
Human Cytomegalovirus Reduces Endothelin-1 Expression in Both Endothelial and Vascular Smooth Muscle Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Stock and Cells
2.2. Virus Infection
2.3. RNA Isolation and TaqMan Real-Time PCR Assay
2.4. Human Endothelin-1 QuantiGlo ELISA Kit
2.5. Immunofluorescent Staining (IF)
2.6. Western Blot Analysis
2.7. Short Interfering RNA (siRNA) against IE72 and IE86
2.8. Statistical Analysis
3. Results
3.1. HCMV Infection Downregulates ET-1 Transcript Production and Release of ET-1 Peptide from Infected Endothelial and Smooth Muscle Cells
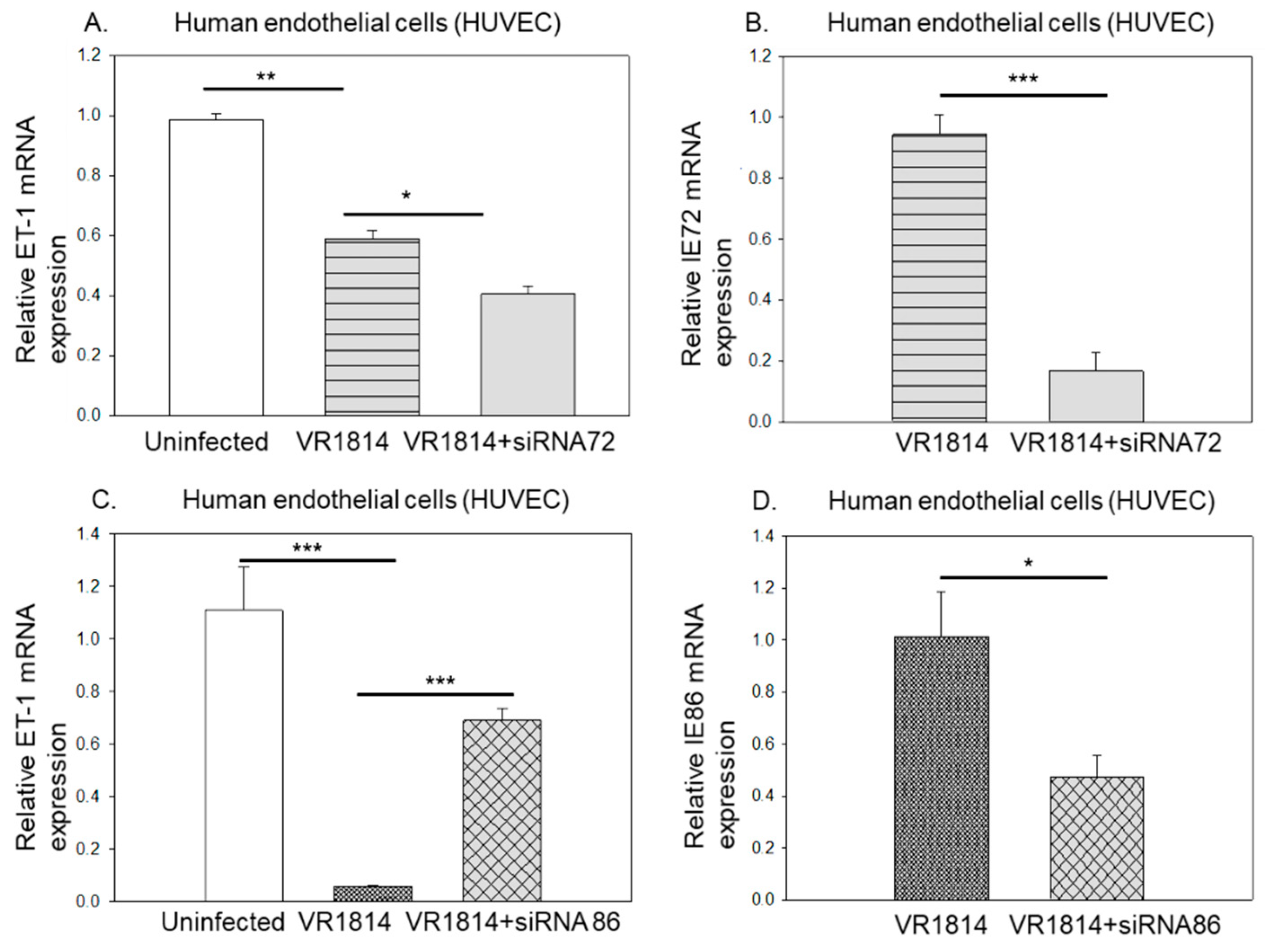
3.2. HCMV IE86 Downregulates ET-1 Transcript Levels
3.3. Ganciclovir Does Not Prevent HCMV’s Inhibitory Effect on ET-1 Production, Which Is Mainly Regulated by HCMV IE2-p86
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.; Yu, D.; Grimwood, J.; Schmutz, J.; Dickson, M.; Jarvis, M.A.; Hahn, G.; Nelson, J.A.; Myers, R.M.; Shenk, T.E. Coding potential of laboratory and clinical strains of human cytomegalovirus. Proc. Natl. Acad. Sci. USA 2003, 100, 14976–14981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern-Ginossar, N.; Weisburd, B.; Michalski, A.; Le, V.T.; Hein, M.Y.; Huang, S.X.; Ma, M.; Shen, B.; Qian, S.B.; Hengel, H.; et al. Decoding human cytomegalovirus. Science 2012, 338, 1088–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, W.; Chou, C.; Li, H.; Hai, R.; Patterson, D.; Stolc, V.; Zhu, H.; Liu, F. Functional profiling of a human cytomegalovirus genome. Proc. Natl. Acad. Sci. USA 2003, 100, 14223–14228. [Google Scholar] [CrossRef] [Green Version]
- Soderberg-Naucler, C. Does cytomegalovirus play a causative role in the development of various inflammatory diseases and cancer? J. Intern. Med. 2006, 259, 219–246. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.S.; Venter, P.C.; de Wet, J.I. Cytomegalovirus and atherosclerosis. S. Afr. Med. J. 1984, 65, 793. [Google Scholar]
- Adam, E.; Melnick, J.L.; Probtsfield, J.L.; Petrie, B.L.; Burek, J.; Bailey, K.R.; McCollum, C.H.; DeBakey, M.E. High levels of cytomegalovirus antibody in patients requiring vascular surgery for atherosclerosis. Lancet 1987, 2, 291–293. [Google Scholar] [CrossRef]
- Grattan, M.T.; Moreno-Cabral, C.E.; Starnes, V.A.; Oyer, P.E.; Stinson, E.B.; Shumway, N.E. Cytomegalovirus infection is associated with cardiac allograft rejection and atherosclerosis. JAMA 1989, 261, 3561–3566. [Google Scholar] [CrossRef] [PubMed]
- Baryawno, N.; Rahbar, A.; Wolmer-Solberg, N.; Taher, C.; Odeberg, J.; Darabi, A.; Khan, Z.; Sveinbjornsson, B.; FuskevAg, O.M.; Segerstrom, L.; et al. Detection of human cytomegalovirus in medulloblastomas reveals a potential therapeutic target. J. Clin. Investig. 2011, 121, 4043–4055. [Google Scholar] [CrossRef] [Green Version]
- Popovic, M.; Smiljanic, K.; Dobutovic, B.; Syrovets, T.; Simmet, T.; Isenovic, E.R. Human cytomegalovirus infection and atherothrombosis. J. Thromb. Thrombolysis 2012, 33, 160–172. [Google Scholar] [CrossRef]
