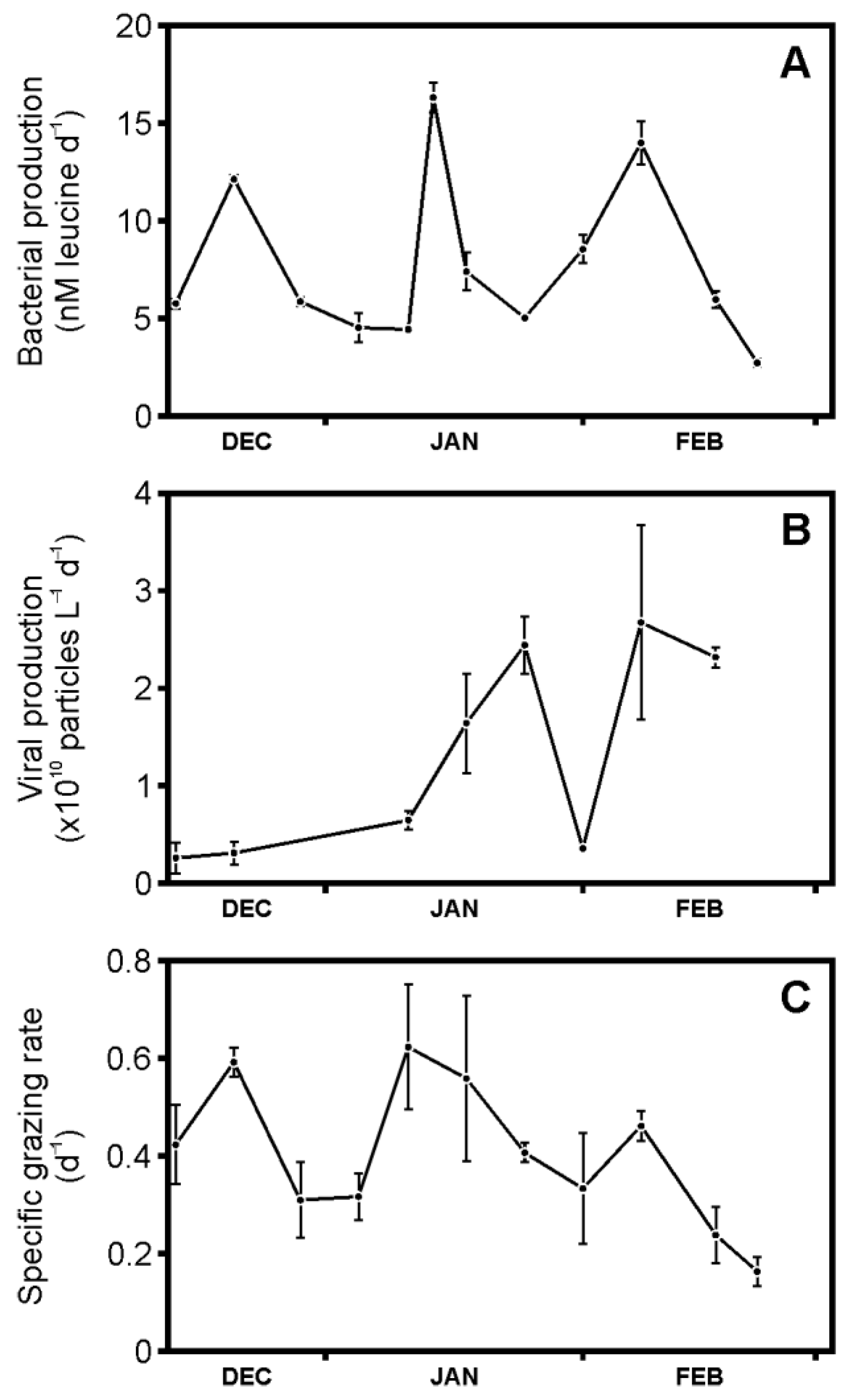
3.2. Bacterial Production, Viral Production, and Grazing Rates
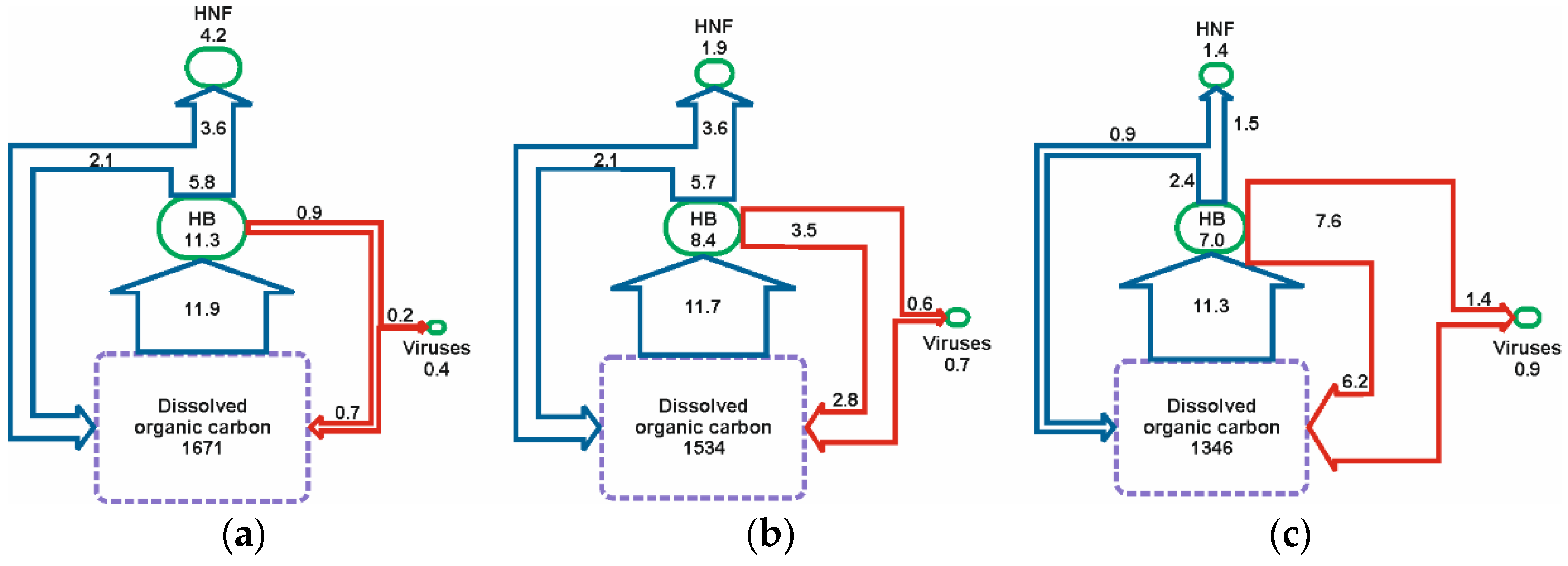
3.3. Budget of Carbon Flow
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pomeroy, L.R. The Ocean’s Food Web, A Changing Paradigm. BioScience 1974, 24, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Azam, T.; Field, J.G.; Gray, J.S.; Reil, M.L.A.; Thingstad, F. The Ecological Role of Water-Column Microbes in the Sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Sherr, B.F.; Sherr, E.B. Proportional distribution of total numbers, biovolume, and bacterivory among size classes of 2–20 mm nonpigmented marine flagellates. Mar. Microb. Food Webs 1991, 5, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Azam, F.; Smith, D.C.; Hollibaugh, J.T. The role of the microbial loop in Antarctic pelagic ecosystems. In Proceedings of the Pro Mare Symposium on Polar Marine Ecology, Trondheim, Norway, 12–16 May 1990; Sakshaug, E., Hopkins, C.C.E., Oritsland, N.A., Eds.; Polar Research: Tromsø, Norway, 1990; Volume 10, pp. 239–243. [Google Scholar]
- Fuhrman, J.A. Marine Viruses and their biogeochemical and ecological effects. Nature 1999, 399, 541–548. [Google Scholar] [CrossRef]
- Middelboe, M.; Jørgensen, N.O.G.; Kroer, N. Effects of viruses on nutrient turnover and growth efficiency of noninfected marine bacterioplankton. Appl. Environ. Microbiol. 1996, 62, 1991–1997. [Google Scholar] [CrossRef] [Green Version]
- Suttle, C.A. Marine viruses-major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ducklow, H. A decadal (2002–2014) analysis for dynamics of heterotrophic bacteria in an Antarctic coastal ecosystem: Variability and physical and biogeochemical forcings. Front. Mar. Sci. 2016, 3, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Carlson, C.A.; Hansell, D.A. DOM Sources, Sinks, Reactivity, and Budgets. In Biogeochemistry of Marine Dissolved Organic Matter, 2nd ed.; Hansell, D.A., Carlson, C.A., Eds.; Elsevier: London, UK, 2014; pp. 65–126. [Google Scholar]
- Vaqué, D.; Agustí, S.; Duarte, C.M. Response of bacterial grazing rates to experimental manipulation of an Antarctic coastal nanoflagellate community. Aquat. Microb. Ecol. 2004, 41, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Tsai, A.Y.; Gong, G.C.; Huang, Y.W. Variations of microbial loop carbon flux in western subtropical Pacific coastal water between warm and cold season. J. Exp. Mar. Biol. Ecol. 2013, 449, 111–117. [Google Scholar] [CrossRef]
