
Interactions of the Streptococcus pneumoniae Toxin-Antitoxin RelBE Proteins with Their Target DNA


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Transformation, and Growth Conditions
2.2. Plasmid Constructions
- -
- pMP220-PrelBE (10,694 bp) carry a 210-bp PCR DNA fragment with the promoter region of the pneumococcal relBE operon. The primers used were: relBE-184E (5′-CGGGATCCGAACTGGCTCATATGACCATGG-3′) and relBE+28K (5′-CGGGGTACCCATCTTTTGTGTCCCTTTTTTAATG-3′).
- -
- pMP220-relBE (11,190 bp) carry a 706-bp DNA fragment containing the promoter and the entire relBE operon. The primers used were: relBE-184E (5′-CGGGATCCGAACTGGCTCATATGACCATGG-3′) and relECK (5′-CGGGGTACCTCAATAAATATCTCTCCGATGACC-3′).
- -
- pMP220-relB (10,934 bp) carry the promoter region and gene relB in a DNA fragment of 450 bp. Primers used were: relBE-184E (5′-CGGGATCCGAACTGGCTCATATGACCATGG-3′) and relB2CK (5′-CGGGGTACCTTATTCATCCTTCAAGCCTAAATC-3′).
2.3. β-Galactosidase Activity Measurements
2.4. Overexpression and Purification of Pneumococcal Proteins RelB-RelE and RelB
2.5. DNase I and Hydroxyl Radical Footprinting Experiments
2.6. Bioinformatics Analyses
3. Results
3.1. The relBE Operon of Streptococcus pneumoniae

3.2. Distribution of the Pneumococcal Toxin RelE


3.3. The Pneumococcal RelB Protein Represses Transcription of the relBE Operon and RelE Acts as an Efficient Co-Repressor

3.4. Determination of DNA Binding Sites of RelB Protein and the RelB:RelE Protein Complex
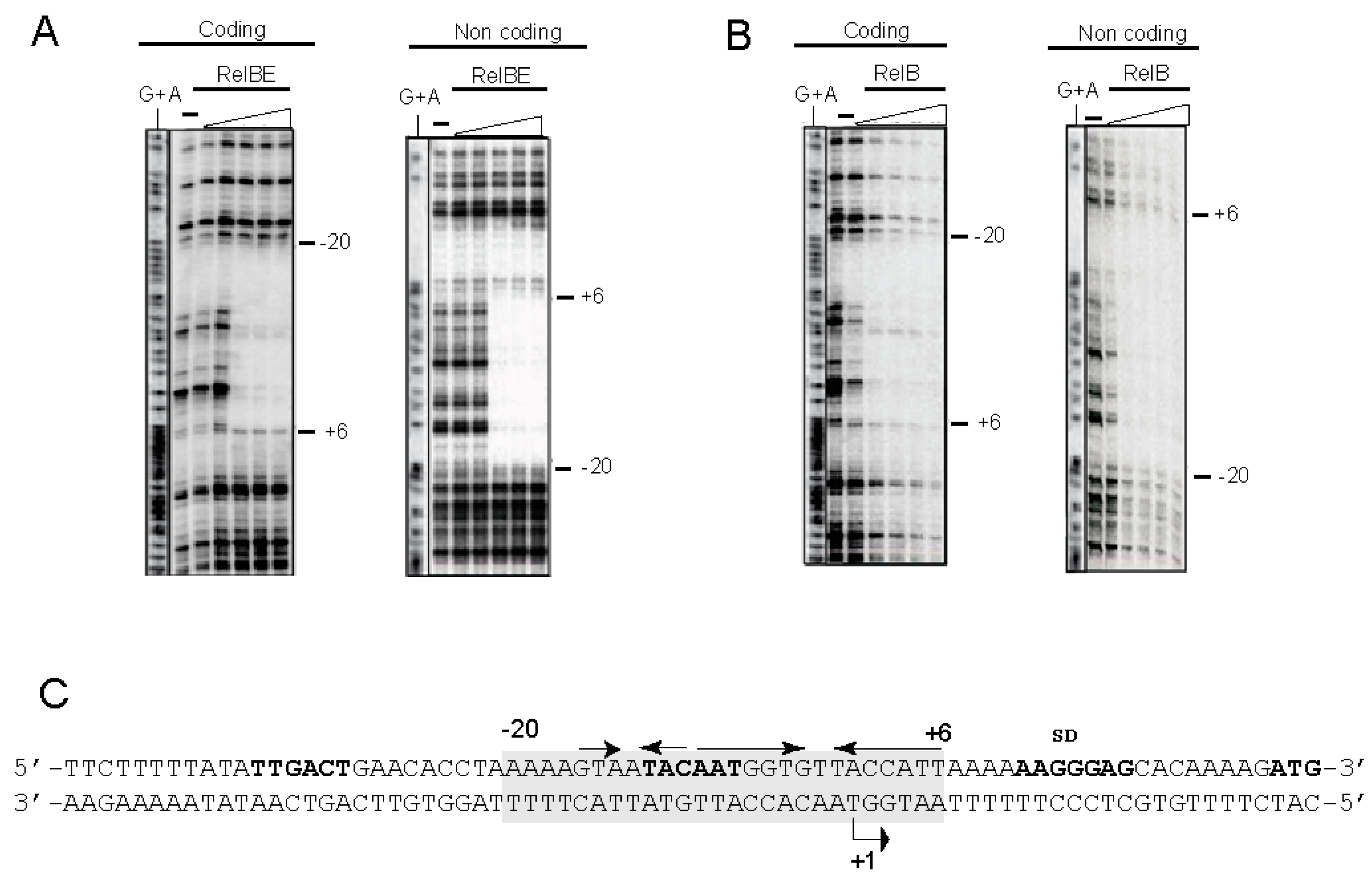
3.5. High-Resolution Footprinting Assays

4. Discussion

Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, W.T.; Moreno-Córdoba, I.; Yeo, C.C.; Espinosa, M. Toxin-Antitoxin Genes of the Gram-Positive Pathogen Streptococcus pneumoniae: So Few and Yet So Many. Microbiol. Mol. Biol. Rev. 2012, 76, 773–791. [Google Scholar] [CrossRef] [Green Version]
- Hayes, F.; van Melderen, L. Toxins-antitoxins: Diversity, evolution and function. Crit. Rev. Biochem. Mol. Biol. 2011, 46, 386–408. [Google Scholar] [CrossRef]
- Pandey, D.P.; Gerdes, K. Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucl. Acids Res. 2005, 33, 966–976. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, K.; Christensen, K.S.; Lobner-Olensen, A. Prokaryotic toxin-antitoxin stress response loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef]
- Muthuramalingam, M.; White, J.C.; Bourne, C.R. Toxin-Antitoxin Modules Are Pliable Switches Activated by Multiple Protease Pathways. Toxins 2016, 8, 214. [Google Scholar] [CrossRef]
- Echan, W.T.; Eespinosa, M.; Eyeo, C.C. Keeping the Wolves at Bay: Antitoxins of Prokaryotic Type II Toxin-Antitoxin Systems. Front. Mol. Biosci. 2016, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Kędzierska, B.; Hayes, F. Transcriptional Control of Toxin-Antitoxin Expression: Keeping Toxins Under Wraps Until the Time is Right. In Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; Wiley: Hoboken, NJ, USA, 2016; pp. 463–472. [Google Scholar]