- Abgueguen, P.; Delbos, V.; Chennebault, J.M.; Payan, C.; Pichard, E. Vascular thrombosis and acute cytomegalovirus infection in immunocompetent patients: Report of 2 cases and literature review. Clin. Infect. Dis. 2003, 36, E134–E139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.F.; Leon, M.B.; Waclawiw, M.A.; Popma, J.J.; Yu, Z.X.; Finkel, T.; Epstein, S.E. Association between prior cytomegalovirus infection and the risk of restenosis after coronary atherectomy. N. Engl. J. Med. 1996, 335, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Gredmark-Russ, S.; Dzabic, M.; Rahbar, A.; Wanhainen, A.; Bjorck, M.; Larsson, E.; Michel, J.B.; Soderberg-Naucler, C. Active cytomegalovirus infection in aortic smooth muscle cells from patients with abdominal aortic aneurysm. J. Mol. Med. 2009, 87, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Roberts, E.T.; Haan, M.N.; Dowd, J.B.; Aiello, A.E. Cytomegalovirus antibody levels, inflammation, and mortality among elderly Latinos over 9 years of follow-up. Am. J. Epidemiol. 2010, 172, 363–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strandberg, T.E.; Pitkala, K.H.; Tilvis, R.S. Cytomegalovirus antibody level and mortality among community-dwelling older adults with stable cardiovascular disease. JAMA 2009, 301, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.W.; Cech, I. Association of type 2 diabetes mellitus and seroprevalence for cytomegalovirus. South. Med. J. 2005, 98, 686–692. [Google Scholar] [CrossRef]
- Hui, J.; Qu, Y.Y.; Tang, N.; Liu, Y.M.; Zhong, H.; Wang, L.M.; Feng, Q.; Li, Z.; He, F. Association of cytomegalovirus infection with hypertension risk: A meta-analysis. Wien. Klin. Wochenschr. 2016, 128, 586–591. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.C.; Hruban, R.H.; Ambinder, R.F.; Pizzorno, M.; Cameron, D.E.; Baumgartner, W.A.; Reitz, B.A.; Hayward, G.S.; Hutchins, G.M. Demonstration of cytomegalovirus nucleic acids in the coronary arteries of transplanted hearts. Am. J. Pathol. 1992, 140, 739–747. [Google Scholar]
- Yonemitsu, Y.; Nakagawa, K.; Tanaka, S.; Mori, R.; Sugimachi, K.; Sueishi, K. In situ detection of frequent and active infection of human cytomegalovirus in inflammatory abdominal aortic aneurysms: Possible pathogenic role in sustained chronic inflammatory reaction. Lab. Investig. 1996, 74, 723–736. [Google Scholar]
- Horvath, R.; Cerny, J.; Benedik, J., Jr.; Hokl, J.; Jelinkova, I.; Benedik, J. The possible role of human cytomegalovirus (HCMV) in the origin of atherosclerosis. J. Clin. Virol. 2000, 16, 17–24. [Google Scholar] [CrossRef]
- Speir, E.; Modali, R.; Huang, E.S.; Leon, M.B.; Shawl, F.; Finkel, T.; Epstein, S.E. Potential role of human cytomegalovirus and p53 interaction in coronary restenosis. Science 1994, 265, 391–394. [Google Scholar] [CrossRef]
- Cobbs, C.S.; Harkins, L.; Samanta, M.; Gillespie, G.Y.; Bharara, S.; King, P.H.; Nabors, L.B.; Cobbs, C.G.; Britt, W.J. Human cytomegalovirus infection and expression in human malignant glioma. Cancer Res. 2002, 62, 3347–3350. [Google Scholar] [PubMed]