- Thingstad, T.F.; Lignell, R. Theoretical models for the control of bacterial growth rate, abundance, diversity and carbon demand. Aquat. Microb. Ecol. 1997, 13, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Thingstad, T.F.; Våge, S.; Storesund, J.E.; Sandaa, R.A.; Giske, J. A theoretical analysis of how strain-specific viruses can control microbial species diversity. Proc. Natl. Acad. Sci. USA 2014, 111, 7813–7818. [Google Scholar] [CrossRef] [Green Version]
- Jürgens, K.; Güde, H. The potential importance of grazing-resistant bacteria in planktonic systems. Mar. Ecol. Prog. Ser. 1994, 112, 169–188. [Google Scholar] [CrossRef]
- Clarke, A.; Meredith, M.P.; Wallace, M.I.; Brandon, M.A.; Thomas, D.N. Seasonal and interannual variability in temperature, chlorophyll and macronutrients in northern Marguerite Bay, Antarctica. Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 18–19. [Google Scholar] [CrossRef]
- Ducklow, H.W.; Schofield, O.; Vernet, M.; Stammerjohn, S.; Erickson, M. Multiscale control of bacterial production by phytoplankton dynamics and sea ice along the western Antarctic Peninsula: A regional and decadal investigation. J. Mar. Syst. 2012, 98–99, 26–39. [Google Scholar] [CrossRef]
- Venables, H.J.; Clarke, A.; Meredith, M.P. Wintertime controls on summer stratification and productivity at the western Antarctic Peninsula. Limnol. Oceanogr. 2013, 58, 1035–1047. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.; Brandsma, J.; Pond, D.W.; Venables, H.J.; Meredith, M.P.; Witte, H.J.; Stammerjohn, S.; Wilson, W.H.; Clarke, A.; Brussaard, C.P.D. Drivers of interannual variability in virioplankton abundance at the coastal Western Antarctic Peninsula and the potential effects of climate change. Environ. Microbiol. 2016, 19, 740–755. [Google Scholar] [CrossRef]
- Mojica, K.D.A.; Evans, C.; Brussaard, C.P.D. Flow cytometric enumeration of marine viral populations at low abundances. Aquat. Microb. Ecol. 2014, 71, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Marie, D.; Partensky, F.; Vaulot, D.; Brussaard, C.P.D. Enumeration of phytoplankton, bacteria, and viruses in marine samples. In Current Protocols in Cytometry; Robinson, J.P., Ed.; John Wiley & Sons: New York, NY, USA, 1999; pp. 11.11.1–11.11.15. [Google Scholar]
- Qian, J.; Mopper, K. Automated high performance, high-temperature combustion total carbon analyzer. Anal. Chem. 1996, 68, 3090–3097. [Google Scholar] [CrossRef]
