
Direct and Indirect Effects of Management Intensity and Environmental Factors on the Functional Diversity of Lichens in Central European Forests

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
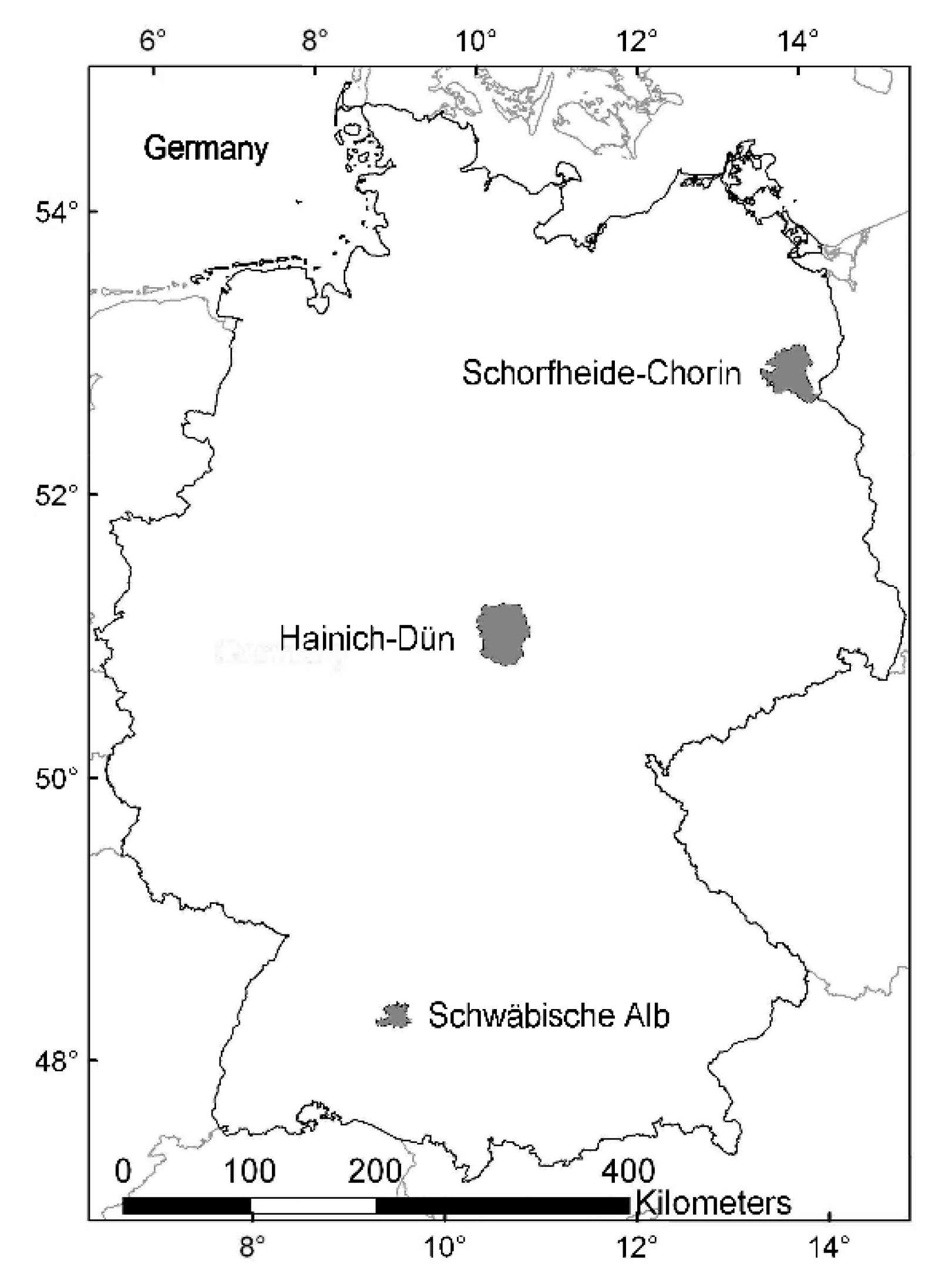
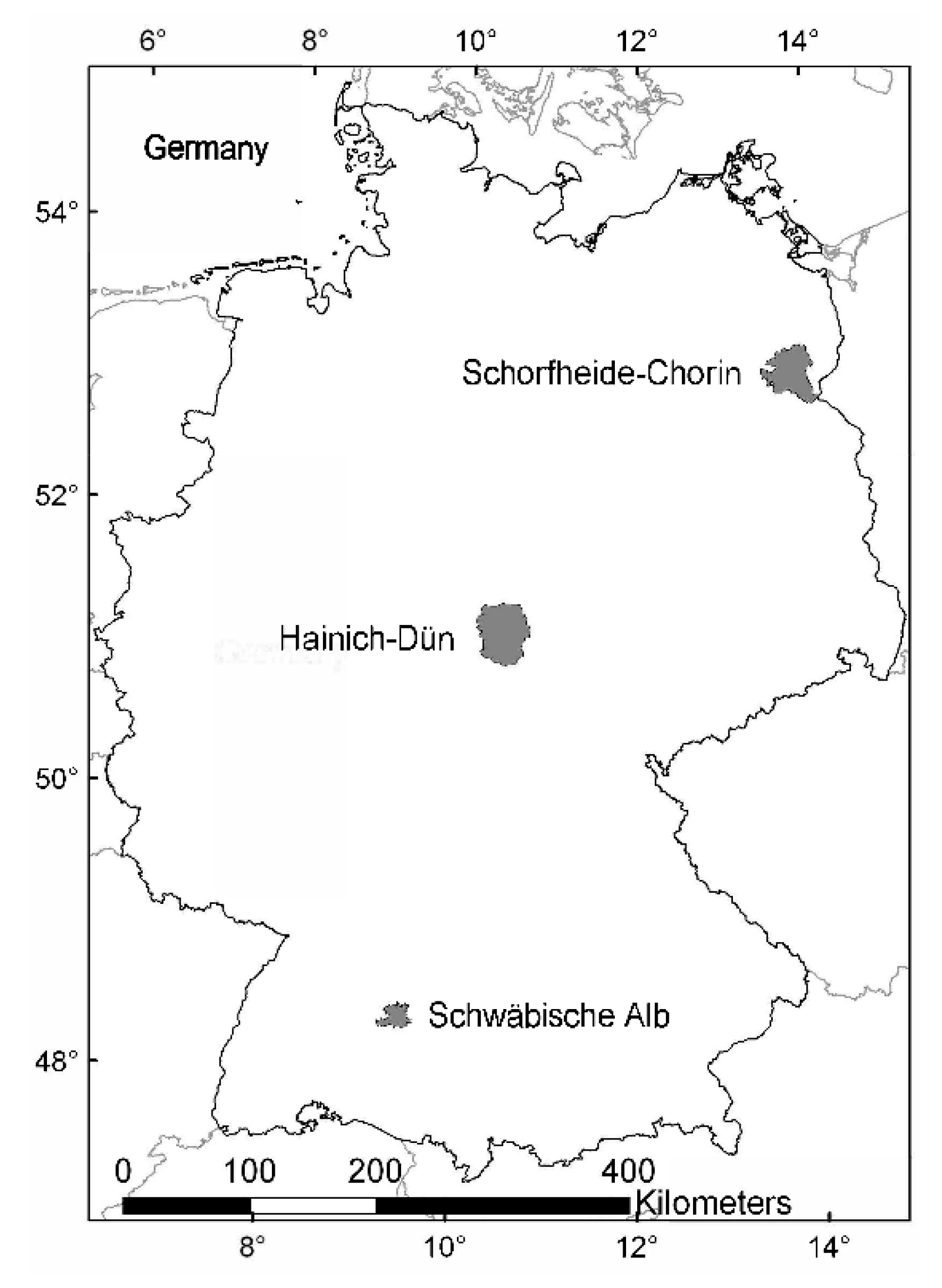
2.1. Study System
2.2. Plot Selection
2.3. Forest Management Intensity
2.4. Vegetation Data and Environmental Forest Variables
2.5. Functional Trait Data and Functional Diversity Calculations
2.6. Data Analysis
3. Results
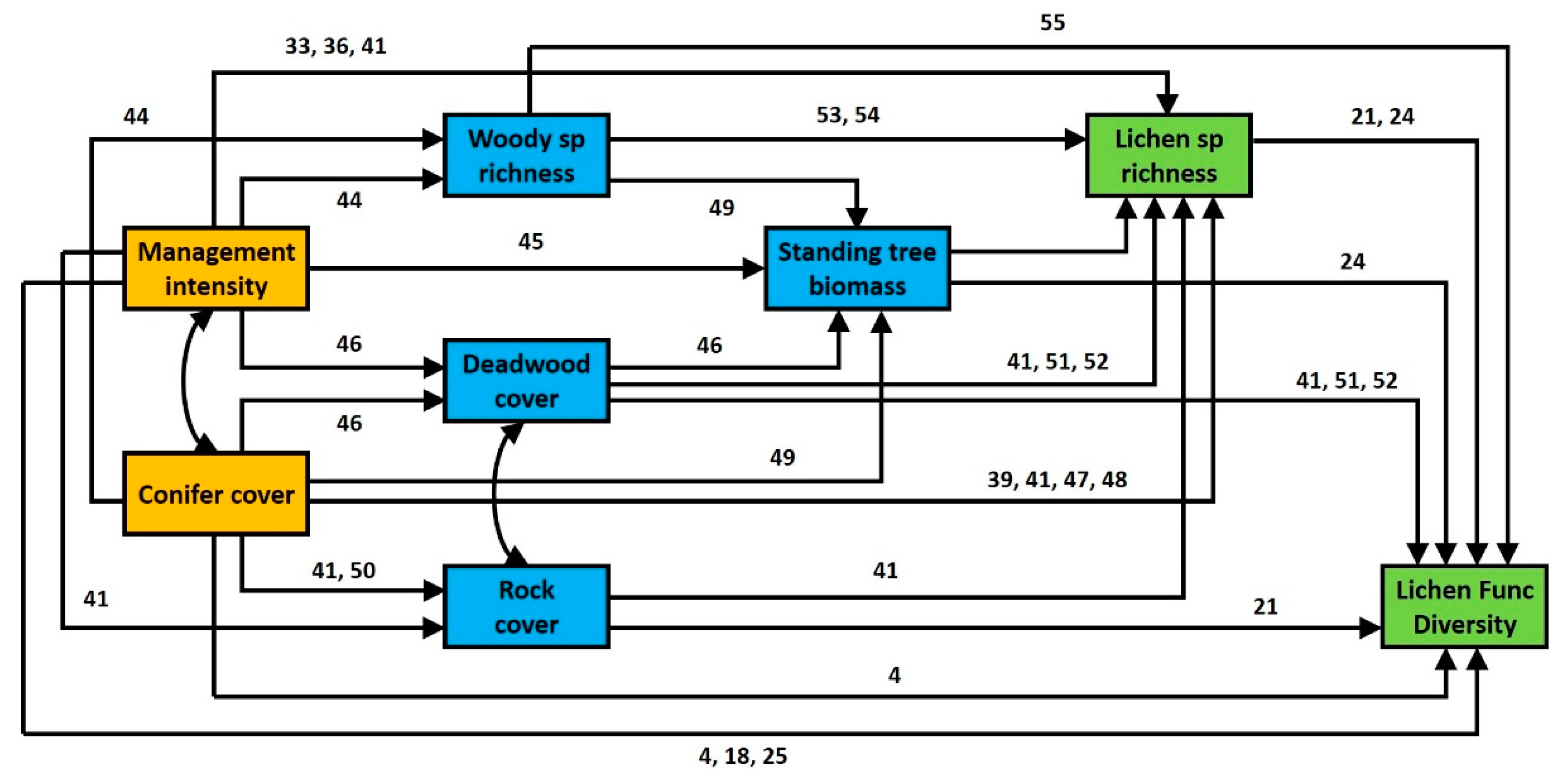
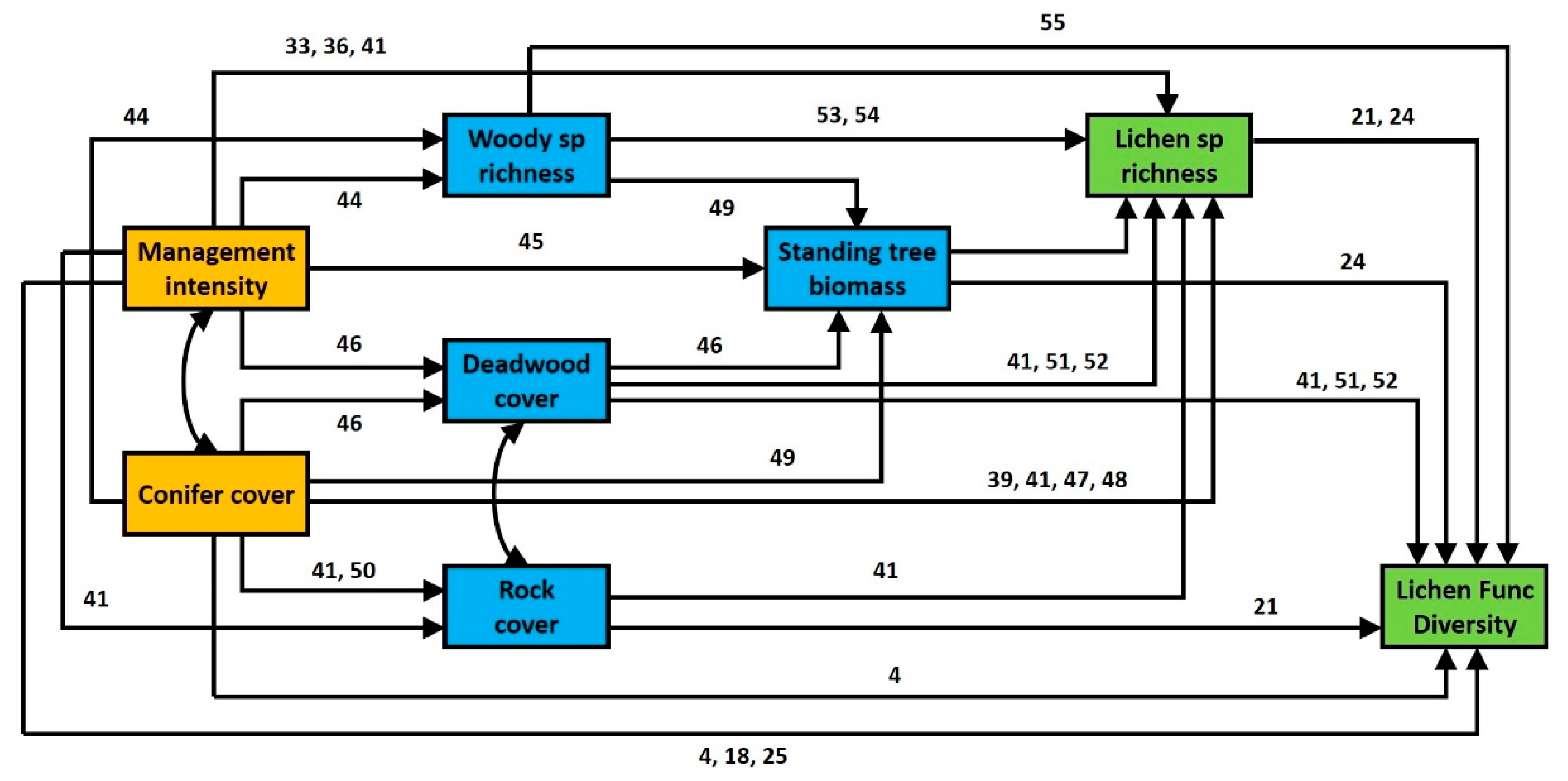
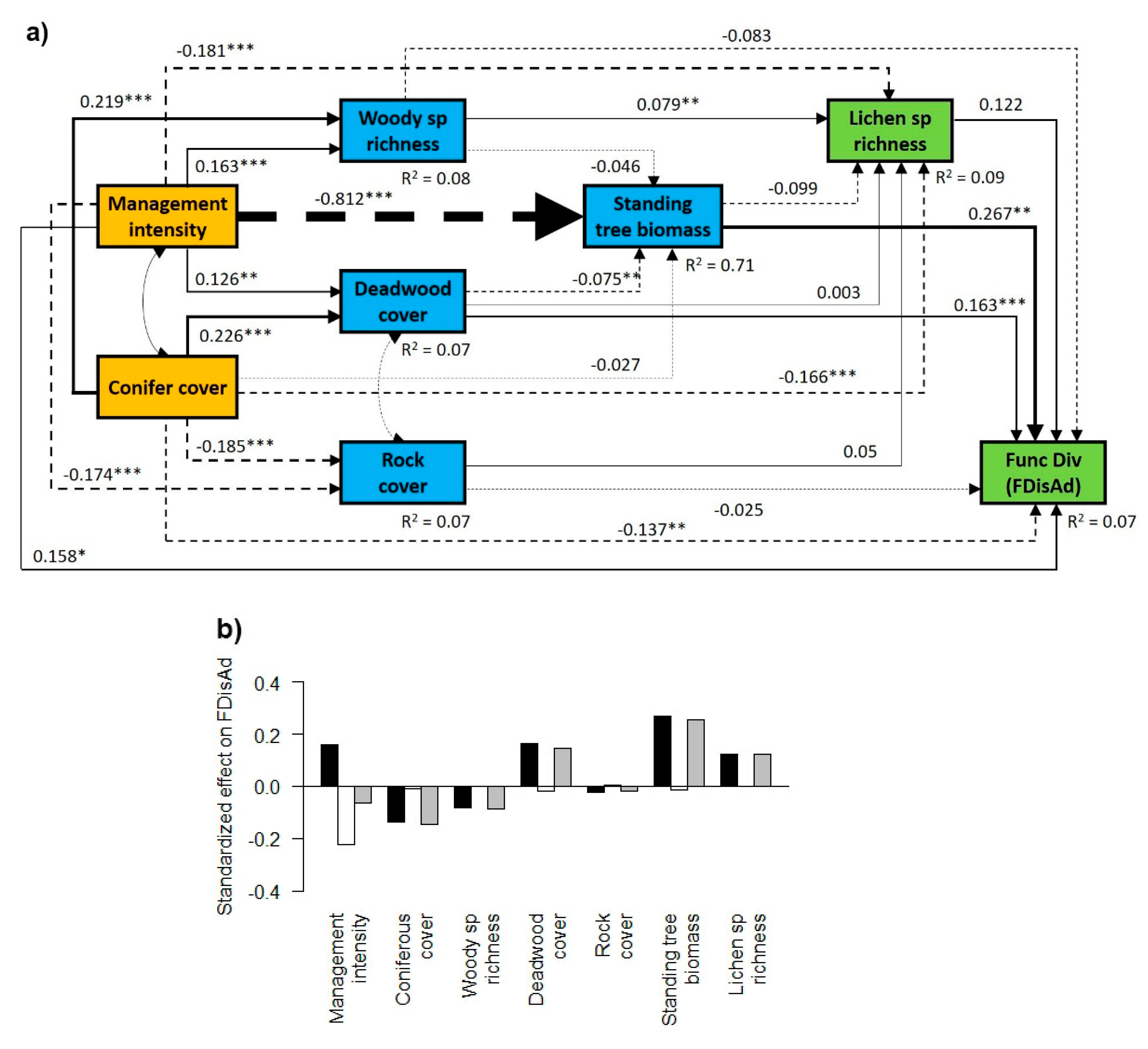
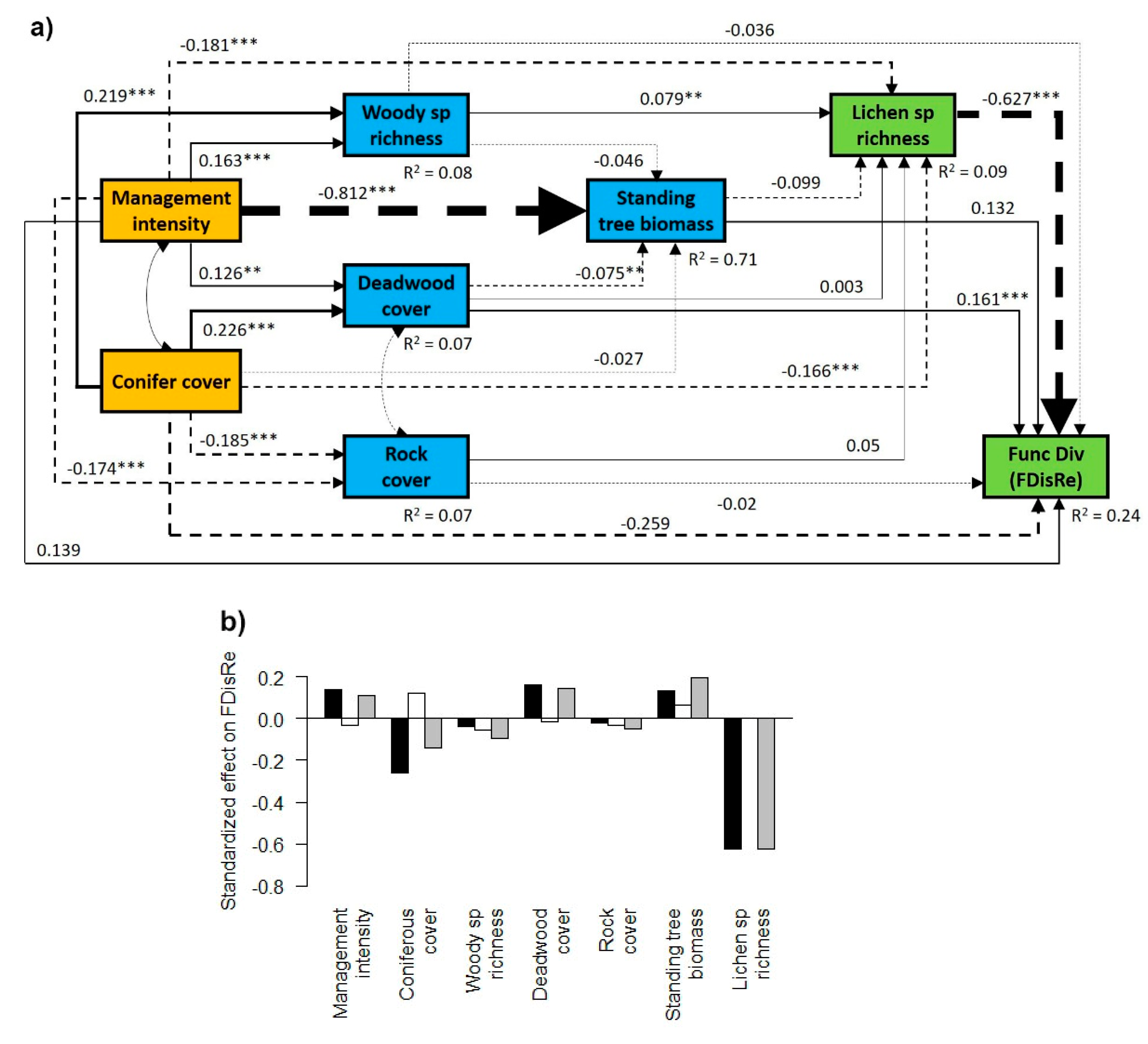
3.1. General Effects of Forest Management Intensity and Ecosystem Components on Lichen Functional Diversity
3.2. Contrasting Findings for FDisTotal, FDisAd and FDisRe
4. Discussion
4.1. Direct Effects of Management Intensity on Environmental Variables and Related Effects on Lichen Species Richness
4.2. Direct and Indirect Effect of Management Intensity and Environmental Variables on Lichen Functional Diversity
4.3. Implications for Forest Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nascimbene, J.; Martellos, S.; Nimis, P.L. Epiphytic lichens of tree-line forests in the central-eastern Italian Alps and their importance for conservation. Lichenologist 2006, 38, 373–382. [Google Scholar] [CrossRef]
- Hauck, M.; de Bruyn, U.; Leuschner, C. Dramatic diversity losses in epiphytic lichens in temperate broad-leaved forests during the last 150 years. Biol. Conserv. 2013, 157, 136–145. [Google Scholar] [CrossRef]
- Nascimbene, J.; Nimis, P.L.; Dainese, M. Epiphytic lichen conservation in the Italian Alps: The role of forest type. Fungal Ecol. 2014, 11, 164–172. [Google Scholar] [CrossRef]
- Łubek, A.; Kukwa, M.; Jaroszewicz, B.; Czortek, P. Identifying mechanisms shaping lichen functional diversity in a primeval forest. For. Ecol. Manag. 2020, 475, 118434. [Google Scholar] [CrossRef]
- Zedda, L.; Rambold, G. The Diversity of Lichenised Fungi: Ecosystem Functions and Ecosystem Services. In Recent Advances in Lichenology: Modern Methods and Approaches in Lichen Systematics and Culture Techniques; Upreti, D.K., Divakar, P.K., Shukla, V., Bajpai, R., Eds.; Springer: New Delhi, India, 2015; Volume 2, pp. 121–145. [Google Scholar]
- Scheidegger, C.; Werth, S. Conservation strategies for lichens: Insights from population biology. Fungal Biol. Rev. 2009, 23, 55ss66. [Google Scholar] [CrossRef]
- Ludwig, G.; Matzke-Hajek, G. Rote Liste gefährdeter Tiere, Pflanzen und Pilze Deutschlands. Band 6: Pilze (Teil 2)—Flechten und Myxomyzeten—Naturschutz und Biologische Vielfalt 70; Landwirtschaftsverlag: Münster, Germany, 2011; p. 240. [Google Scholar]
