
Encystation of Entamoeba histolytica in Axenic Culture

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. E. histolytica Culture Maintenance
2.3. Encystation and Sarkosyl Treatment
2.4. Viability and Excystation
2.5. Staining and Fluorescence Microscopy
2.6. Flow Cytometry Analysis
3. Results
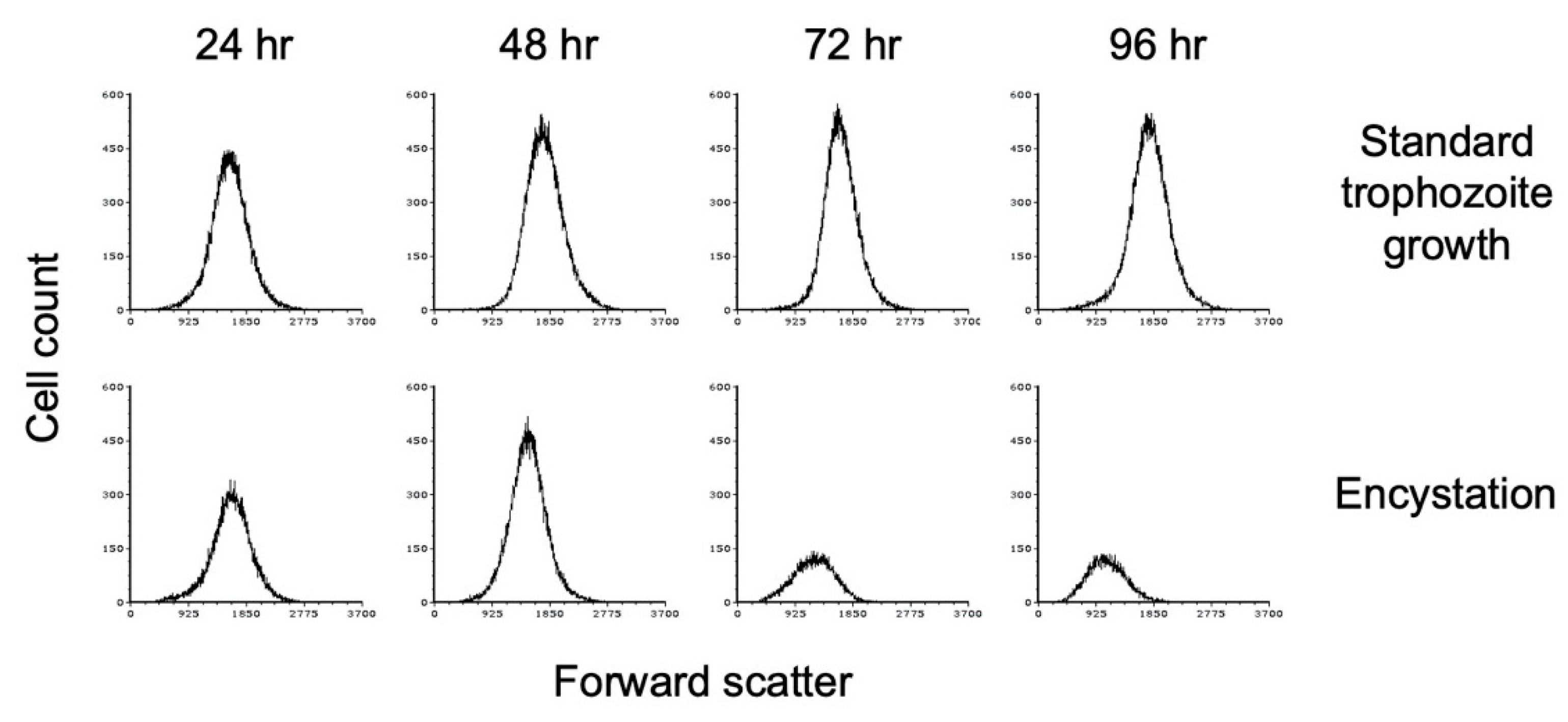
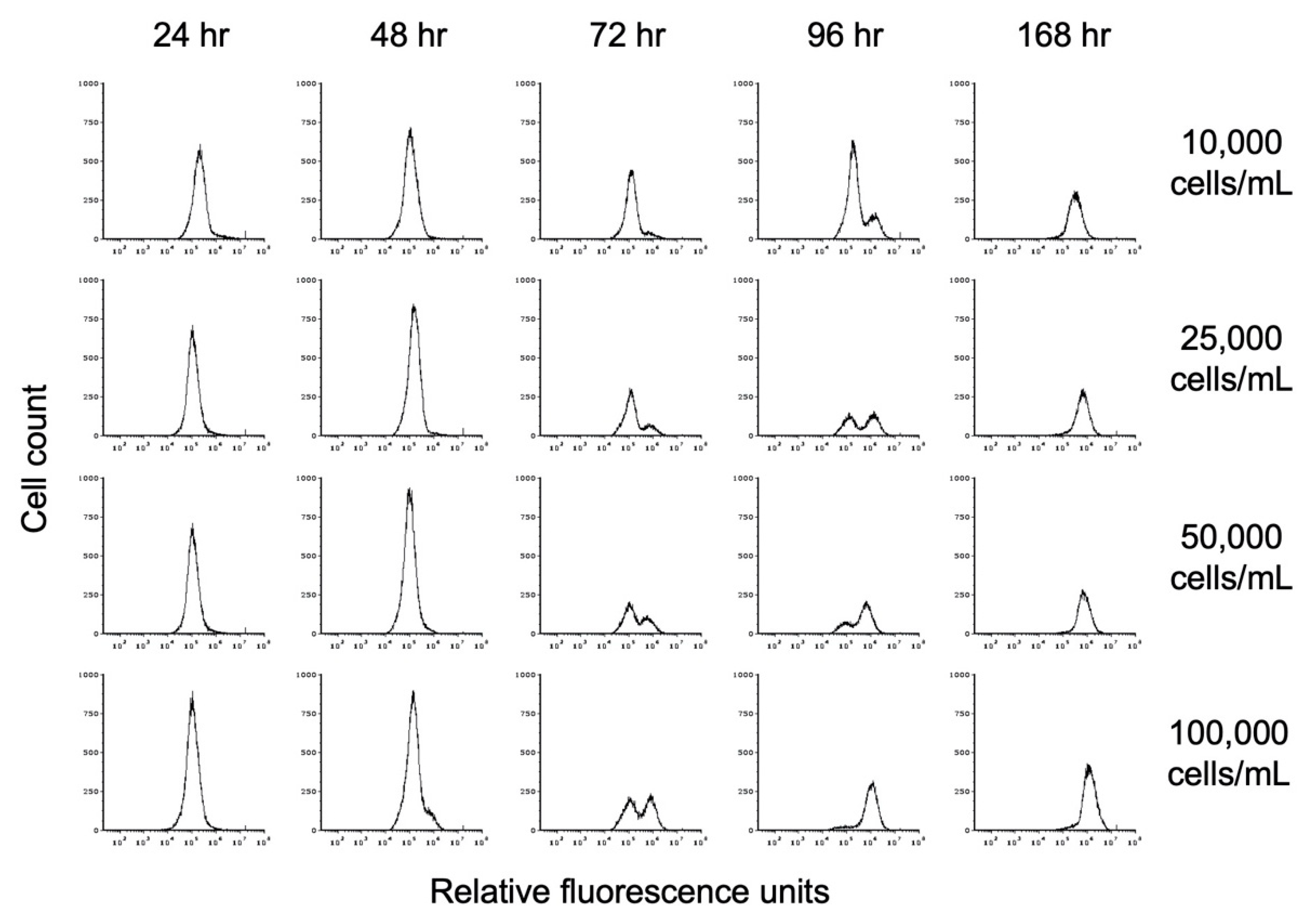
3.1. Growth Conditions for Encystation
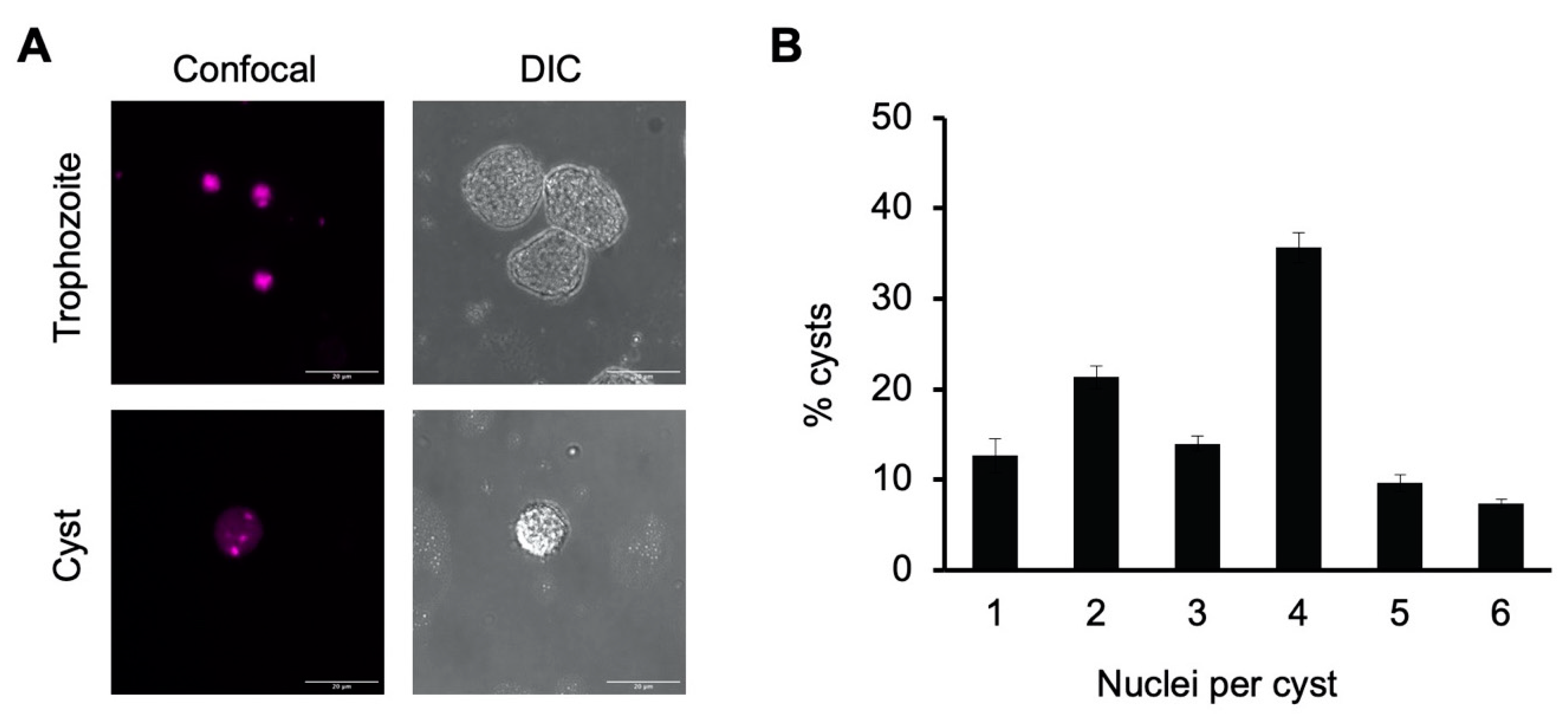
3.2. Tetranucleation
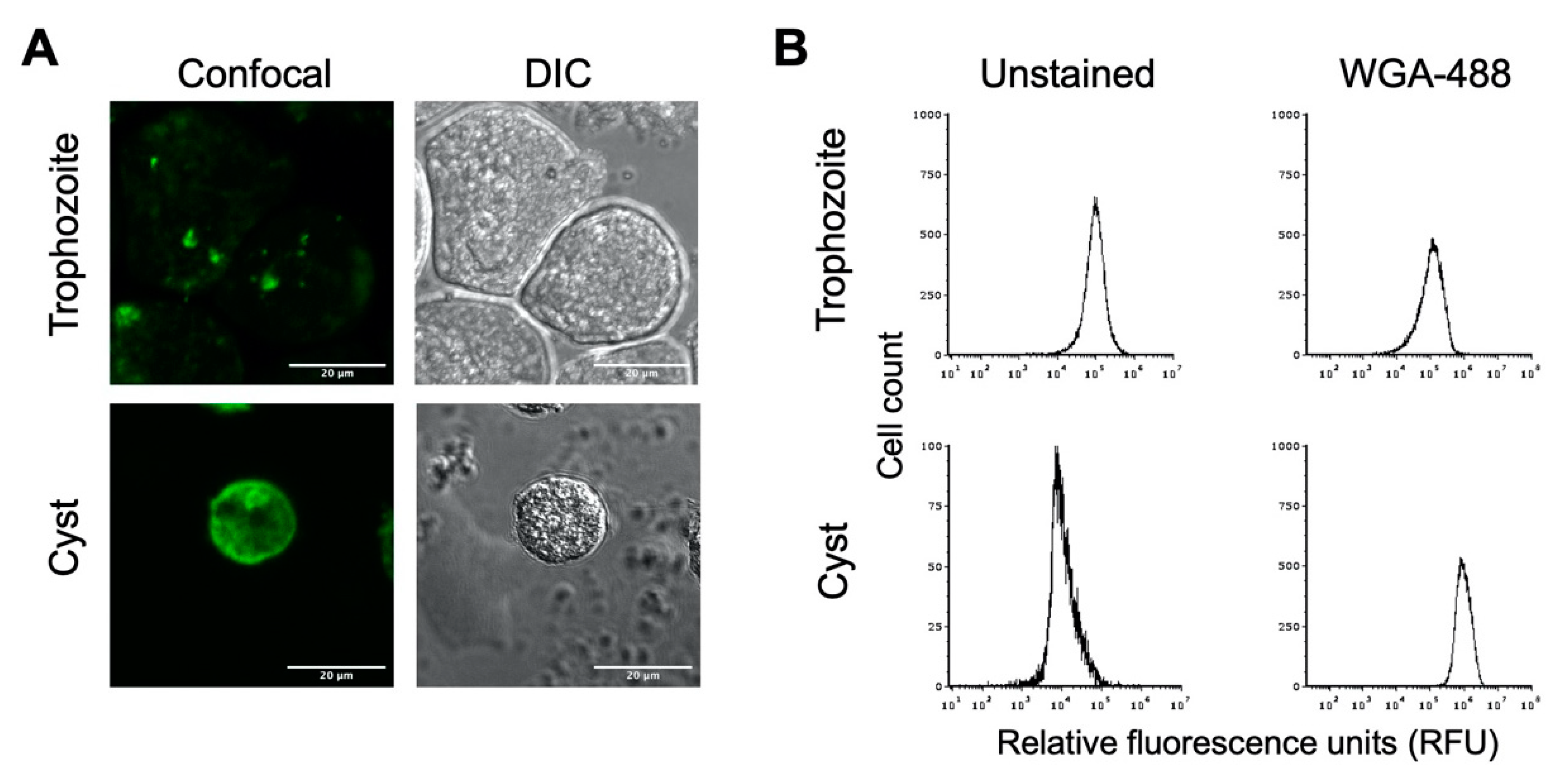
3.3. Chitin Cell Wall
3.4. Small Size and Roundedness
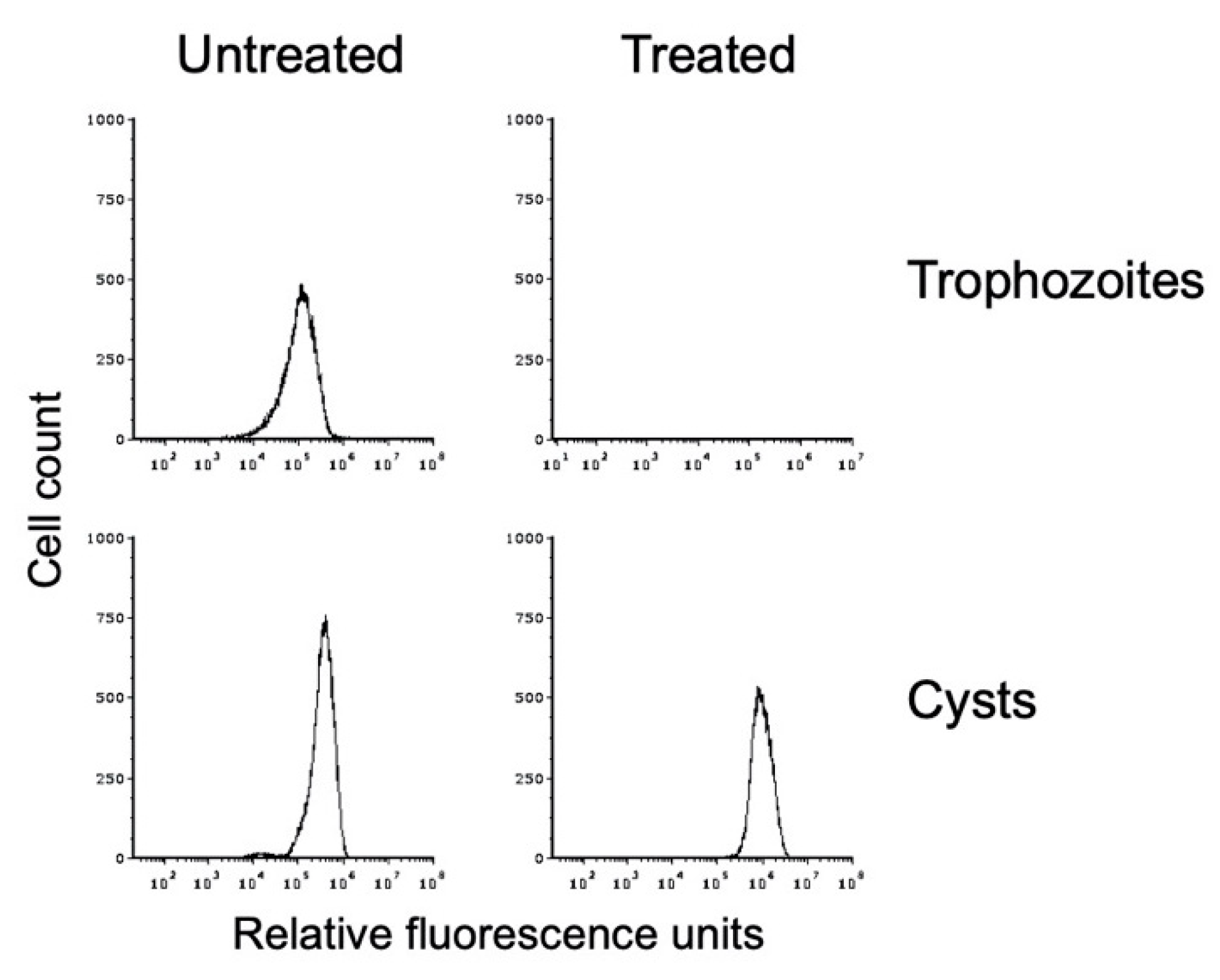
3.5. Detergent Resistance
3.6. Viability and Excystation
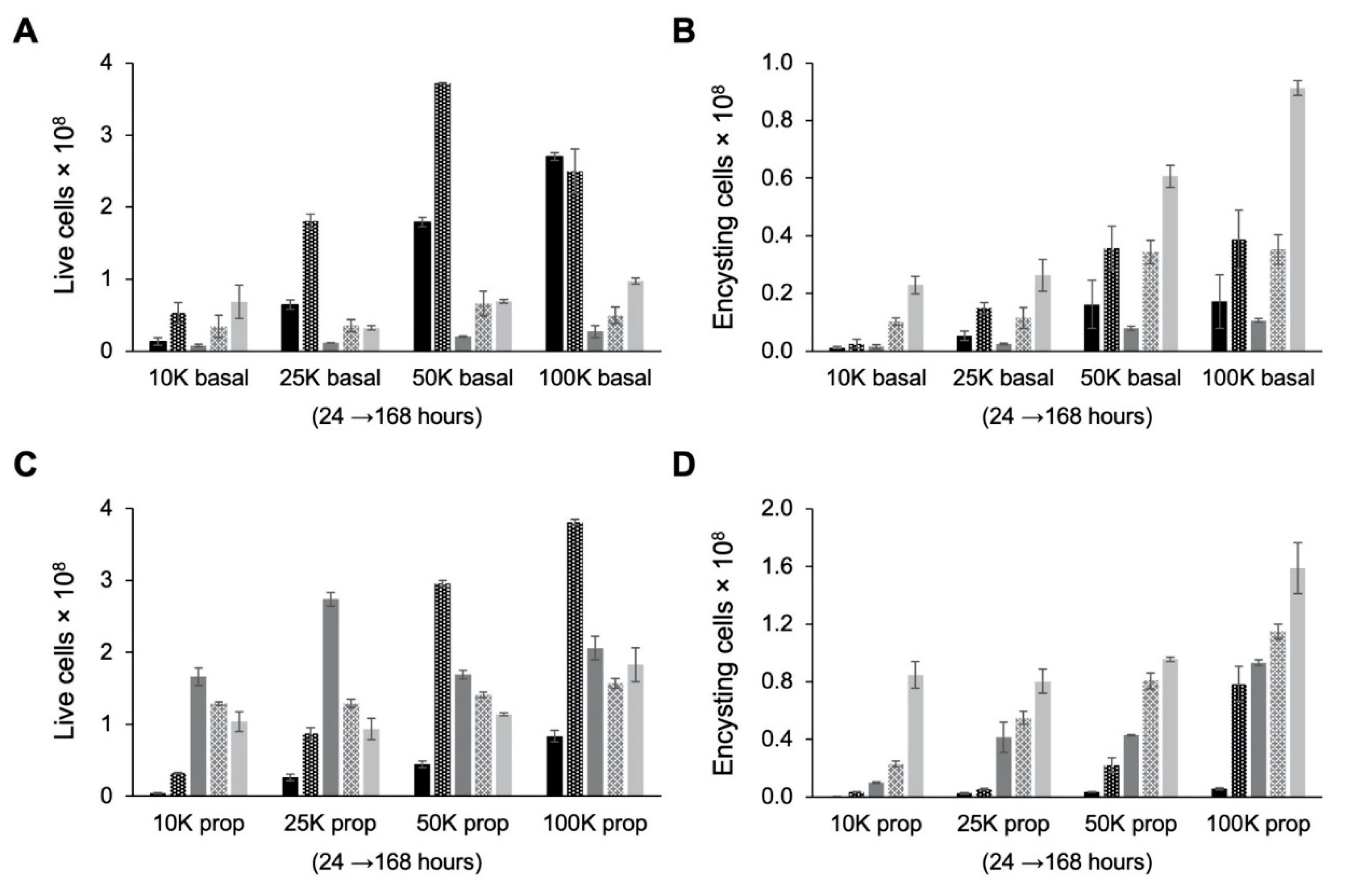
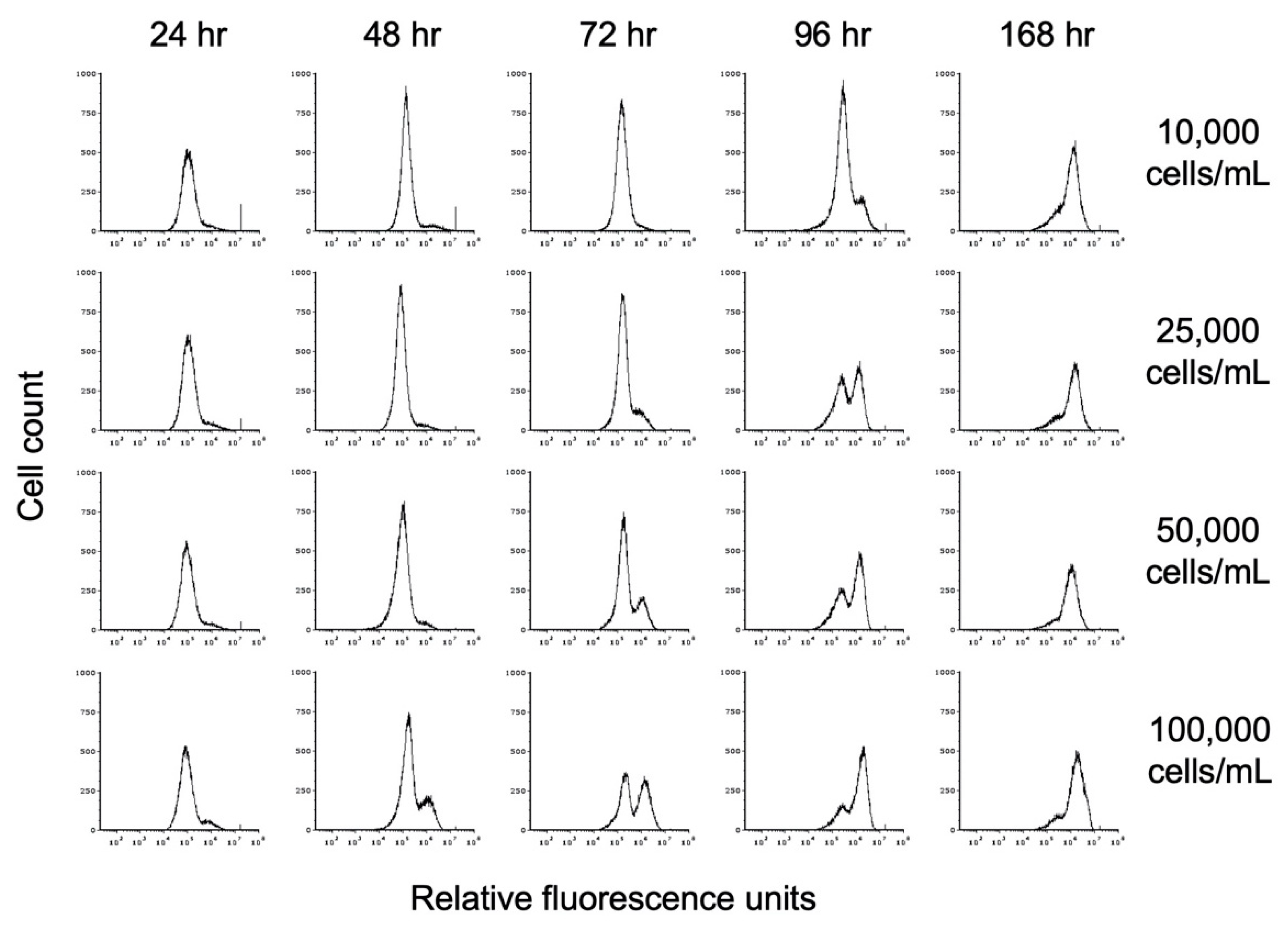
3.7. Role of Cell Density in Encystation
3.8. Effect of Propionate on Encystation
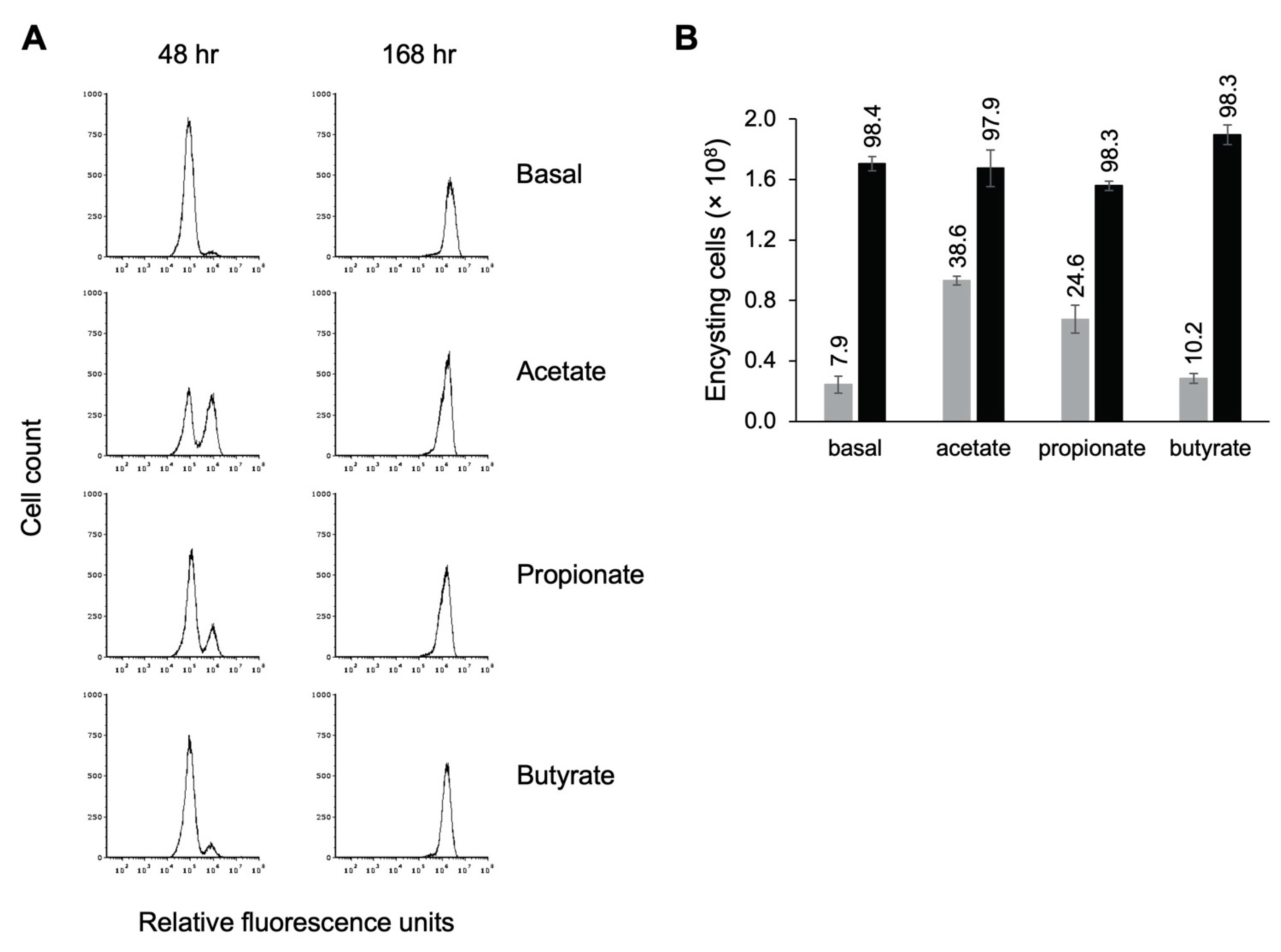
3.9. Effect of Other SCFAs on Encystation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ximenez, C.; Moran, P.; Rojas, L.; Valadez, A.; Gomez, A. Reassessment of the epidemiology of amoebiasis: State of the art. Infect. Genet. Evol. 2009, 9, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiology Reference Group 2007–2015. Available online: https://apps.who.int/iris/bitstream/handle/10665/199350/9789241565165_eng.pdf (accessed on 16 April 2021).
- Entamoeba taxonomy. Bull. World Health Organ. 1997, 75, 291–294.
- Verkerke, H.P.; Petri, W.A.; Marie, C.S. The dynamic interdependence of amebiasis, innate immunity, and undernutrition. Semin. Immunopathol. 2012, 34, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Baxt, L.A.; Singh, U. New insights into Entamoeba histolytica pathogenesis. Curr. Opin. Infect. Dis. 2008, 21, 489–494. [Google Scholar] [CrossRef]