- Harkins, L.; Volk, A.L.; Samanta, M.; Mikolaenko, I.; Britt, W.J.; Bland, K.I.; Cobbs, C.S. Specific localisation of human cytomegalovirus nucleic acids and proteins in human colorectal cancer. Lancet 2002, 360, 1557–1563. [Google Scholar] [CrossRef]
- Samanta, M.; Harkins, L.; Klemm, K.; Britt, W.J.; Cobbs, C.S. High prevalence of human cytomegalovirus in prostatic intraepithelial neoplasia and prostatic carcinoma. J. Urol. 2003, 170, 998–1002. [Google Scholar] [CrossRef] [PubMed]
- Wolmer-Solberg, N.; Baryawno, N.; Rahbar, A.; Fuchs, D.; Odeberg, J.; Taher, C.; Wilhelmi, V.; Milosevic, J.; Mohammad, A.A.; Martinsson, T.; et al. Frequent detection of human cytomegalovirus in neuroblastoma: A novel therapeutic target? Int. J. Cancer 2013, 133, 2351–2361. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Chen, Y.Y.; Chen, W.G.; Diamond, D.J.; Mamelak, A.N.; Zaia, J.A.; Weiss, L.M. Lack of association of cytomegalovirus with human brain tumors. Mod. Pathol. 2005, 18, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Hart, H.; Neill, W.A.; Norval, M. Lack of association of cytomegalovirus with adenocarcinoma of the colon. Gut 1982, 23, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridker, P.M.; Hennekens, C.H.; Stampfer, M.J.; Wang, F. Prospective study of herpes simplex virus, cytomegalovirus, and the risk of future myocardial infarction and stroke. Circulation 1998, 98, 2796–2799. [Google Scholar] [CrossRef] [Green Version]
- Sorlie, P.D.; Nieto, F.J.; Adam, E.; Folsom, A.R.; Shahar, E.; Massing, M. A prospective study of cytomegalovirus, herpes simplex virus 1, and coronary heart disease: The atherosclerosis risk in communities (ARIC) study. Arch. Intern. Med. 2000, 160, 2027–2032. [Google Scholar] [CrossRef]
- Dziurzynski, K.; Chang, S.M.; Heimberger, A.B.; Kalejta, R.F.; McGregor Dallas, S.R.; Smit, M.; Soroceanu, L.; Cobbs, C.S. Consensus on the role of human cytomegalovirus in glioblastoma. Neuro Oncol. 2012, 14, 246–255. [Google Scholar] [CrossRef]
- Hadaczek, P.; Ozawa, T.; Soroceanu, L.; Yoshida, Y.; Matlaf, L.; Singer, E.; Fiallos, E.; James, C.D.; Cobbs, C.S. Cidofovir: A novel antitumor agent for glioblastoma. Clin. Cancer Res. 2013, 19, 6473–6483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vliegen, I.; Duijvestijn, A.; Grauls, G.; Herngreen, S.; Bruggeman, C.; Stassen, F. Cytomegalovirus infection aggravates atherogenesis in apoE knockout mice by both local and systemic immune activation. Microbes Infect. 2004, 6, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Streblow, D.N.; Orloff, S.L.; Nelson, J.A. Do pathogens accelerate atherosclerosis? J. Nutr. 2001, 131, 2798S–2804S. [Google Scholar] [CrossRef] [Green Version]
- Hodson, E.M.; Ladhani, M.; Webster, A.C.; Strippoli, G.F.; Craig, J.C. Antiviral medications for preventing cytomegalovirus disease in solid organ transplant recipients. Cochrane Database Syst. Rev. 2013, 16, CD003774. [Google Scholar] [CrossRef]