- Parviainen, J.; Bozzano, M.; Estreguil, C.; Koskela, J.; Lier, M.; Vogt, P.; Ostapowicz, K. Maintenance, conservation and appropriate enhancement of biological diversity in forest ecosystems. In State of Europe’s Forests 2007—MCPFE Report on Sustainable Forest Management in Europe; Köhl, M., Rametsteiner, E., Eds.; MCPFE, Liaison Unit: Warsaw, Poland, 2007; pp. 45–72. [Google Scholar]
- BMELV (Bundesministerium für Ernährung, Landwirtschaft und Verbraucherschutz). Ergebnisse der Bundeswaldinventur 2012; Druck- und Verlagshaus Zarbock GmbH & Co. KG: Frankfurt am Main, Germany, 2012; p. 280. [Google Scholar]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Busch, V.; Klaus, V.H.; Penone, C.; Schäfer, D.; Boch, S.; Prati, D.; Müller, J.; Socher, S.A.; Niinemets, Ü.; Peñuelas, J.; et al. Nutrient stoichiometry and land use rather than species richness determine plant functional diversity. Ecol. Evol. 2018, 8, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Boeddinghaus, R.; Marhan, S.; Berner, D.; Boch, S.; Fischer, M.; Hoelzel, N.; Kattge, J.; Klaus, V.H.; Kleinebecker, T.; Oelmann, Y.; et al. Plant functional trait shifts explain concurrent changes in the structure and function of grassland soil microbial communities. J. Ecol. 2019, 107, 2197–2210. [Google Scholar] [CrossRef] [Green Version]
- Laureto, L.M.O.; Cianciaruso, M.V.; Samia, D.S.M. Functional diversity: An overview of its history and applicability. Nat. Conserv. 2015, 13, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Lavorel, S.; McIntyre, S.; Falcazuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.; Sternberg, M.; Noy-meir, I.; et al. Plant trait responses to grazing? A global synthesis. Glob. Chang. Biol. 2007, 13, 313–341. [Google Scholar] [CrossRef]
- Hillebrand, H.; Matthiessen, B. Biodiversity in a complex world: Consolidation and progress in functional biodiversity research. Ecol. Lett. 2009, 12, 1405–1419. [Google Scholar] [CrossRef]
- Stofer, S.; Bergamini, A.; Aragón, G.; Carvalho, P.; Coppins, B.J.; Davey, S.; Dietrich, M.; Farkas, E.; Kärkkäinen, K.; Keller, C.; et al. Species richness of lichen functional groups in relation to land use intensity. Lichenologist 2006, 38, 331–353. [Google Scholar] [CrossRef] [Green Version]
- Giordani, P.; Brunialti, G.; Bacaro, G.; Nascimbene, J. Functional traits of epiphytic lichens as potential indicators of environmental conditions in forest ecosystems. Ecol. Indic. 2012, 18, 413–420. [Google Scholar] [CrossRef]
- Malíček, J.; Palice, Z.; Vondrák, J.; Kostovčík, M.; Lenzová, V.; Hofmeister, J. Lichens in old-growth and managed mountain spruce forests in the Czech Republic: Assessment of biodiversity, functional traits and bioindicators. Biodivers. Conserv. 2019, 28, 3497–3528. [Google Scholar] [CrossRef]