- Schaap, P.; Schilde, C. Encystation: The most prevalent and under investigated differentiation pathway of eukaryotes. Microbiology 2018, 164, 727–739. [Google Scholar] [CrossRef]
- World Health Organization. Amoebiasis. Wkly. Epidemiol. Rec. 1997, 72, 97–100. [Google Scholar]
- WHO; PAHO; UNESCO. Report: A consultation with experts on amoebiasis. Epidemiol. Bull. Paho 1997, 18, 13–14. [Google Scholar]
- Beaver, P.C.; Jung, R.C.; Read, T.R.; Robinson, T.A.; Sherman, H.J. Experimental Entamoeba histolytica infections in man. Am. J. Trop. Med. Hyg. 1956, 5, 1000–1009. [Google Scholar] [CrossRef]
- Stanley, S.L., Jr. Amoebiasis. Lancet 2003, 361, 1025–1034. [Google Scholar] [CrossRef]
- Haque, R.; Huston, C.D.; Hughes, M.; Houpt, E.; Petri, W.A., Jr. Amebiasis. N. Engl. J. Med. 2003, 348, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Segovia-Gamboa, N.C.; Talamas-Rohana, P.; Angel-Martinez, A.; Cazares-Raga, F.E.; Gonzalez-Robles, A.; Hernandez-Ramirez, V.I.; Martinez-Palomo, A.; Chavez-Munguia, B. Differentiation of Entamoeba histolytica: A possible role for enolase. Exp. Parasitol. 2011, 129, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Diaz, H.; Diaz-Gallardo, M.; Laclette, J.P.; Carrero, J.C. In vitro induction of Entamoeba histolytica cyst-like structures from trophozoites. PLoS Negl. Trop. Dis. 2010, 4, e607. [Google Scholar] [CrossRef] [PubMed]
- Barron-Gonzalez, M.P.; Villarreal-Trevino, L.; Resendez-Perez, D.; Mata-Cardenas, B.D.; Morales-Vallarta, M.R. Entamoeba histolytica: Cyst-like structures in vitro induction. Exp. Parasitol. 2008, 118, 600–603. [Google Scholar] [CrossRef] [PubMed]
- Eichinger, D. Encystation in parasitic protozoa. Curr. Opin. Microbiol. 2001, 4, 421–426. [Google Scholar] [CrossRef]