- Simon, M.; Azam, F. Protein content and protein synthesis rates of planktonic marine bacteria. Mar. Ecol. Prog. Ser. 1989, 51, 201–213. [Google Scholar] [CrossRef]
- Ducklow, H.W.; Dickson, M.L.; Kirchman, D.L.; Steward, G.; Orchardo, J.; Marra, J.; Azam, F. Constraining bacterial production, conversion efficiency and respiration in the Ross Sea, Antarctica, January–February, 1997. Deep Sea Res. Part II Top. Stud. Oceanogr. 2000, 47, 3227–3247. [Google Scholar] [CrossRef]
- Winget, D.M.; Williamson, K.E.; Helton, R.R.; Wommack, K.E. Tangential flow diafiltration: An improved technique for estimation of virioplankton production. Aquat. Microb. Ecol. 2005, 41, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, S.W.; Brigden, S.N.; Suttle, C.A. A dilution technique for the direct measurement of viral production: A comparison in stratified and tidally mixed coastal waters. Microb. Ecol. 2002, 43, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Sherr, B.F.; Sherr, E.B.; Fallon, R.D. Use of monodispersed, fluorescently labeled bacteria to estimate in situ protozoan bacterivory. Appl. Environ. Microbiol. 1987, 53, 958–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salat, J.; Marrasé, C. Exponential and linear estimations of grazing on bacteria: Effects of changes in the proportion of marked cells. Mar. Ecol. Prog. Ser. 1994, 104, 205–209. [Google Scholar] [CrossRef]
- Fukuda, R.; Ogawa, H.; Nagata, T.; Koike, I. Direct determination of carbon and nitrogen contents of natural bacterial assemblages in marine environments. Appl. Environ. Microbiol. 1998, 64, 3352–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jover, L.F.; Effler, T.C.; Buchan, A.; Wilhelm, S.W.; Weitz, J.S. The elemental composition of virus particles: Implications for marine biogeochemical cycles. Nat. Rev. Microbiol. 2014, 12, 519–528. [Google Scholar] [CrossRef]
- Børsheim, K.Y.; Bratbak, G. Cell volume to cell carbon conversion factors for bacterivorous Monas sp. Enriched from seawater. Mar. Ecol. Prog. Ser. 1987, 36, 171–175. [Google Scholar] [CrossRef]
- Baltar, F.; Arístegui, J.; Montero, M.F.; Espino, M.; Gasol, J.M.; Herndl, G.J. Mesoscale variability modulates seasonal changes in trophic structure of nano- and picoplankton communities across the NW Africa-Canary Islands transition zone. Prog. Oceanogr. 2009, 83, 180–188. [Google Scholar] [CrossRef]
- Brum, J.R.; Hurwitz, B.L.; Schofield, O.; Ducklow, H.W.; Sullivan, M.B. Seasonal time bombs: Dominant temperate viruses affect Southern Ocean microbial dynamics. ISME J. 2016, 10, 437–449. [Google Scholar] [CrossRef]