- Mutschler, H.; Gebhardt, M.; Shoeman, R.L.; Meinhart, A. A Novel Mechanism of Programmed Cell Death in Bacteria by Toxin–Antitoxin Systems Corrupts Peptidoglycan Synthesis. PLoS Biol. 2011, 9, e1001033. [Google Scholar] [CrossRef]
- Maisonneuve, E.; Gerdes, K. Molecular Mechanisms Underlying Bacterial Persisters. Cell 2014, 157, 539–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Melderen, L. Toxin-antitoxin systems: Why so many, what for? Curr. Opin. Microbiol. 2010, 13, 781–785. [Google Scholar] [CrossRef]
- Wen, Y.; Behiels, E.; Devreese, B. Toxin-Antitoxin systems: Their role in persistence, biofilm formation, and pathogenicity. Pathog. Dis. 2014, 70, 240–249. [Google Scholar] [CrossRef]
- Goeders, N.; van Melderen, L. Toxin-Antitoxin Systems as Multilevel Interaction Systems. Toxins 2014, 6, 304–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, Targets, and Triggers: An Overview of Toxin-Antitoxin Biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef] [Green Version]
- Fraikin, N.; Goormaghtigh, F.; van Melderen, L. Type II Toxin-Antitoxin Systems: Evolution and Revolutions. J. Bacteriol. 2020, 202, 00719–00763. [Google Scholar] [CrossRef] [Green Version]
- Jurėnas, D.; Van Melderen, L. The Variety in the Common Theme of Translation Inhibition by Type II Toxin–Antitoxin Systems. Front. Genet. 2020, 11, 262. [Google Scholar] [CrossRef]
- Kędzierska, B.; Hayes, F. Emerging Roles of Toxin-Antitoxin Modules in Bacterial Pathogenesis. Molecules 2016, 21, 790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Inoue, K.; Yoshizumi, S.; Kobayashi, H.; Zhang, Y.; Ouyang, M.; Kato, F.; Sugai, M.; Inouye, M. Staphylococcus aureus MazF Specifically Cleaves a Pentad Sequence, UACAU, Which Is Unusually Abundant in the mRNA for Pathogenic Adhesive Factor SraP. J. Bacteriol. 2009, 191, 3248–3255. [Google Scholar] [CrossRef] [Green Version]
- Wen, W.; Liu, B.; Xue, L.; Zhu, Z.; Niu, L.; Sun, B. Autoregulation and Virulence Control by the Toxin-Antitoxin System SavRS inStaphylococcus aureus. Infect. Immun. 2018, 86, e00032-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Sun, C.; Li, Y.; Tang, K.; Ni, S.; Wang, X. Antitoxin HigA inhibits virulence gene mvfR expression in Pseudomonas aeruginosa. Environ. Microbiol. 2019, 21, 2707–2723. [Google Scholar] [CrossRef]