- Singh, U.; Ehrenkaufer, G.M. Recent insights into Entamoeba development: Identification of transcriptional networks associated with stage conversion. Int. J. Parasitol. 2009, 39, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Chadee, K.; Meerovitch, E. Entamoeba histolytica: Early progressive pathology in the cecum of the gerbil (Meriones unguiculatus). Am. J. Trop. Med. Hyg. 1985, 34, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Ghadirian, E.; Kongshavn, P.A. Genetic control of susceptibility of mice to infection with E. histolytica. Parasite Immunol. 1984, 6, 349–360. [Google Scholar] [CrossRef]
- Ghosh, P.K.; Gupta, S.; Ortiz-Ortiz, L. Intestinal amoebiasis: Delayed-type hypersensitivity response in mice. J. Health Popul. Nutr. 2000, 18, 109–114. [Google Scholar]
- Ghosh, P.K.; Mancilla, R.; Ortiz-Ortiz, L. Intestinal amebiasis: Histopathologic features in experimentally infected mice. Arch. Med. Res. 1994, 25, 297–302. [Google Scholar]
- Houpt, E.R.; Glembocki, D.J.; Obrig, T.G.; Moskaluk, C.A.; Lockhart, L.A.; Wright, R.L.; Seaner, R.M.; Keepers, T.R.; Wilkins, T.D.; Petri, W.A., Jr. The mouse model of amebic colitis reveals mouse strain susceptibility to infection and exacerbation of disease by CD4+ T cells. J. Immunol. 2002, 169, 4496–4503. [Google Scholar] [CrossRef]
- Rivero-Nava, L.; Aguirre-Garcia, J.; Calderon, J. Production of amebic intestinal lesions in BALB/c mice. Arch. Med. Res. 1997, 28, 220–222. [Google Scholar] [PubMed]
- Said-Fernandez, S.; Mata-Cardenas, B.D.; Gonzalez-Garza, M.T.; Navarro-Marmolejo, L.; Rodriguez-Perez, E. Entamoeba histolytica cysts with a defective wall formed under axenic conditions. Parasitol. Res. 1993, 79, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Rengpien, S.; Bailey, G.B. Differentiation of Entamoeba: A new medium and optimal conditions for axenic encystation of E. invadens. J. Parasitol. 