- Hagström, Å.; Azam, F.; Andersson, A.; Wikner, J.; Rassoulzadegam, F. Microbial loop in an oligotrophic pelagic marine ecosystem: Possible roles of cyanobacteria and nanoflagellates in the organic fluxes. Mar. Ecol. Prog. Ser. 1988, 49, 171–178. [Google Scholar] [CrossRef]
- Vernet, M.; Martinson, D.; Iannuzzi, R.; Stammerjohn, S.; Kozlowski, W.; Sines, K.; Smith, R.; Garibotti, I. Primary production within the sea-ice zone west of the Antarctic Peninsula: I—Sea ice, summer mixed layer, and irradiance. Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 2068–2085. [Google Scholar] [CrossRef]
- Saba, G.K.; Fraser, W.R.; Saba, V.S.; Iannuzzi, R.A.; Coleman, K.E.; Doney, S.C.; Ducklow, H.W.; Martinson, D.G.; Miles, T.N.; Patterson-Fraser, D.L.; et al. Winter and spring controls on the summer food web of the coastal West Antarctic Peninsula. Nat. Commun. 2013, 5, 4318. [Google Scholar] [CrossRef]
- Ducklow, H.W.; Carlson, C.A.; Church, M.J.; Kirchman, D.L.; Smith, D.C.; Steward, G. The seasonal development of the bacterioplankton bloom in the Ross Sea Antarctica 1994-1997. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 48, 4199–4221. [Google Scholar] [CrossRef]
- Anderson, M.R.; Rivkin, R.B. Seasonal patterns in grazing mortality of bacterioplankton in polar oceans: A bipolar comparison. Aquat. Microb. Ecol. 2001, 25, 195–206. [Google Scholar] [CrossRef]
- Tsai, A.Y.; Gong, G.C.; Hung, J. Seasonal variations of virus and nanoflagellate-mediated mortality of heterotrophic bacteria in the coastal ecosystem of subtropical western Pacific. Biogeosciences 2013, 10, 3055–3065. [Google Scholar] [CrossRef] [Green Version]
- Hyun, J.H.; Kim, S.H.; Yang, E.J.; Choi, A.; Lee, S.H. Biomass, production, and control of heterotrophic bacterioplankton during a late phytoplankton bloom in the Amundsen Sea Polynya, Antarctica. Deep Sea Res. Part II Top. Stud. Oceanogr. 2016, 123, 102–112. [Google Scholar] [CrossRef]
- Garza, D.R.; Suttle, C.A. Large double-stranded DNA viruses which cause the lysis of a marine heterotrophic nanoflagellate (Bodo sp.) occur in natural marine viral communities. Aquat. Microb. Ecol. 1995, 9, 203–210. [Google Scholar] [CrossRef]
- Massana, R.; Del Campo, J.; Dinter, C.; Sommaruga, R. Crash of a population of the marine heterotrophic flagellate Cafeteria roenbergensis by viral infection. Environ. Microbiol. 2007, 9, 2660–2669. [Google Scholar] [CrossRef] [Green Version]