- Ni, S.; Li, B.; Tang, K.; Yao, J.; Wood, T.K.; Wang, P.; Wang, X. Conjugative plasmid-encoded toxin–antitoxin system PrpT/PrpA directly controls plasmid copy number. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Lorenzo-Díaz, F.; Fernández-López, C.; Lurz, R.; Bravo, A.; Espinosa, M. Crosstalk between vertical and horizontal gene transfer: Plasmid replication control by a conjugative relaxase. Nucleic Acids Res. 2017, 45, 7774–7785. [Google Scholar] [CrossRef] [PubMed]
- Christensen-Dalsgaard, M.; Jørgensen, M.G.; Gerdes, K. Three new RelE-homologous mRNA interferases of Escherichia coli differentially induced by environmental stresses. Mol. Microbiol. 2010, 75, 333–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neubauer, C.; Gao, Y.-G.; Andersen, K.R.; Dunham, C.M.; Kelley, A.C.; Hentschel, J.; Gerdes, K.; Ramakrishnan, V.; Brodersen, D.E. The Structural Basis for mRNA Recognition and Cleavage by the Ribosome-Dependent Endonuclease RelE. Cell 2009, 139, 1084–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, K.; Zavialov, A.V.; Pavlov, M.Y.; Elf, J.; Gerdes, K.; Ehrenberg, M. The Bacterial Toxin RelE Displays Codon-Specific Cleavage of mRNAs in the Ribosomal A Site. Cell 2003, 112, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Cataudella, I.; Trusina, A.; Sneppen, K.; Gerdes, K.; Mitarai, N. Conditional cooperativity in toxin–antitoxin regulation prevents random toxin activation and promotes fast translational recovery. Nucleic Acids Res. 2012, 40, 6424–6434. [Google Scholar] [CrossRef] [PubMed]
- Hoskins, J.; Alborn, W.E.; Arnold, J.; Blaszczak, L.C.; Burgett, S.; Dehoff, B.S.; Estrem, S.T.; Fritz, L.; Fu, D.-J.; Fuller, W.; et al. Genome of the Bacterium Streptococcus pneumoniae Strain R6. J. Bacteriol. 2001, 183, 5709–5717. [Google Scholar] [CrossRef] [Green Version]
- Khoo, S.K.; Loll, B.; Chan, W.T.; Shoeman, R.L.; Ngoo, L.; Yeo, C.C.; Meinhart, A. Molecular and Structural Characterization of the PezAT Chromosomal Toxin-Antitoxin System of the Human Pathogen Streptococcus pneumoniae. J. Biol. Chem. 2007, 282, 19606–19618. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.T.; Espinosa, M. The Streptococcus pneumoniae pezAT Toxin–Antitoxin System Reduces β-Lactam Resistance and Genetic Competence. Front. Microbiol. 2016, 7, 1322. [Google Scholar] [CrossRef] [Green Version]
- Nieto, C.; Pellicer, T.; Balsa, D.; Christensen, S.K.; Gerdes, K.; Espinosa, M. The chromosomal relBE2 toxin-antitoxin locus of Streptococcus pneumoniae: Characterization and use of a bioluminescence resonance energy transfer assay to detect toxin-antitoxin interaction. Mol. Microbiol. 2006, 59, 1280–1296. [Google Scholar] [CrossRef]
- Chan, W.T.; Nieto, C.; Harikrishna, J.A.; Khoo, S.K.; Othman, R.Y.; Espinosa, M.; Yeo, C.C. Genetic Regulation of the yefM-yoeB Toxin-Antitoxin Locus of Streptococcus pneumoniae. J. Bacteriol. 2011, 193, 4612–4625. [Google Scholar] [CrossRef] [Green Version]