1975, 61, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Thepsuparungsikul, V.; Seng, L.; Bailey, G.B. Differentiation of Entamoeba: Encystation of E. invadens in monoxenic and axenic cultures. J. Parasitol. 1971, 57, 1288–1292. [Google Scholar] [CrossRef] [PubMed]
- Avron, B.; Stolarsky, T.; Chayen, A.; Mirelman, D. Encystation of Entamoeba invadens IP-1 is induced by lowering the osmotic pressure and depletion of nutrients from the medium. J. Protozool. 1986, 33, 522–525. [Google Scholar] [CrossRef] [PubMed]
- Vazquezdelara-Cisneros, L.G.; Arroyo-Begovich, A. Induction of encystation of Entamoeba invadens by removal of glucose from the culture medium. J. Parasitol. 1984, 70, 629–633. [Google Scholar] [CrossRef]
- Eichinger, D. Encystation of Entamoeba parasites. Bioessays 1997, 19, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Richards, C.S.; Goldman, M.; Cannon, L.T. Cultivation of Entamoeba histolytica and Entamoeba histolytica-like strains at reduced temperature and behavior of the amebae in diluted media. Am. J. Trop. Med. Hyg. 1966, 15, 648–655. [Google Scholar] [CrossRef]
- Ehrenkaufer, G.M.; Weedall, G.D.; Williams, D.; Lorenzi, H.A.; Caler, E.; Hall, N.; Singh, U. The genome and transcriptome of the enteric parasite Entamoeba invadens, a model for encystation. Genome Biol. 2013, 14, R77. [Google Scholar] [CrossRef]
- Ali, I.K.; Haque, R.; Siddique, A.; Kabir, M.; Sherman, N.E.; Gray, S.A.; Cangelosi, G.A.; Petri, W.A., Jr. Proteomic analysis of the cyst stage of Entamoeba histolytica. PLoS Negl. Trop. Dis. 2012, 6, e1643. [Google Scholar] [CrossRef]