- Vaqué, D.; Paz, C.J.I.; Boixereu, G.N.; Aliό, P.C. Spatial distribution of microbial biomass and activity (bacterivory and bacterial production) in the northern Weddell Sea during the austral summer (January 1994). Aquat. Microb. Ecol. 2002, 29, 107–121. [Google Scholar] [CrossRef] [Green Version]
- Boixereu, G.N.; Vaqué, D.; Gasol, J.M.; Cámara, S.J.; Alió, P.C. Viral distribution and activity in Antarctic waters. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 827–845. [Google Scholar] [CrossRef]
- Evans, C.; Brussaard, C.P.D. Regional variation in lytic and lysogenic viral infection in the Southern Ocean and its contribution to biogeochemical cycling. Appl. Environ. Microbiol. 2012, 78, 6741–6748. [Google Scholar] [CrossRef] [Green Version]
- Ghiglione, J.F.; Murray, A.E. Pronounced summer to winter differences and higher wintertime richness in coastal Antarctic marine bacterioplankton. Environ. Microbiol. 2012, 14, 617–629. [Google Scholar] [CrossRef]
- Grzymski, J.J.; Riesenfeld, C.S.; Williams, T.J.; Dussaq, A.M.; Ducklow, H.; Erickson, M.; Cavicchioli, R.; Murray, A.E. A metagenomic assessment of winter and summer bacterioplankton from Antarctica Peninsula coastal surface waters. ISME J. 2012, 6, 1901–1915. [Google Scholar] [CrossRef] [PubMed]
- Boras, J.A.; Sala, M.M.; Domínguez, V.E.; Weinbauer, M.G.; Vaqué, D. Annual changes of bacterial mortality due to viruses and protists in an oligotrophic coastal environment (NW Mediterranean). Environ. Microbiol. 2009, 11, 1181–1193. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.; Archer, S.D.; Jacquet, S.; Wilson, W.H. Direct estimates of the contribution of viral lysis and microzooplankton grazing to the decline of a Micromonas spp. population. Aquat. Microb. Ecol. 2003, 30, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Pasulka, A.L.; Samo, T.J.; Landry, M.R. Grazer and viral impacts on microbial growth and mortality in the southern California Current Ecosystem. J. Plankton Res. 2015, 37, 320–336. [Google Scholar] [CrossRef]
- Boras, J.A.; Sala, M.M.; Arrieta, J.A.; Sà, E.L.; Felipe, J.; Agustí, S.; Duarte, C.M.; Vaqué, D. Effect of ice melting on bacterial carbon fluxes channeled by viruses and protists in the Arctic Ocean. Polar Biol. 2010, 33, 1695–1707. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Hornák, K.; Jezbera, J.; Nedoma, J.; Dolan, J.R.; Šimek, K. Synergistic and antagonistic effects of viral lysis and protistan grazing on bacterial biomass, production and diversity. Environ. Microbiol. 2007, 9, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Thingstad, T.F.; Skjodal, E.F.; Bohne, R.A. Phosphorus cycling and algal-bacterial competition in Sandsfjord, western Norway. Mar. Ecol. Prog. Ser. 1993, 99, 239–259. [Google Scholar] [CrossRef]
