
Insufficient Anthrax Lethal Toxin Neutralization Is Associated with Antibody Subclass and Domain Specificity in the Plasma of Anthrax-Vaccinated Individuals


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Human Blood Samples
2.2. Anti-PA IgG (Total and Subclass) Concentration
2.3. LT Neutralization Activity (LTNA)
2.4. Anti-PA IgG Avidity
2.5. Recombinant Protective Antigen Domains and ELISA
2.6. Statistical Analysis
3. Results
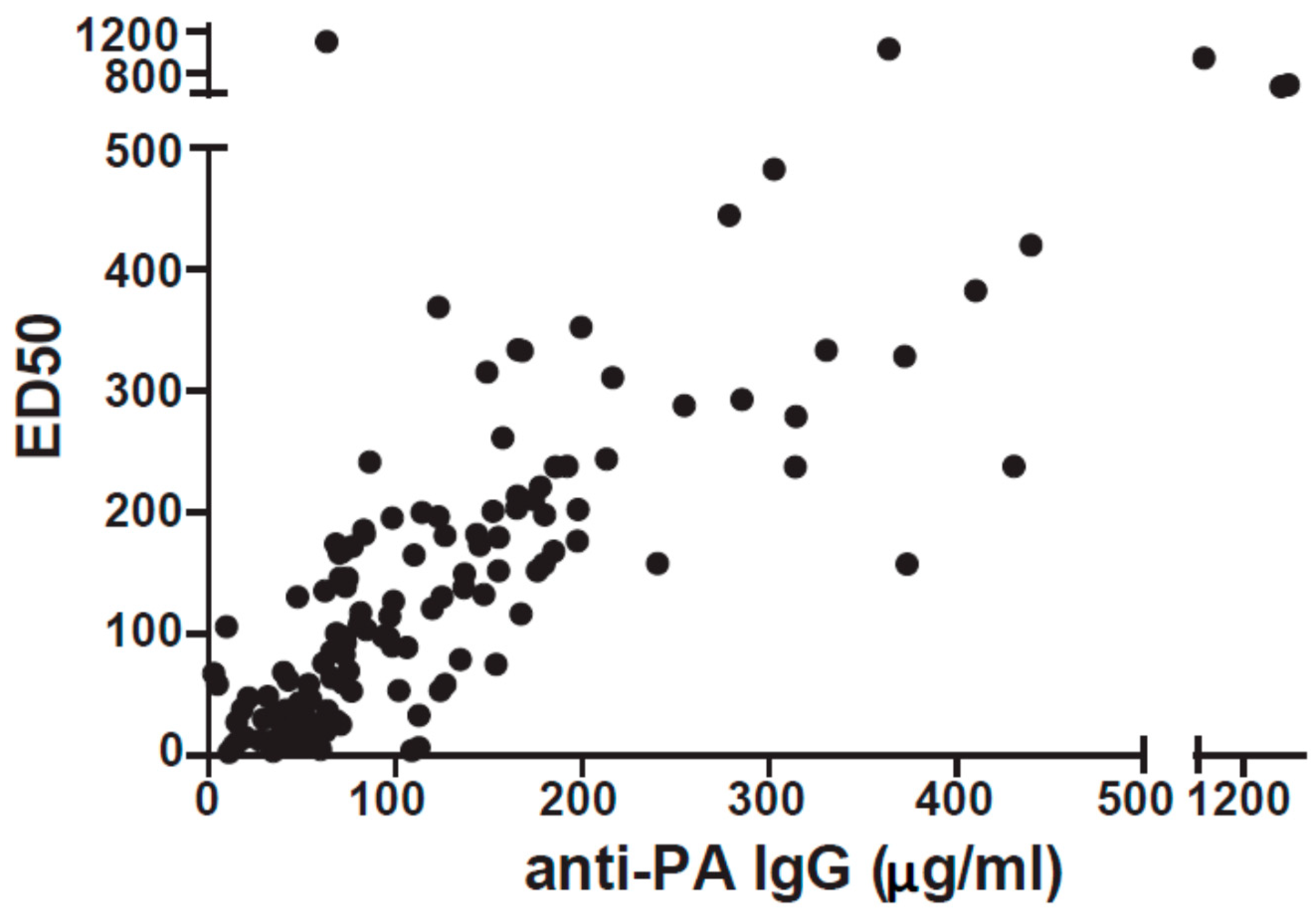
3.1. Over Half of AVA Vaccinees Produce Insufficient Quantity and/or Quality of Anti-PA
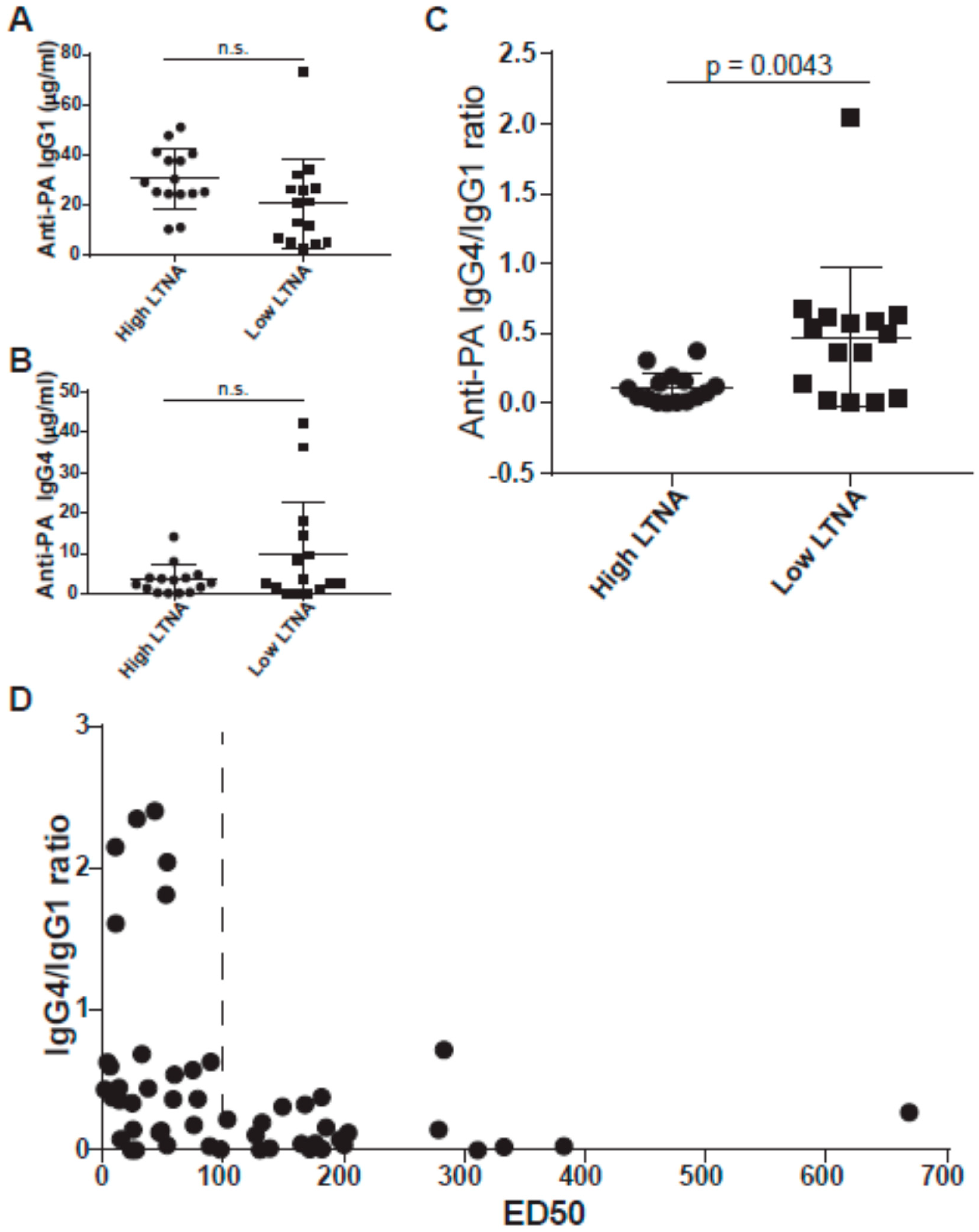
3.2. IgG4/IgG1 Ratios Are Associated with Neutralization
3.3. Anti-PA Avidity Is Associated with the Number of AVA Vaccinations
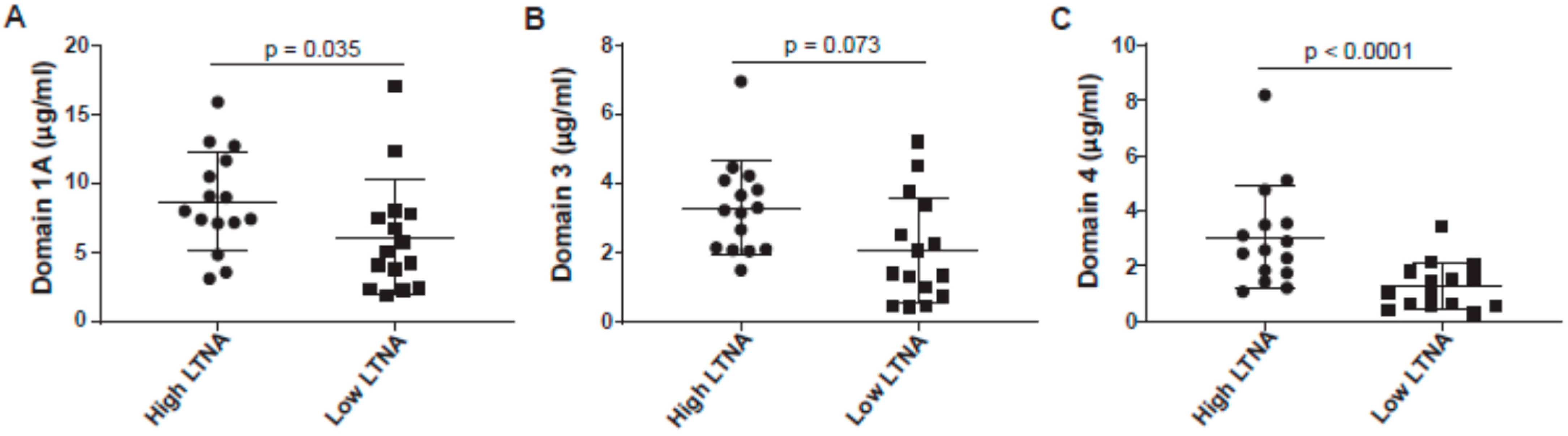
3.4. Anti-Domain 4 Is Strongly Associated with Neutralization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- WHO. Anthrax in Humans and Animals, 4th ed.; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Mourez, M. Anthrax toxins. Rev. Physiol. Biochem. Pharmacol. 2004, 152, 135–164. [Google Scholar]
- Pitt, M.L.M.; Little, S.; Ivins, B.E.; Fellows, P.; Boles, J.; Barth, J.; Hewetson, J.; Friedlander, A.M. In vitro correlate of immunity in an animal model of inhalational anthrax. J. Appl. Microbiol. 1999, 87, 304. [Google Scholar] [CrossRef]
- Reuveny, S.; White, M.D.; Adar, Y.Y.; Kafri, Y.; Altboum, Z.; Gozes, Y.; Kobiler, D.; Shafferman, A.; Velan, B. Search for correlates of protective immunity conferred by anthrax vaccine. Infect. Immun. 2001, 69, 2888–2893. [Google Scholar] [CrossRef] [Green Version]