- Diamond, L.S. Techniques of axenic cultivation of Entamoeba histolytica Schaudinn, 1903 and E. histolytica-like amebae. J. Parasitol. 1968, 54, 1047–1056. [Google Scholar] [CrossRef]
- Schupp, D.G.; Januschka, M.M.; Sherlock, L.A.; Stibbs, H.H.; Meyer, E.A.; Bemrick, W.J.; Erlandsen, S.L. Production of viable Giardia cysts in vitro: Determination by fluorogenic dye staining, excystation, and animal infectivity in the mouse and Mongolian gerbil. Gastroenterology 1988, 95, 1–10. [Google Scholar] [CrossRef]
- Mirelman, D.; Avron, B. Cyst formation in Entamoeba. In Amebiasis: Human Infection by Entamoeba Histolytica; Ravdin, J.I., Ed.; Wiley Medical Publications: New York, NY, USA, 1988; pp. 768–781. [Google Scholar]
- Lopez-Romero, E.; Villagomez-Castro, J.C. Encystation in Entamoeba invadens. Parasitol. Today 1993, 9, 225–227. [Google Scholar] [CrossRef]
- Tanyuksel, M.; Petri, W.A. Laboratory diagnosis of amoebiasis. Clin. Microbiol. Rev. 2003, 16, 713–729. [Google Scholar] [CrossRef]
- Clark, C.G.; Espinosa-Cantellano, M.; Bhattacharya, A. Entamoeba histolytica: An overview of the biology of the organism. In Amebiasis; Radvin, J.I., Ed.; Series on Tropical Medicine; Imperial College Press: London, UK, 2000; pp. 1–45. [Google Scholar]
- Mukherjee, C.; Clark, C.G.; Lohia, A. Entamoeba shows reversible variation in ploidy under different growth conditions and between life cycle phases. PLoS Negl. Trop. Dis. 2008, 2, e281. [Google Scholar] [CrossRef]
- Niculescu, V.F. Developmental and non developmental polyploidy in xenic and axenic cultured stem cell lines of Entamoeba invadens and Entamoeba histolytica. Insights Stem Cells 2016, 2, 1–10. [Google Scholar]
- Ganguly, A.; Lohia, A. The cell cycle of Entamoeba invadens during vegetative growth and differentiation. Mol. Biochem. Parasitol. 2001, 112, 277–285. [Google Scholar] [CrossRef]