- Monger, B.; Landry, M. Prey-size dependency of grazing by free-living marine flagellates. Mar. Ecol. Prog. Ser. 1991, 74, 239–248. [Google Scholar] [CrossRef]
- Šimek, K.; Chrzanowski, T.H. Direct and indirect evidence of size-selective grazing on pelagic bacteria by freshwater nanoflagellates. Appl. Environ. Microbiol. 1992, 58, 3715–3720. [Google Scholar] [CrossRef] [Green Version]
- Pernthaler, J.; Posch, T.; Šimek, K.; Vrba, J.; Amann, R.; Psenner, R. Contrasting bacterial strategies to coexist with a flagellate predator in an experimental microbial assemblage. Appl. Environ. Microbiol. 1997, 63, 596–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimek, K.; Vrba, J.; Pernthaler, J.; Posch, T.; Hartman, P.; Nemoda, J.; Psenner, R. Morphological and compositional shifts in an experimental bacterial community influenced by protest with contrasting feeding modes. Appl. Environ. Microbiol. 1997, 63, 587–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eissler, Y.; Sahlsten, E.; Quiñones, R.A. Effects of virus infection on respiration rates of marine phytoplankton and microplankton communities. Mar. Ecol. Prog. Ser. 2003, 262, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.; Lu, H.; White, I.; King, J.C.; Phillips, T.; Hosking, J.; Bracegirdle, T.J.; Marshall, G.J.; Mulvaney, R.; Deb, P. Absence of 21st century warming on Antarctic Peninsula consistent with natural variability. Nature 2016, 535, 411–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moline, M.A.; Claustre, H.; Frazer, T.K.; Schofield, O.; Vernet, M. Alteration of the food web along the Antarctic Peninsula in response to a regional warming trend. Glob. Chang. Biol. 2004, 10, 1973–1980. [Google Scholar] [CrossRef]
- Cota, G.F.; Kottmeier, S.T.; Robinson, D.H.; Smith, W.O., Jr.; Sullivan, C.W. Bacterioplankton in the marginal ice zone of the Weddell Sea: Biomass, production and metabolic activities during austral autumn. Deep Sea Res. 1990, 37, 1145–1167. [Google Scholar] [CrossRef]