- Wan, S.; Ellis, C. Are lichen growth form categories supported by continuous functional traits: Water-holding capacity and specific thallus mass? Edinb. J. Bot. 2019, 77, 65–76. [Google Scholar] [CrossRef]
- Nimis, P.L.; Martellos, S.; Chiarucci, A.; Ongaro, S.; Peplis, M.; Pittao, E.; Nascimbene, J. Exploring the relationships between ecology and species traits in cyanolichens: A case study on Italy. Fungal Ecol. 2020, 47, 100950. [Google Scholar] [CrossRef]
- Giordani, P.; Malaspina, P.; Benesperi, R.; Incerti, G.; Nascimbene, J. Functional over-redundancy and vulnerability of lichen communities decouple across spatial scales and environmental severity. Sci. Total Environ. 2019, 666, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, P.; Prieto, M.; Martínez-Vilalta, J.; Giordani, P.; Aragón, G.; López-Angulo, J.; Košuthová, A.; Merinero, S.; Díaz-Peña, E.M.; Rosas, T.; et al. Disentangling functional trait variation and covariation in epiphytic lichens along a continent-wide latitudinal gradient. Proc. Royal Soc. B 2020, 287, 20192862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurtado, P.; Prieto, M.; Aragón, G.; Escudero, A.; Martínez, I. Critical predictors of functional, phylogenetic and taxonomic diversity are geographically structured in lichen epiphytic communities. J. Ecol. 2019, 107, 2303–2316. [Google Scholar] [CrossRef]
- Bässler, C.; Cadotte, M.W.; Beudert, B.; Heibl, C.; Blaschke, M.; Bradtka, J.H.; Langbehn, T.; Werth, S.; Müller, J. Contrasting patterns of lichen functional diversity and species richness across an elevation gradient. Ecography 2016, 39, 689–698. [Google Scholar] [CrossRef]
- Lelli, C.; Bruun, H.H.; Chiarucci, A.; Donati, D.; Frascaroli, F.; Fritz, Ö.; Goldberg, I.; Nascimbene, J.; Tøttrup, A.P.; Rahbek, C.; et al. Biodiversity response to forest structure and management: Comparing species richness, conservation relevant species and functional diversity as metrics in forest conservation. For. Ecol. Manag. 2019, 432, 707–717. [Google Scholar] [CrossRef]
- Flynn, D.F.B.; Gogol-Prokurat, M.; Nogeire, T.; Molinari, N.; Trautman Richers, B.; Lin, B.B.; Simpson, N.; Mayfield, M.M.; DeClerck, F. Loss of functional diversity under land use intensification across multiple taxa. Ecol. Lett. 2009, 12, 22–33. [Google Scholar] [CrossRef]
- Clavel, R.; Julliard, R.; Devictor, V. Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 2011, 9, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Gámez-Virués, S.; Perović, D.J.; Gossner, M.M.; Börschig, C.; Blüthgen, N.; de Jong, H.; Simons, N.K.; Klein, A.M.; Krauss, J.; Maier, G.; et al. Landscape simplification filters species traits and drives biotic homogenization. Nat. Commun. 2015, 6, 8568. [Google Scholar] [CrossRef]