- Luna-Nacar, M.; Navarrete-Perea, J.; Moguel, B.; Bobes, R.J.; Laclette, J.P.; Carrero, J.C. Proteomic study of Entamoeba histolytica trophozoites, cysts, and cyst-like structures. PLoS ONE 2016, 11, e0156018. [Google Scholar] [CrossRef]
- Garcia-Zapien, A.G.; Hernandez-Gutierrez, R.; Mora-Galindo, J. Simultaneous growth and mass encystation of Entamoeba invadens under axenic conditions. Arch. Med. Res. 1995, 26, 257–262. [Google Scholar]
- Campos-Gongora, E.; Ebert, F.; Willhoeft, U.; Said-Fernandez, S.; Tannich, E. Characterization of chitin synthases from Entamoeba. Protist 2004, 155, 323–330. [Google Scholar] [CrossRef]
- Chatterjee, A.; Ghosh, S.K.; Jang, K.; Bullitt, E.; Moore, L.; Robbins, P.W.; Samuelson, J. Evidence for a “wattle and daub” model of the cyst wall of Entamoeba. PLoS Pathog. 2009, 5, e1000498. [Google Scholar] [CrossRef]
- Makioka, A.; Kumagai, M.; Hiranuka, K.; Kobayashi, S.; Takeuchi, T. Different structure and mRNA expression of Entamoeba invadens chitinases in the encystation and excystation. Parasitol. Res. 2011, 109, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Mi-Ichi, F.; Miyake, Y.; Tam, V.K.; Yoshida, H. A flow cytometry method for dissecting the cell differentiation process of Entamoeba encystation. Front. Cell Infect. Microbiol 2018, 8, 250. [Google Scholar] [CrossRef] [PubMed]
- Welter, B.H.; Sehorn, M.G.; Temesvari, L.A. Flow cytometric characterization of encystation in Entamoeba invadens. Mol. Biochem. Parasitol. 2017, 218, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Begovich, A.; Carabez-Trejo, A. Location on chitin in the cyst wall of Entamoeba invadens with colloidal gold tracers. J. Parasitol. 1982, 68, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Begovich, A.; Carabez-Trejo, A.; Ruiz-Herrera, J. Identification of the structural component in the cyst wall of Entamoeba invadens. J. Parasitol. 1980, 66, 735–741. [Google Scholar] [CrossRef]