- Nieto, C.; Cherny, I.; Khoo, S.K.; García de Lacoba, M.; Chan, W.T.; Yeo, C.C.; Gazit, E.; Espinosa, M. The yefM-yoeB toxin-antitoxin systems of Escherichia coli and Streptococcus pneumoniae: Functional and structural correlation. J. Bacteriol. 2007, 189, 1266–1278. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.T.; Yeo, C.C.; Sadowy, E.; Espinosa, M. Functional validation of putative toxin-antitoxin genes from the Gram-positive pathogen Streptococcus pneumoniae: Phd-doc is the fourth bona-fide operon. Front. Microbiol. 2014, 5, 677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerdes, K.; Maisonneuve, E. Bacterial Persistence and Toxin-Antitoxin Loci. Annu. Rev. Microbiol. 2012, 66, 103–123. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.K.; Gerdes, K. RelE toxins from Bacteria and Archaea cleave mRNAs on translating ribosomes, which are rescued by tmRNA. Mol. Microbiol. 2003, 48, 1389–1400. [Google Scholar] [CrossRef] [PubMed]
- Christensen-Dalsgaard, M.; Gerdes, K. Translation affects YoeB and MazF messenger RNA interferase activities by different mechanisms. Nucleic Acids Res. 2008, 36, 6472–6481. [Google Scholar] [CrossRef]
- Nieto, C.; Sadowy, E.; de la Campa, A.G.; Hryniewicz, W.; Espinosa, M. The relBE2Spn Toxin-Antitoxin System of Streptococcus pneumoniae: Role in Antibiotic Tolerance and Functional Conservation in Clinical Isolates. PLoS ONE 2010, 5, e11289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croucher, N.J.; Vernikos, G.S.; Parkhill, J.; Bentley, S.D. Identification, variation and transcription of pneumococcal repeat sequences. BMC Genom. 2011, 12, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, W.T.; Domenech, M.; Moreno-Córdoba, I.; Navarro-Martínez, V.; Nieto, C.; Moscoso, M.; García, E.; Espinosa, M. The Streptococcus pneumoniae yefM-yoeB and relBE Toxin-Antitoxin Operons Participate in Oxidative Stress and Biofilm Formation. Toxins 2018, 10, 378. [Google Scholar] [CrossRef] [Green Version]
- Aprianto, R.; Slager, J.; Holsappel, S.; Veening, J.-W. High-resolution analysis of the pneumococcal transcriptome under a wide range of infection-relevant conditions. Nucleic Acids Res. 2018, 46, 9990–10006. [Google Scholar] [CrossRef]
- LeRoux, M.; Culviner, P.H.; Liu, Y.J.; Littlehale, M.L.; Laub, M.T. Stress Can Induce Transcription of Toxin-Antitoxin Systems without Activating Toxin. Mol. Cell 2020, 79, 280–292. [Google Scholar] [CrossRef]
- Inniss, N.L.; Prehna, G.; Morrison, D.A. The pneumococcal σX activator, ComW, is a DNA-binding protein critical for natural transformation. J. Biol. Chem. 2019, 294, 11101–11118. [Google Scholar] [CrossRef]
- Studier, F.W.; Rosenberg, A.H.; Dunn, J.J.; Dubendorff, J.W. Use of T7 RNA polymerase to direct expression of cloned genes. Methods Enzymol. 1990, 185, 60–89. [Google Scholar] [CrossRef] [PubMed]