- Werth, S.; Wagner, H.H.; Gugerli, F.; Holderegger, R.; Csencsics, D.; Kalwij, J.M.; Scheidegger, C. Quantifying dispersal and establishment limitation in a population of an epiphytic lichen. Ecology 2006, 87, 2037–2046. [Google Scholar] [CrossRef] [Green Version]
- Lesica, P.; McCune, B.; Cooper, S.V.; Hong, W.S. Differences in lichen and bryophyte communities between old-growth and managed second-growth forests in the Swan Valley, Montana. Can. J. Bot. 1991, 69, 1745–1755. [Google Scholar] [CrossRef]
- Bergamini, A.; Scheidegger, C.; Stofer, S.; Carvalho, P.; Davey, S.; Dietrich, M.; Dubs, F.; Farkas, E.; Groner, U.; Kärkkäinen, K.; et al. Performance of macrolichens and lichen genera as indicators of lichen species richness and composition. Conserv. Biol. 2005, 19, 1051–1062. [Google Scholar] [CrossRef]
- Rudolphi, J.; Gustafsson, L. Forests regenerating after clear-cutting function as habitat for bryophyte and lichen species of conservation concern. PLoS ONE 2011, 6, e18639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paillet, Y.; Bergès, L.; Hjältén, J.; Ódor, P.; Avon, C.; Bernhardt-Römermann, M.; Bijlsma, R.-J.; de Bruyn, L.; Fuhr, M.; Grandin, U.; et al. Biodiversity differences between managed and unmanaged forests: Meta-analysis of species richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef]
- Friedel, A.; von Oheimb, G.; Dengler, J.; Härdtle, W. Species diversity and species composition of epiphytic bryophytes and lichens—a comparison of managed and unmanaged beech forests in NE Germany. Feddes Repert. 2006, 117, 172–185. [Google Scholar] [CrossRef]
- Fritz, Ö.; Gustafsson, L.; Larsson, K. Does forest continuity matter in conservation?—A study of epiphytic lichens and bryophytes in beech forests of southern Sweden. Biol. Conserv. 2008, 141, 655–668. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L.; Nimis, P.L. Influence of forest management on epiphytic lichens in a temperate beech forest of northern Italy. For. Ecol. Manag. 2007, 247, 43–47. [Google Scholar] [CrossRef]
- Fritz, Ö.; Caldiz, M.S.; Brunet, J. Interacting effects of tree characteristics on the occurrence of rare epiphytes in a Swedish beech forest area. Bryologist 2009, 112, 488–505. [Google Scholar] [CrossRef]
- Fritz, Ö.; Niklasson, M.; Churski, M. Tree age is a key factor for the conservation of epiphytic lichens and bryophytes in beech forests. Appl. Veg. Sci. 2009, 12, 93–106. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L.; Nimis, P.L. Epiphytic lichen diversity in old-growth and managed Picea abies stands in Alpine spruce forests. For. Ecol. Manag. 2010, 260, 603–609. [Google Scholar] [CrossRef]