- Goossens, P.L. Animal models of human anthrax: The Quest for the Holy Grail. Mol. Asp. Med. 2009, 30, 467–480. [Google Scholar] [CrossRef]
- Little, S.F.; Ivins, B.E.; Fellows, P.F.; Friedlander, A.M. Passive protection by polyclonal antibodies against Bacillus anthracis infection in guinea pigs. Infect. Immun. 1997, 65, 5171–5175. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, P.C. Anthrax vaccines: Past, present and future. Vaccine 1991, 9, 533–539. [Google Scholar] [CrossRef]
- Marano, N.; Plikaytis, B.D.; Martin, S.W.; Rose, C.; Semenova, V.A.; Martin, S.K.; Freeman, A.E.; Li, H.; Mulligan, M.J.; Parker, S.D.; et al. Effects of a reduced dose schedule and intramuscular administration of anthrax vaccine adsorbed on immunogenicity and safety at 7 months: A randomized trial. JAMA 2008, 300, 1532–1543. [Google Scholar] [CrossRef]
- Quinn, C.P.; Sabourin, C.L.; Niemuth, N.A.; Li, H.; Semenova, V.A.; Rudge, T.L.; Mayfield, H.J.; Schiffer, J.; Mittler, R.S.; Ibegbu, C.C.; et al. A three-dose intramuscular injection schedule of anthrax vaccine adsorbed generates sustained humoral and cellular immune responses to protective antigen and provides long-term protection against inhalation anthrax in rhesus macaques. Clin. Vaccine Immunol. 2012, 19, 1730–1745. [Google Scholar] [CrossRef] [Green Version]
- Fay, M.P.; Follmann, D.A.; Lynn, F.; Schiffer, J.M.; Stark, G.V.; Kohberger, R.; Quinn, C.P.; Nuzum, E.O. Anthrax vaccine-induced antibodies provide cross-species prediction of survival to aerosol challenge. Sci. Transl. Med. 2012, 4, 151ra26. [Google Scholar] [CrossRef] [Green Version]