- Tettelin, H.; Nelson, K.E.; Paulsen, I.T.; Eisen, J.A.; Read, T.D.; Peterson, S.; Heidelberg, J.; de Boy, R.T.; Haft, D.H.; Dodson, R.J.; et al. Complete genome sequence of a virulent isolate of Streptococcus pneumoniae. Science 2001, 293, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Maniatis, T.; Fritsch, E.F.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1982. [Google Scholar]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Ruiz-Cruz, S.; Solano-Collado, V.; Espinosa, M.; Bravo, A. Novel plasmid-based genetic tools for the study of promoters and terminators in Streptococcus pneumoniae and Enterococcus faecalis. J. Microbiol. Methods 2010, 83, 156–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory, Cold Spring Harbor: New York, NY, USA, 1972; p. 466. [Google Scholar]
- Moreno-Córdoba, I.; Diago-Navarro, E.; Barendregt, A.; Heck, A.J.R.; Alfonso, C.; Díaz-Orejas, R.; Nieto, C.; Espinosa, M. The toxin-antitoxin proteins relBE2Spn of Streptococcus pneumoniae: Characterization and association to their DNA target. Proteins Struct. Funct. Bioinform. 2012, 80, 1834–1846. [Google Scholar] [CrossRef] [PubMed]
- Maxam, A.M.; Gilbert, W. Sequencing end-labeled DNA with base-specific chemical cleavages. Methods Enzymol. 1980, 65, 499–560. [Google Scholar] [CrossRef] [PubMed]
- Del Solar, G.; Pérez-Martín, J.; Espinosa, M. Plasmid pLS1-encoded RepA protein regulates transcription from repAB promoter by binding to a DNA sequence containing a 13-base pair symmetric element. J. Biol. Chem. 1990, 265, 12569–12575. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2--a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oggioni, M.R.; Claverys, J.P. Repeated extragenic sequences in prokaryotic genomes: A proposal for the origin and dynamics of the RUP element in Streptococcus pneumoniae. Microbiology 1999, 145, 2647–2653. [Google Scholar] [CrossRef] [Green Version]
- Slager, J.; Aprianto, R.; Veening, J.-W. Refining the Pneumococcal Competence Regulon by RNA Sequencing. J. Bacteriol. 2019, 201, e00780-18. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo-Diaz, F.; Moreno-Córdoba, I.; Espinosa, M. Complete labelling of pneumococcal DNA-binding proteins with seleno-L-methionine. J. Microbiol. Methods 2019, 166, 105720. [Google Scholar] [CrossRef] [PubMed]
- Kędzierska, B.; Lian, L.-Y.; Hayes, F. Toxin-antitoxin regulation: Bimodal interaction of YefM-YoeB with paired DNA palindromes exerts transcriptional autorepression. Nucleic Acids Res. 2006, 35, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.E.S.; Hayes, F. Influence of operator site geometry on transcriptional control by the YefM-YoeB toxin-antitoxin complex. J. Bacteriol. 2009, 191, 762–772. [Google Scholar] [CrossRef] [Green Version]