- Schall, P.; Gossner, M.M.; Heinrichs, S.; Fischer, M.; Boch, S.; Prati, D.; Jung, K.; Baumgartner, V.; Blaser, S.; Böhm, S.; et al. The impact of even-aged and unevenaged forest management on regional biodiversity of multiple taxa in European beech forests. J. Appl. Ecol. 2018, 55, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Boch, S.; Prati, D.; Hessenmöller, D.; Schulze, E.-D.; Fischer, M. Richness of lichen species, especially of threatened ones, is promoted by management methods furthering stand continuity. PLoS ONE 2013, 8, e55461. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M.; Bossdorf, O.; Gockel, S.; Hänsel, F.; Hemp, A.; Hessenmöller, D.; Korte, G.; Nieschulze, J.; Pfeiffer, S.; Prati, D.; et al. Implementing large-scale and long-term functional biodiversity research: The Biodiversity Exploratories. Basic Appl. Ecol. 2010, 11, 473–485. [Google Scholar] [CrossRef]
- Grace, J.B.; Anderson, T.M.; Olff, H.; Scheiner, S.M. On the specification of structural equation models for ecological systems. Ecol. Monogr. 2010, 80, 67–87. [Google Scholar] [CrossRef] [Green Version]
- Boch, S.; Prati, D.; Müller, J.; Socher, S.A.; Baumbach, H.; Buscot, F.; Gockel, S.; Hemp, A.; Hessenmöller, D.; Kalko, E.K.V.; et al. High plant species richness indicates management-related disturbances, rather than the conservation status of forests. Basic Appl. Ecol. 2013, 14, 496–505. [Google Scholar] [CrossRef]
- Schall, P.; Ammer, C. How to quantify forest management intensity in Central European forests. Eur. J. Forest Res. 2013, 132, 379–396. [Google Scholar] [CrossRef] [Green Version]
- Blaser, S.; Prati, D.; Senn-Irlet, B.; Fischer, M. Effects of forest management on the diversity of deadwood-inhabiting fungi in Central European forests. For. Ecol. Manag. 2013, 304, 42–48. [Google Scholar] [CrossRef]
- Humphrey, J.W.; Davey, S.; Peace, A.J.; Ferris, R.; Harding, K. Lichens and bryophyte communities of planted and semi-natural forests in Britain: The influence of site type, stand structure and dead wood. Biol. Conserv. 2002, 107, 165–180. [Google Scholar] [CrossRef]
- Neitlich, P.N.; McCune, B. Hotspots of epiphytic lichen diversity in two young managed forests. Conserv. Biol. 1997, 11, 172–182. [Google Scholar] [CrossRef]
- Hessenmöller, D.; Nieschulze, J.; von Lüpke, N.; Schulze, E.-D. Identification of forest management types from ground-based and remotely sensed variables and the effects of forest management on forest structure and composition. Forstarchiv 2011, 82, 171–183. [Google Scholar]
- Müller, J.; Boch, S.; Prati, D.; Socher, S.A.; Pommer, U.; Hessenmöller, D.; Schall, P.; Schulze, E.-D.; Fischer, M. Effects of forest management on bryophyte species richness in Central European forests. For. Ecol. Manag. 2019, 432, 850–859. [Google Scholar] [CrossRef]