- Brenneman, K.E.; Doganay, M.; Akmal, A.; Goldman, S.; Galloway, D.R.; Mateczun, A.J.; Cross, A.S.; Baillie, L.W. The early humoral immune response to Bacillus anthracis toxins in patients infected with cutaneous anthrax. FEMS Immunol. Med. Microbiol. 2011, 62, 164–172. [Google Scholar] [CrossRef] [Green Version]
- Boyer, A.E.; Quinn, C.P.; Beesley, C.A.; Gallegos-Candela, M.; Marston, C.K.; Cronin, L.X.; Lins, R.C.; Stoddard, R.A.; Li, H.; Schiffer, J.; et al. Lethal factor toxemia and anti-protective antigen antibody activity in naturally acquired cutaneous anthrax. J. Infect. Dis. 2011, 204, 1321–1327. [Google Scholar] [CrossRef]
- Charlton, S.; Herbert, M.; McGlashan, J.; King, A.; Jones, P.; West, K.; Roberts, A.; Silman, N.; Marks, T.; Hudson, M.; et al. A study of the physiology of Bacillus anthracis sterne during manufacture of the UK acellular anthrax vaccine. J. Appl. Microbiol. 2007, 103, 1453–1460. [Google Scholar] [CrossRef]
- Dumas, E.K.; Garman, L.; Cuthbertson, H.; Charlton, S.; Hallis, B.; Engler, R.J.; Choudhari, S.; Picking, W.D.; James, J.A.; Farris, A.D. Lethal factor antibodies contribute to lethal toxin neutralization in recipients of anthrax vaccine precipitated. Vaccine 2017, 35, 3416–3422. [Google Scholar] [CrossRef]
- Baillie, L.; Townend, T.; Walker, N.; Eriksson, U.; Williamson, D. Characterization of the human immune response to the UK anthrax vaccine. FEMS Immunol. Med. Microbiol. 2004, 42, 267–270. [Google Scholar] [CrossRef]