- Owers, D.S.; Webster, A.C.; Strippoli, G.F.; Kable, K.; Hodson, E.M. Pre-emptive treatment for cytomegalovirus viraemia to prevent cytomegalovirus disease in solid organ transplant recipients. Cochrane Database Syst. Rev. 2013, 2013, CD005133. [Google Scholar] [CrossRef] [PubMed]
- Potena, L.; Holweg, C.T.; Chin, C.; Luikart, H.; Weisshaar, D.; Narasimhan, B.; Fearon, W.F.; Lewis, D.B.; Cooke, J.P.; Mocarski, E.S.; et al. Acute rejection and cardiac allograft vascular disease is reduced by suppression of subclinical cytomegalovirus infection. Transplantation 2006, 82, 398–405. [Google Scholar] [CrossRef]
- Valentine, V.G.; Weill, D.; Gupta, M.R.; Raper, B.; Laplace, S.G.; Lombard, G.A.; Bonvillain, R.W.; Taylor, D.E.; Dhillon, G.S. Ganciclovir for cytomegalovirus: A call for indefinite prophylaxis in lung transplantation. J. Heart Lung. Transplant. 2008, 27, 875–881. [Google Scholar] [CrossRef]
- Palmer, S.M.; Limaye, A.P.; Banks, M.; Gallup, D.; Chapman, J.; Lawrence, E.C.; Dunitz, J.; Milstone, A.; Reynolds, J.; Yung, G.L.; et al. Extended valganciclovir prophylaxis to prevent cytomegalovirus after lung transplantation: A randomized, controlled trial. Ann. Intern. Med. 2010, 152, 761–769. [Google Scholar] [CrossRef]
- Begos, D.G.; Rappaport, R.; Jain, D. Cytomegalovirus infection masquerading as an ulcerative colitis flare-up: Case report and review of the literature. Yale J. Biol. Med. 1996, 69, 323–328. [Google Scholar]
- Rahbar, A.; Bostrom, L.; Lagerstedt, U.; Magnusson, I.; Soderberg-Naucler, C.; Sundqvist, V.A. Evidence of active cytomegalovirus infection and increased production of IL-6 in tissue specimens obtained from patients with inflammatory bowel diseases. Inflamm. Bowel Dis. 2003, 9, 154–161. [Google Scholar] [CrossRef]
- Xu, X.; Estekizadeh, A.; Davoudi, B.; Varani, S.; Malmstrom, V.; Rahbar, A.; Soderberg-Naucler, C. Detection of human cytomegalovirus in synovial neutrophils obtained from patients with rheumatoid arthritis. Scand. J. Rheumatol. 2020, 50, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Einsele, H.; Steidle, M.; Muller, C.A.; Fritz, P.; Zacher, J.; Schmidt, H.; Saal, J.G. Demonstration of cytomegalovirus (CMV) DNA and anti-CMV response in the synovial membrane and serum of patients with rheumatoid arthritis. J. Rheumatol. 1992, 19, 677–681. [Google Scholar]
- Tsuchiya, N.; Murayama, T.; Yoshinoya, S.; Matsuta, K.; Shiota, M.; Furukawa, T.; Ito, K. Antibodies to human cytomegalovirus 65-kilodalton Fc binding protein in rheumatoid arthritis: Idiotypic mimicry hypothesis of rheumatoid factor production. Autoimmunity 1993, 15, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Choo, H.M.C.; Cher, W.Q.; Kwan, Y.H.; Fong, W.W.S. Risk factors for cytomegalovirus disease in systemic lupus erythematosus (SLE): A systematic review. Adv. Rheumatol. 2019, 59, 12. [Google Scholar] [CrossRef] [Green Version]