- Moning, C.; Werth, S.; Dziock, F.; Bässler, C.; Bradtka, J.; Hothorn, T.; Müller, J. Lichen diversity in temperate montane forests is influenced by forest structure more than climate. Basic Appl. Ecol. 2009, 258, 745–751. [Google Scholar] [CrossRef]
- Ardelean, I.V.; Keller, C.; Scheidegger, C. Effects of management on lichen species richness, ecological traits and community structure in the Rodnei Mountains National Park (Romania). PLoS ONE 2015, 10, e0145808. [Google Scholar]
- Ames, S.; Pischke, K.; Schoenfuss, N.; Snobl, Z.; Soine, J.; Weiher, E.; Wellnitz, T. Biogeographic patterns of lichens and trees on islands of the Boundary Waters Canoe Area Wilderness. BIOS J. 2012, 83, 145–154. [Google Scholar] [CrossRef]
- Mežaka, A.; Brūmelis, G.; Piterāns, A. Tree and stand-scale factors affecting richness and composition of epiphytic bryophytes and lichens in deciduous woodland key habitats. Biodivers. Conserv. 2012, 21, 3221–3241. [Google Scholar] [CrossRef]
- Saiz, H.; Dainese, M.; Chiarucci, A.; Nascimbene, J. Networks of epiphytic lichens and host trees along elevation gradients: Climate change implications in mountain ranges. J. Ecol. in press.
- Schall, P.; Heinrichs, S.; Ammer, C.; Ayasse, M.; Boch, S.; Buscot, F.; Fischer, M.; Goldmann, K.; Overmann, J.; Schulze, E.-D.; et al. Can multi-taxa diversity in European beech forest landscapes be increased by combining different management systems? J. Appl. Ecol. 2020, 57, 1363–1375. [Google Scholar] [CrossRef] [Green Version]
- Boch, S.; Müller, J.; Prati, D.; Blaser, S.; Fischer, M. Up in the tree—the overlooked richness of bryophytes and lichens in tree crowns. PLoS ONE 2013, 8, e84913. [Google Scholar] [CrossRef] [Green Version]
- Kiebacher, T.; Keller, C.; Scheidegger, C.; Bergamini, A. Hidden crown jewels: The role of tree crowns for bryophyte and lichen species richness in sycamore maple wooded pastures. Biodivers. Conserv. 2016, 25, 1605–1624. [Google Scholar] [CrossRef]
- Scholz, P. Katalog der Flechten und flechtenbewohnenden Pilze Deutschlands. Schriftenr. Vegetationskd. 2000, 31, 1–298. [Google Scholar]
- Smith, C.W.; Aptroot, A.; Coppins, B.J.; Fletcher, A.; Gilbert, O.L.; James, P.W.; Wolseley, P.A. The Lichens of Great Britain and Ireland, 2nd ed.; British Lichen Society: London, UK, 2009; p. 1046. [Google Scholar]
- Wirth, V.; Hauck, M.; Schulz, M. Die Flechten Deutschlands; Eugen Ulmer: Stuttgart, Germany, 2013; Volumes 1–2, p. 1244. [Google Scholar]
- CNALH (Consortium of North American Lichen Herbaria). Available online: https://lichenportal.org/cnalh/ (accessed on 24 June 2020).
- Rambold, G.; Davydov, E.; Elix, J.A.; Nash, T.H., III; Scheidegger, C.; Zedda, L. LIAS Light—A Database for Rapid Identification of Lichens. 2020. Available online: http://liaslight.lias.net/ (accessed on 11 May 2020).