- Manganelli, M.; Malfatti, F.; Samo, T.J.; Mitchell, B.G.; Wang, H.; Azam, F. Major role of microbes in carbon fluxes during austral winter in the Southern Drake Passage. PLoS ONE 2009, 4, e6941. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Bacterial Production (×108 Cells l−1 Day−1) | VMM (×108 Cells l−1 Day−1) a,b | GMM (×108 Cells l−1 Day−1) a,b | VMM (% of Bacterial Standing Stock d−1) a,b | GMM (% of Bacterial Standing Stock d−1) a,b | VMM (% of Total Available Bacteria d−1) a,b | GMM (% of Total Available Bacteria d−1) a,b | Bacterial Production Virally Lysed or Grazed (% d−1) a,b | VMM (µg C l−1 d−1) c | GMM (µg C l−1 d−1) c |
|---|---|---|---|---|---|---|---|---|---|---|
| 13 December 2010 | 7.0 | 0.6 | 3.9 | 7.7 | 46.9 | 4.2 | 25.4 | 64.6 | 0.8 | 4.8 |
| 20 December 2010 | 14.7 | 0.8 | 8.3 | 5.7 | 61.8 | 2.7 | 29.6 | 62.0 | 0.9 | 10.3 |
| 28 December 2010 | 7.1 | 1.7 | 31.5 | 13.7 | 2.1 | |||||
| 04 January 2011 | 5.5 | 3.3 | 38.7 | 23.7 | 4.1 | |||||
| 10 January 2011 | 5.4 | 1.6 | 5.8 | 20.1 | 74.2 | 11.9 | 44.1 | 138.1 | 2.0 | 7.2 |
| 14 January 2011 | 19.7 | |||||||||
| 24 January 2011 | 6.1 | 6.0 | 2.6 | 98.9 | 43.1 | 49.2 | 21.4 | 140.4 | 7.4 | 3.2 |
| 31 January 2011 | 10.4 | 0.9 | 1.4 | 21.0 | 34.3 | 6.0 | 9.8 | 22.2 | 1.1 | 1.8 |
| 07 February 2011 | 16.9 | 6.5 | 2.2 | 161.4 | 53.5 | 31.1 | 10.3 | 51.3 | 8.1 | 2.7 |
| 16 February 2011 | 7.2 | 5.7 | 1.9 | 74.7 | 25.5 | 38.2 | 13.1 | 105.1 | 7.0 | 2.4 |
| 21 February 2011 | 3.3 | 1.1 | 20.0 | 12.3 | 1.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, C.; Brandsma, J.; Meredith, M.P.; Thomas, D.N.; Venables, H.J.; Pond, D.W.; Brussaard, C.P.D. Shift from Carbon Flow through the Microbial Loop to the Viral Shunt in Coastal Antarctic Waters during Austral Summer. Microorganisms 2021, 9, 460. https://doi.org/10.3390/microorganisms9020460
Evans C, Brandsma J, Meredith MP, Thomas DN, Venables HJ, Pond DW, Brussaard CPD. Shift from Carbon Flow through the Microbial Loop to the Viral Shunt in Coastal Antarctic Waters during Austral Summer. Microorganisms. 2021; 9(2):460. https://doi.org/10.3390/microorganisms9020460
Chicago/Turabian StyleEvans, Claire, Joost Brandsma, Michael P. Meredith, David N. Thomas, Hugh J. Venables, David W. Pond, and Corina P. D. Brussaard. 2021. "Shift from Carbon Flow through the Microbial Loop to the Viral Shunt in Coastal Antarctic Waters during Austral Summer" Microorganisms 9, no. 2: 460. https://doi.org/10.3390/microorganisms9020460
APA StyleEvans, C., Brandsma, J., Meredith, M. P., Thomas, D. N., Venables, H. J., Pond, D. W., & Brussaard, C. P. D. (2021). Shift from Carbon Flow through the Microbial Loop to the Viral Shunt in Coastal Antarctic Waters during Austral Summer. Microorganisms, 9(2), 460. https://doi.org/10.3390/microorganisms9020460