- Pirtskhalaishvili, G.; Nelson, J.B. Endothelium-derived factors as paracrine mediators of prostate cancer progression. Prostate 2000, 44, 77–87. [Google Scholar] [CrossRef]
- Stow, L.R.; Jacobs, M.E.; Wingo, C.S.; Cain, B.D. Endothelin-1 gene regulation. FASEB J. 2011, 25, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Bagnato, A.; Spinella, F.; Rosano, L. Emerging role of the endothelin axis in ovarian tumor progression. Endocr. Relat. Cancer 2005, 12, 761–772. [Google Scholar] [CrossRef] [Green Version]
- Rosano, L.; Spinella, F.; Di Castro, V.; Nicotra, M.R.; Dedhar, S.; de Herreros, A.G.; Natali, P.G.; Bagnato, A. Endothelin-1 promotes epithelial-to-mesenchymal transition in human ovarian cancer cells. Cancer Res. 2005, 65, 11649–11657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega Mateo, A.; de Artinano, A.A. Highlights on endothelins: A review. Pharmacol. Res. 1997, 36, 339–351. [Google Scholar] [CrossRef]
- Kedzierski, R.M.; Yanagisawa, M. Endothelin system: The double-edged sword in health and disease. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 851–876. [Google Scholar] [CrossRef]
- Grazia Revello, M.; Baldanti, F.; Percivalle, E.; Sarasini, A.; De-Giuli, L.; Genini, E.; Lilleri, D.; Labo, N.; Gerna, G. In vitro selection of human cytomegalovirus variants unable to transfer virus and virus products from infected cells to polymorphonuclear leukocytes and to grow in endothelial cells. J. Gen. Virol. 2001, 82, 1429–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafsson, R.K.; Jeffery, H.C.; Yaiw, K.C.; Wilhelmi, V.; Kostopoulou, O.N.; Davoudi, B.; Rahbar, A.; Benard, M.; Renne, T.; Soderberg-Naucler, C.; et al. Direct infection of primary endothelial cells with human cytomegalovirus prevents angiogenesis and migration. J. Gen. Virol. 2015, 96, 3598–3612. [Google Scholar] [CrossRef] [PubMed]
- Yaiw, K.C.; Mohammad, A.A.; Taher, C.; Wilhelmi, V.; Davoudi, B.; Straat, K.; Assinger, A.; Ovchinnikova, O.; Shlyakhto, E.; Rahbar, A.; et al. Human cytomegalovirus induces upregulation of arginase II: Possible implications for vasculopathies. Basic Res. Cardiol. 2014, 109, 401. [Google Scholar] [CrossRef]
- Zhang, Z.; Evers, D.L.; McCarville, J.F.; Dantonel, J.C.; Huong, S.M.; Huang, E.S. Evidence that the human cytomegalovirus IE2-86 protein binds mdm2 and facilitates mdm2 degradation. J. Virol. 2006, 80, 3833–3843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafsson, K.L.R.; Renne, T.; Soderberg-Naucler, C.; Butler, L.M. Human cytomegalovirus replication induces endothelial cell interleukin-11. Cytokine 2018, 111, 563–566. [Google Scholar] [CrossRef] [PubMed]