- Bailey, G.B.; Rengypian, S. Osmotic stress as a factor controlling encystation of Entamoeba invadens. Arch. Investig. Med. 1980, 11, 11–16. [Google Scholar]
- Mi-Ichi, F.; Yoshida, H.; Hamano, S. Entamoeba encystation: New targets to prevent the transmission of amebiasis. PLoS Pathog. 2016, 12, e1005845. [Google Scholar] [CrossRef]
- Balamuth, W. Effects of some environmental factors upon growth and encystation of Entamoeba invadens. J. Parasitol. 1962, 48, 101–109. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef]
- Cummings, J.H.; MacFarlane, G.T. Role of intestinal bacteria in nutrient metabolism. Clin. Nutr. 1997, 16, 3–11. [Google Scholar] [CrossRef]
- Roediger, W.E. Role of anaerobic bacteria in the metabolic welfare of the colonic mucosa in man. Gut 1980, 21, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Byers, J.; Faigle, W.; Eichinger, D. Colonic short-chain fatty acids inhibit encystation of Entamoeba invadens. Cell Microbiol. 2005, 7, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Barboza, P.S. Digesta passage and functional anatomy of the digestive tract in the desert tortoise (Xerobates agassizii). J. Comp. Physiol 1995, 165, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Bjorndal, K.A.; Bolten, A.B. Digestive processing in a herbivorous freshwater turtle: Consequences of small intestine fermentation. Physiol. Zool. 1990, 63, 1232–1247. [Google Scholar] [CrossRef]
- Bjorndal, K.A. Cellulose digestion and volatile fatty acid production in the green turtle. Chelonia mydas. Comp. Biochem. Physiol. 1979, 63A, 127–133. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Starting Cell Density (cells/mL) | TYI Basal | TYI Propionate | p-Value: Basal vs. Propionate |