- Wirth, V. Ökologische Zeigerwerte von Flechten. Herzogia 2010, 23, 229–248. [Google Scholar] [CrossRef]
- Sparrius, L.B.; van den Top, G.G.; van Swaay, C.A.M. An approach to calculate a species temperature index for flora based on open data. Gorteria 2018, 40, 73–78. [Google Scholar]
- Wamelink, G.W.W.; Joosten, V.; van Dobben, H.F.; Berendse, F. Validity of Ellenberg indicator values judged from physico-chemical field measurements. J. Veg. Sci. 2002, 13, 269–278. [Google Scholar] [CrossRef]
- Diekmann, M. Species indicator values as an important tool in applied plant ecology—A review. Basic Appl. Ecol. 2003, 4, 493–506. [Google Scholar] [CrossRef]
- Boch, S.; Kurtogullari, Y.; Allan, E.; Lessard-Therrien, M.; Rieder, N.S.; Fischer, M.; Martínez De León, G.; Arlettaz, R.; Humbert, J.-Y. Effects of fertilization and irrigation on vascular plant species richness, functional composition and yield in mountain grasslands. J. Environ. Manag. 2021, 279, 111629. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology; R Package Version 1.0-12; 2014. Available online: https://mran.microsoft.com/snapshot/2014-11-17/web/packages/FD/FD.pdf (accessed on 19 August 2020).
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 623–637. [Google Scholar] [CrossRef]
- Mason, N.W.H.; De Bello, F.; Mouillot, D.; Pavoine, S.; Dray, S.A. Guide for using functional diversity indices to reveal changes in assembly processes along ecological gradients. J. Veg. Sci. 2013, 24, 794–806. [Google Scholar] [CrossRef]
- Boch, S.; Allan, E.; Humbert, J.-Y.; Kurtogullari, Y.; Lessard-Therrien, M.; Müller, J.; Prati, D.; Rieder, N.S.; Arlettaz, R.; Fischer, M. Direct and indirect effects of land use on bryophytes in grasslands. Sci. Total Environ. 2018, 644, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Frisch, A.; Rudolphi, J.; Sheil, D.; Caruso, A.; Thor, G.; Gustafsson, L. Tree Species Composition predicts epiphytic lichen communities in an African montane rain forest. Biotropica 2015, 47, 542–549. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Rosseel, Y.; Oberski, D.; Byrnes, J.; Vanbrabant, L.; Savalei, V.; Merkle, E.; Hallquist, M.; Rhemtulla, M.; Katsikatsou, M.; Barendse, M.; et al. Package ‘lavaan’. 2017. Available online: https://cran.microsoft.com/snapshot/2017-04-09/web/packages/lavaan/lavaan.pdf (accessed on 5 December 2020).
- Müller, J.; Boch, S.; Blaser, S.; Fischer, M.; Prati, D. Effects of forest management on bryophyte communities on deadwood. Nova Hedwigia 2015, 100, 423–438. [Google Scholar] [CrossRef]
- Ellis, C.J. Lichen epiphyte diversity: A species, community and trait-based review. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 131–152. [Google Scholar] [CrossRef]
- Zellweger, F.; Coomes, D.; Lenoir, J.; Depauw, L.; Maes, S.L.; Wulf, M.; Kirby, K.J.; Brunet, J.; Kopecký, V.; Máliš, F.; et al. Seasonal drivers of understory temperature buffering in temperate deciduous forests across Europe. Glob. Ecol. Biogeogr. 2019, 28, 1774–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormann, C.F.; Bagnara, M.; Boch, S.; Hinderling, J.; Janeiro‑Otero, A.; Schäfer, D.; Schall, P.; Hartig, F. Plant species richness increases with light availability, but not variability, in temperate forests understorey. BMC Ecol. 2020, 20, 43. [Google Scholar] [CrossRef]
- Rambo, T.R.; North, M.P. Canopy microclimate response to pattern and density of thinning in a Sierra Nevada forest. For. Ecol. Manag. 2009, 257, 435–442. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Solhaug, K.A. Differences in the susceptibility to light stress between epiphytic lichens of ancient and young boreal forest stands. Funct. Ecol. 1996, 10, 344–354. [Google Scholar] [CrossRef]
- McGee, G.G.; Kimmerer, R.W. Forest age and management effects on epiphytic bryophyte communities in Adirondack northern hardwood forests, New York, U.S.A. Can. J. For. Res. 2002, 32, 1562–1576. [Google Scholar] [CrossRef]
- Bardat, J.; Aubert, M. Impact of forest management on the diversity of corticolous bryophyte assemblages in temperate forests. Biol. Cons. 2007, 139, 47–66. [Google Scholar] [CrossRef]
- Sillett, S.C.; Antoine, M.E. Lichens and bryophytes in forest canopies. In Forest Canopies; Lowman, M., Rinker, H.B., Eds.; Academic Press: Burlington, NJ, USA, 2004; pp. 151–173. [Google Scholar]
- Király, I.; Ódor, P. The effect of stand structure and tree species composition on epiphytic bryophytes in mixed deciduous–coniferous forests of Western Hungary. Biol. Conserv. 2010, 143, 2063–2069. [Google Scholar] [CrossRef]
- Nordén, B.; Dahlberg, A.; Brandrud, T.E.; Fritz, Ö.; Ejrnaes, R.; Ovaskainen, O. Effects of ecological continuity on species richness and composition in forests and woodlands: A review. Ecoscience 2014, 21, 34–45. [Google Scholar] [CrossRef]
- Schall, P.; Heinrichs, S.; Ammer, C.; Ayasse, M.; Boch, S.; Buscot, F.; Fischer, M.; Goldmann, K.; Overmann, J.; Schulze, E.-D.; et al. Heterogeneity among different developmental phases is key for forest biodiversity in managed beech forests, but this does not question the general value of unmanaged forests: Response to Bruun & Heilmann Clausen (2020). J. Appl. Ecol. 2020, in press. [Google Scholar]
- Barkman, J.J. Phytosociology and Ecology of Cryptogamic Epiphytes; Van Gorcum: Assen, The Netherlands, 1958; p. 628. [Google Scholar]
- Fengel, D.; Wegener, G. Wood: Chemistry, Ultrastructure, Reactions; Walter de Gruyter: Berlin, Germany, 1984; p. 613. [Google Scholar]
- Caruso, A.; Rudolphi, J.; Thor, G. Lichen species diversity and substrate amounts in young planted boreal forests: A comparison between slash and stumps of Picea abies. Biol. Conserv. 2008, 141, 47–55. [Google Scholar] [CrossRef]
- Sabatini, F.; Burrascano, S.; Keeton, W.S.; Levers, C.; Lindner, M.; Pötzschner, F.; Verkerk, P.J.; Bauhus, J.; Buchwald, E.; Chaskovsky, O.; et al. Where are Europe’s last primary forests? Divers. Distrib. 2018, 24, 1426–1439. [Google Scholar] [CrossRef] [Green Version]