- McSharry, J.J.; Lurain, N.S.; Drusano, G.L.; Landay, A.L.; Notka, M.; O’Gorman, M.R.; Weinberg, A.; Shapiro, H.M.; Reichelderfer, P.S.; Crumpacker, C.S. Rapid ganciclovir susceptibility assay using flow cytometry for human cytomegalovirus clinical isolates. Antimicrob. Agents Chemother. 1998, 42, 2326–2331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohm, F.; Pernow, J. The importance of endothelin-1 for vascular dysfunction in cardiovascular disease. Cardiovasc. Res. 2007, 76, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Pascual, F.; Busnadiego, O.; Lagares, D.; Lamas, S. Role of endothelin in the cardiovascular system. Pharmacol. Res. 2011, 63, 463–472. [Google Scholar] [CrossRef]
- Thorin, E.; Clozel, M. The cardiovascular physiology and pharmacology of endothelin-1. Adv. Pharmacol. 2010, 60, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Nelson, J.; Bagnato, A.; Battistini, B.; Nisen, P. The endothelin axis: Emerging role in cancer. Nat. Rev. Cancer 2003, 3, 110–116. [Google Scholar] [CrossRef]
- Reeves, M.; Murphy, J.; Greaves, R.; Fairley, J.; Brehm, A.; Sinclair, J. Autorepression of the human cytomegalovirus major immediate-early promoter/enhancer at late times of infection is mediated by the recruitment of chromatin remodeling enzymes by IE86. J. Virol. 2006, 80, 9998–10009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Channick, R.N.; Simonneau, G.; Sitbon, O.; Robbins, I.M.; Frost, A.; Tapson, V.F.; Badesch, D.B.; Roux, S.; Rainisio, M.; Bodin, F.; et al. Effects of the dual endothelin-receptor antagonist bosentan in patients with pulmonary hypertension: A randomised placebo-controlled study. Lancet 2001, 358, 1119–1123. [Google Scholar] [CrossRef]
- Zebadua, R.; Hernandez-Perez, A.P.; Garcia, A.; Zayas, N.; Sandoval, J.; Lopez, J.; Pulido, T. Macitentan in the treatment of pulmonary arterial hypertension. Future Cardiol. 2021, 17, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, Y.; Xu, Z.; Zhang, R.; Liu, X.; Li, Y.; Mao, P. Cytomegalovirus infection and outcome in immunocompetent patients in the intensive care unit: A systematic review and meta-analysis. BMC Infect. Dis. 2018, 18, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Ke, Q.; Jin, Z.; Wang, H.; Kocher, O.; Morgan, J.P.; Zhang, J.; Crumpacker, C.S. Cytomegalovirus infection causes an increase of arterial blood pressure. PLoS Pathog. 2009, 5, e1000427. [Google Scholar] [CrossRef] [Green Version]
- Hayase, Y.; Tobita, K. Suppression of endothelin system in Vero cells latently infected with influenza virus B/Lee/40. Arch. Virol. 1999, 144, 1421–1427. [Google Scholar] [CrossRef]
- Meissner, N.N.; Koschel, K. Downregulation of endothelin receptor mRNA synthesis in C6 rat astrocytoma cells by persistent measles virus and canine distemper virus infections. J. Virol. 1995, 69, 5191–5194. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, A.; Hahn, S.; Gartner, S.; Pardo, C.A.; Netesan, S.K.; McArthur, J.; Nath, A. Molecular programming of endothelin-1 in HIV-infected brain: Role of Tat in up-regulation of ET-1 and its inhibition by statins. FASEB J. 2007, 21, 777–789. [Google Scholar] [CrossRef] [Green Version]
- Kanmogne, G.D.; Primeaux, C.; Grammas, P. Induction of apoptosis and endothelin-1 secretion in primary human lung endothelial cells by HIV-1 gp120 proteins. Biochem. Biophys. Res. Commun. 2005, 333, 1107–1115. [Google Scholar] [CrossRef]