|---|---|---|---|
| Total cysts produced (× 107) | |||
| 10K | 2.29 ± 0.30 | 8.48 ± 0.93 | p = 0.0004 |
| 25K | 2.63 ± 0.55 | 8.04 ± 0.84 | p = 0.0007 |
| 50K | 6.07 ± 0.39 | 9.57 ± 0.16 | p = 0.0001 |
| 100K | 9.13 ± 0.25 | 15.90 ± 1.77 | p = 0.0028 |
| Encystation efficiency (% cysts/live cells) | |||
| 10K | 33.5 ± 4.3 | 81.5 ± 8.9 | p = 0.0011 |
| 25K | 81.7 ± 17.0 | 86.1 ± 9.0 | NS |
| 50K | 87.6 ± 5.6 | 83.9 ± 1.4 | NS |
| 100K | 93.8 ± 2.6 | 86.9 ± 9.7 | NS |
| Encystation rate (total cysts/total starting cell population) | |||
| 10K | 44.0 ± 5.7 | 163.0 ± 17.8 | p = 0.0004 |
| 25K | 20.2 ± 4.2 | 61.8 ± 6.5 | p = 0.0007 |
| 50K | 23.3 ± 1.5 | 36.8 ± 0.6 | p = 0.0001 |
| 100K | 17.6 ± 0.5 | 30.6 ± 3.4 | p = 0.0028 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wesel, J.; Shuman, J.; Bastuzel, I.; Dickerson, J.; Ingram-Smith, C. Encystation of Entamoeba histolytica in Axenic Culture. Microorganisms 2021, 9, 873. https://doi.org/10.3390/microorganisms9040873
Wesel J, Shuman J, Bastuzel I, Dickerson J, Ingram-Smith C. Encystation of Entamoeba histolytica in Axenic Culture. Microorganisms. 2021; 9(4):873. https://doi.org/10.3390/microorganisms9040873
Chicago/Turabian StyleWesel, Jordan, Jennifer Shuman, Irem Bastuzel, Julie Dickerson, and Cheryl Ingram-Smith. 2021. "Encystation of Entamoeba histolytica in Axenic Culture" Microorganisms 9, no. 4: 873. https://doi.org/10.3390/microorganisms9040873
APA StyleWesel, J., Shuman, J., Bastuzel, I., Dickerson, J., & Ingram-Smith, C. (2021). Encystation of Entamoeba histolytica in Axenic Culture. Microorganisms, 9(4), 873. https://doi.org/10.3390/microorganisms9040873