- Tullius, T.D.; Dombroski, B.A. Hydroxyl radical footprinting: High-resolution information about DNA-protein contacts and application to lambda repressor and Cro protein. Proc. Natl. Acad. Sci. USA 1986, 83, 5469–5473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, D.; Klug, A. Helical periodicity of DNA determined by enzyme digestion. Nat. Cell Biol. 1980, 286, 573–578. [Google Scholar] [CrossRef]
- Rhodes, D.; Klug, A. An underlying repeat in some transcriptional control sequences corresponding to half a double helical turn of DNA. Cell 1986, 46, 123–132. [Google Scholar] [CrossRef]
- Bøggild, A.; Sofos, N.; Andersen, K.R.; Feddersen, A.; Easter, A.D.; Passmore, L.A.; Brodersen, D.E. The Crystal Structure of the Intact E. coli RelBE Toxin-Antitoxin Complex Provides the Structural Basis for Conditional Cooperativity. Structure 2012, 20, 1641–1648. [Google Scholar] [CrossRef] [Green Version]
- Miallau, L.; Jain, P.; Jacobs, W.R.; Eisenberg, D.S.; Arbing, M.A.; Cascio, D.; Phan, T.; Ahn, C.J.; Chan, S.; Chernishof, I.; et al. Comparative Proteomics Identifies the Cell-Associated Lethality of M. tuberculosis RelBE-like Toxin-Antitoxin Complexes. Structure 2013, 21, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Overgaard, M.; Borch, J.; Jørgensen, M.G.; Gerdes, K. Messenger RNA interferase RelE controls relBE transcription by conditional cooperativity. Mol. Microbiol. 2008, 69, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Kędzierska, B.; Potrykus, K.; Szalewska-Pałasz, A.; Wodzikowska, B. Insights into transcriptional repression of the homologous Toxin-Antitoxin cassettes yefM-yoeB and axe-txe. Intl. J. Mol. Sci. 2020, 21, 9062. [Google Scholar] [CrossRef]
- Xue, L.; Yue, J.; Ke, J.; Khan, M.H.; Wen, W.; Sun, B.; Zhu, Z.; Niu, L. Distinct oligomeric structures of the YoeB–YefM complex provide insights into the conditional cooperativity of type II toxin–antitoxin system. Nucleic Acids Res. 2020, 48, 10527–10541. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Córdoba, I.; Chan, W.-T.; Nieto, C.; Espinosa, M. Interactions of the Streptococcus pneumoniae Toxin-Antitoxin RelBE Proteins with Their Target DNA. Microorganisms 2021, 9, 851. https://doi.org/10.3390/microorganisms9040851
Moreno-Córdoba I, Chan W-T, Nieto C, Espinosa M. Interactions of the Streptococcus pneumoniae Toxin-Antitoxin RelBE Proteins with Their Target DNA. Microorganisms. 2021; 9(4):851. https://doi.org/10.3390/microorganisms9040851
Chicago/Turabian StyleMoreno-Córdoba, Inmaculada, Wai-Ting Chan, Concha Nieto, and Manuel Espinosa. 2021. "Interactions of the Streptococcus pneumoniae Toxin-Antitoxin RelBE Proteins with Their Target DNA" Microorganisms 9, no. 4: 851. https://doi.org/10.3390/microorganisms9040851