- Marini, L.; Nascimbene, J.; Nimis, P.L. Large-scale patterns of epiphytic lichen species richness: Photobiont-dependent response to climate and forest structure. Sci. Total Environ. 2011, 409, 4381–4386. [Google Scholar] [CrossRef]
- Rundel, P.W. The ecological role of secondary substances. Biochem. Syst. Ecol. 1978, 6, 157–170. [Google Scholar] [CrossRef]
- Boch, S.; Martins, A.; Ruas, S.; Fontinha, S.; Carvalho, P.; Reis, F.; Bergamini, A.; Sim-Sim, M. Bryophyte and macrolichen diversity show contrasting elevation relationships and are negatively affected by disturbances in laurel forests of Madeira island. J. Veg. Sci. 2019, 30, 1122–1133. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Lie, M.; Solhaug, K.A.; Ohlson, M. Growth and ecophysiological acclimation of the foliose lichen Lobaria pulmonaria in forests with contrasting light climates. Oecologia 2006, 147, 406–416. [Google Scholar] [CrossRef]
- Büdel, B.; Scheidegger, C. Thallus morphology and anatomy. In Lichen Biology, 2nd ed.; Nash, T.H., III, Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 40–68. [Google Scholar]
- Király, I.; Nascimbene, J.; Tinya, F.; Ódor, P. Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodivers. Conserv. 2013, 22, 209–223. [Google Scholar] [CrossRef] [Green Version]
- Raevel, V.; Violle, C.; Munoz, F. Mechanisms of ecological succession: Insights from plant functional strategies. Oikos 2012, 121, 1761–1770. [Google Scholar] [CrossRef]
- Sanders, W.B.; Lücking, R. Reproductive strategies, relichenization and thallus development observed in situ in leaf-dwelling lichen communities. New Phytol. 2002, 155, 425–435. [Google Scholar] [CrossRef]
- Johansson, V.; Snäll, T.; Ranius, T. Epiphyte metapopulation dynamics are explained by species traits, connectivity and patch dynamics. Ecology 2012, 93, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Ranius, T.; Johansson, P.; Berg, N.; Niklasson, M. The influence of forest age and microhabitat quality on the occurrence of crustose lichens associated with old oaks. J. Veg. Sci. 2008, 19, 653–662. [Google Scholar] [CrossRef]
- Fritz, Ö.; Heilmann-Clausen, J. Rot holes create key microhabitats for epiphytic lichens and bryophytes on beech (Fagus sylvatica). Biol. Conserv. 2010, 143, 1008–1016. [Google Scholar] [CrossRef]
- Pentecost, A. Some observations on the size and shape of lichen ascospores in relation to ecology and taxonomy. New Phytol. 1981, 89, 667–678. [Google Scholar] [CrossRef]
- Gustafsson, L.; Appelgren, L.; Jonsson, F.; Nordin, U.; Persson, A.; Weslien, J.-O. High occurrence of red-listed bryophytes and lichens in mature managed forests in boreal Sweden. Basic Appl. Ecol. 2004, 5, 123–129. [Google Scholar] [CrossRef]
- Moning, C.; Müller, J. Critical forest age thresholds for the diversity of lichens, molluscs and birds in beech (Fagus sylvatica L.) dominated forests. Ecol. Ind. 2009, 9, 922–932. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Schwäbische Alb | Hainich-Dün | Schorfheide-Chorin | |
|---|---|---|---|
| Location | SW Germany | Central Germany | NE Germany |
| Size | ~422 km2 | ~1300 km2 | ~1300 km2 |
| Geology | Calcareous bedrock | Calcareous bedrock | Young glacial landscape |
| Elevation a.s.l. | 460–860 m | 285–550 m | 3–140 m |
| Annual mean temperature | 6–7 °C | 6.5–8 °C | 8–8.5 °C |
| Annual mean precipitation | 700–1000 mm | 500–800 mm | 500–600 mm |
| N plots | 158 | 175 | 309 |
| Lichen species richness | |||
| Mean (se) | 18.61 (0.7) | 5.0 (0.2) | 5.9 (0.2) |
| Range | 3–54 | 0–15 | 0–17 |
| FDisTotal | |||
| Mean (se) | 0.3 (<0.01) | 0.3 (<0.01) | 0.3 (<0.01) |
| Range | 0.2–0.5 | 0.2–0.5 | 0.2–0.5 |
| FDisAd | |||
| Mean (se) | 0.2 (<0.01) | 0.3 (<0.01) | 0.3 (<0.01) |
| Range | 0.1–0.5 | 0.1–0.4 | 0.1–0.5 |
| FDisRe | |||
| Mean (se) | 0.3 (<0.01) | 0.3 (<0.01) | 0.4 (<0.01) |
| Range | 0.2–0.4 | 0.2–0.5 | 0.2–0.5 |
| Standing tree biomass [m3/ha] | |||
| Mean (se) | 323.9 (15.8) | 400.1 (14.9) | 441.9 (10.8) |
| Range | 0–1017 | 0–882 | 0–1002 |
| SMId | |||
| Mean (se) | 0.5 (<0.1) | 0.4 (<0.1) | 0.3 (<0.1) |
| Range | 0–1 | 0–1 | 0–1 |
| Deadwood cover [%] | |||
| Mean (se) | 3.7 (0.2) | 3.1 (0.2) | 3.8 (0.2) |
| Range | 0.5–20.0 | 0.5–15.0 | 0.5–25.0 |
| Rock cover [%] | |||
| Mean (se) | 1.0 (0.2) | 0.2 (<0.1) | 0.3 (<0.1) |
| Range | 0–11 | 0–4 | 0–6 |
| Proportional conifer cover | |||
| Mean (se) | 18.6 (3.2) | 3.4 (1.1) | 23.1 (1.9) |
| Range | 0–100 | 0–91 | 0–100 |
| Woody species richness | |||
| Mean (se) | 7.5 (0.3) | 6.8 (0.2) | 5.3 (0.2) |
| Range | 1–16 | 1–14 | 1–16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boch, S.; Saiz, H.; Allan, E.; Schall, P.; Prati, D.; Schulze, E.-D.; Hessenmöller, D.; Sparrius, L.B.; Fischer, M. Direct and Indirect Effects of Management Intensity and Environmental Factors on the Functional Diversity of Lichens in Central European Forests. Microorganisms 2021, 9, 463. https://doi.org/10.3390/microorganisms9020463
Boch S, Saiz H, Allan E, Schall P, Prati D, Schulze E-D, Hessenmöller D, Sparrius LB, Fischer M. Direct and Indirect Effects of Management Intensity and Environmental Factors on the Functional Diversity of Lichens in Central European Forests. Microorganisms. 2021; 9(2):463. https://doi.org/10.3390/microorganisms9020463
Chicago/Turabian StyleBoch, Steffen, Hugo Saiz, Eric Allan, Peter Schall, Daniel Prati, Ernst-Detlef Schulze, Dominik Hessenmöller, Laurens B. Sparrius, and Markus Fischer. 2021. "Direct and Indirect Effects of Management Intensity and Environmental Factors on the Functional Diversity of Lichens in Central European Forests" Microorganisms 9, no. 2: 463. https://doi.org/10.3390/microorganisms9020463
APA StyleBoch, S., Saiz, H., Allan, E., Schall, P., Prati, D., Schulze, E.-D., Hessenmöller, D., Sparrius, L. B., & Fischer, M. (2021). Direct and Indirect Effects of Management Intensity and Environmental Factors on the Functional Diversity of Lichens in Central European Forests. Microorganisms, 9(2), 463. https://doi.org/10.3390/microorganisms9020463