- Crowe, S.R.; Ash, L.L.; Engler, R.J.M.; Ballard, J.D.; Harley, J.B.; Farris, A.D.; James, J.A. Select human anthrax protective antigen epitope-specific antibodies provide protection from lethal toxin challenge. J. Infect. Dis. 2010, 202, 251–260. [Google Scholar] [CrossRef]
- Crowe, S.R.; Garman, L.; Engler, R.J.; Farris, A.D.; Ballard, J.D.; Harley, J.B.; James, J.A. Anthrax vaccination induced anti-lethal factor IgG: Fine specificity and neutralizing capacity. Vaccine 2011, 29, 3670–3678. [Google Scholar] [CrossRef] [Green Version]
- Aalberse, R.C.; Stapel, S.O.; Schuurman, J.; Rispens, T. Immunoglobulin G4: An odd antibody. Clin. Exp. Allergy 2009, 39, 469–477. [Google Scholar] [CrossRef]
- Abboud, N.; Chow, S.K.; Saylor, C.; Janda, A.; Ravetch, J.V.; Scharff, M.D.; Casadevall, A. A requirement for FcγR in antibody-mediated bacterial toxin neutralization. J. Exp. Med. 2010, 207, 2395–2405. [Google Scholar] [CrossRef] [Green Version]
- Bruhns, P.; Iannascoli, B.; England, P.; Mancardi, D.A.; Fernandez, N.; Jorieux, S.; Daëron, M. Specificity and affinity of human Fcgamma receptors and their polymorphic variants for human IgG subclasses. Blood 2009, 113, 3716–3725. [Google Scholar] [CrossRef]
- Murphy, K.; Travers, P.; Walport, M. Janeway’s Immunobiology, 7th ed.; Garland Science: New York, NY, USA, 2008; pp. 379–417. [Google Scholar]