- Speciale, L.; Biffi, R.; Mancuso, R.; Borghi, E.; Mazziotti, R.; Ferrante, P. Big endothelin-1 and interleukin-6 modulation in human microvascular endothelial cells after human herpesvirus 8 infection. Exp. Biol. Med. 2006, 231, 1171–1175. [Google Scholar]
- Vasaikar, S.; Tsipras, G.; Landazuri, N.; Costa, H.; Wilhelmi, V.; Scicluna, P.; Cui, H.L.; Mohammad, A.A.; Davoudi, B.; Shang, M.; et al. Overexpression of endothelin B receptor in glioblastoma: A prognostic marker and therapeutic target? BMC Cancer 2018, 18, 154. [Google Scholar] [CrossRef]
- Fornara, O.; Bartek, J., Jr.; Rahbar, A.; Odeberg, J.; Khan, Z.; Peredo, I.; Hamerlik, P.; Bartek, J.; Stragliotto, G.; Landazuri, N.; et al. Cytomegalovirus infection induces a stem cell phenotype in human primary glioblastoma cells: Prognostic significance and biological impact. Cell Death Differ. 2015. [Google Scholar] [CrossRef] [Green Version]
- Stragliotto, G.; Rahbar, A.; Solberg, N.W.; Lilja, A.; Taher, C.; Orrego, A.; Bjurman, B.; Tammik, C.; Skarman, P.; Peredo, I.; et al. Effects of valganciclovir as an add-on therapy in patients with cytomegalovirus-positive glioblastoma: A randomized, double-blind, hypothesis-generating study. Int. J. Cancer 2013, 133, 1204–1213. [Google Scholar] [CrossRef] [Green Version]
- Rahbar, A.; Strååt, K.; Rahbar, A.; Dzabic, M.; Söderberg-Nauclér, C.; Stragliotto, G.; Orrego, A.; Orrego, A.; Peredo, I.; Peredo, I.; et al. Human cytomegalovirus infection levels in glioblastoma multiforme are of prognostic value for survival. J. Clin. Virol. 2013, 57, 36–42. [Google Scholar] [CrossRef]
- Guruli, G.; Pflug, B.R.; Pecher, S.; Makarenkova, V.; Shurin, M.R.; Nelson, J.B. Function and survival of dendritic cells depend on endothelin-1 and endothelin receptor autocrine loops. Blood 2004, 104, 2107–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaiw, K.-C.; Mohammad, A.-A.; Taher, C.; Cui, H.L.; Costa, H.; Kostopoulou, O.N.; Jung, M.; Assinger, A.; Wilhelmi, V.; Yang, J.; et al. Human Cytomegalovirus Reduces Endothelin-1 Expression in Both Endothelial and Vascular Smooth Muscle Cells. Microorganisms 2021, 9, 1137. https://doi.org/10.3390/microorganisms9061137
Yaiw K-C, Mohammad A-A, Taher C, Cui HL, Costa H, Kostopoulou ON, Jung M, Assinger A, Wilhelmi V, Yang J, et al. Human Cytomegalovirus Reduces Endothelin-1 Expression in Both Endothelial and Vascular Smooth Muscle Cells. Microorganisms. 2021; 9(6):1137. https://doi.org/10.3390/microorganisms9061137
Chicago/Turabian StyleYaiw, Koon-Chu, Abdul-Aleem Mohammad, Chato Taher, Huanhuan Leah Cui, Helena Costa, Ourania N. Kostopoulou, Masany Jung, Alice Assinger, Vanessa Wilhelmi, Jiangning Yang, and et al. 2021. "Human Cytomegalovirus Reduces Endothelin-1 Expression in Both Endothelial and Vascular Smooth Muscle Cells" Microorganisms 9, no. 6: 1137. https://doi.org/10.3390/microorganisms9061137