- Cook, D.A.; Owen, T.; Wagstaff, S.C.; Kinne, J.; Wernery, U.; Harrison, R.A. Analysis of camelid antibodies for antivenom development: Neutralisation of venom-induced pathology. Toxicon Off. J. Int. Soc. Toxinol. 2010, 56, 373–380. [Google Scholar] [CrossRef]
- Aref, N.E.; Saeed, A.M. Generation of high-titer of neutralizing polyclonal antibodies against heat-stable enterotoxin (STa) of enterotoxigenic Escherichia coli. Vaccine 2012, 30, 6341–6346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reason, D.; Liberato, J.; Sun, J.; Keitel, W.; Zhou, J. Frequency and domain specificity of toxin-neutralizing paratopes in the human antibody response to anthrax vaccine adsorbed. Infect. Immun. 2009, 77, 2030–2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.; Crowe, S.R.; Garman, L.; Guthridge, C.J.; Muther, J.J.; McKee, E.; Zheng, N.-Y.; Farris, A.D.; Guthridge, J.M.; Wilson, P.C.; et al. Human monoclonal antibodies generated following vaccination with AVA provide neutralization by blocking furin cleavage but not by preventing oligomerization. Vaccine 2012, 30, 4276–4283. [Google Scholar] [CrossRef] [Green Version]
- Manish, M.; Rahi, A.; Kaur, M.; Bhatnagar, R.; Singh, S. A single-dose PLGA encapsulated protective antigen domain 4 nanoformulation protects mice against Bacillua anthracis spore challenge. PLoS ONE 2013, 8, e61885. [Google Scholar] [CrossRef] [Green Version]
- Gogoi, H.; Mani, R.; Aggarwal, S.; Malik, A.; Munde, M.; Bhatnagar, R. Crystalline and amorphous preparation of aluminum hydroxide nanoparticles enchances protective antigen domain 4 specific immunogeniticity and proves protection against anthrax. Int. J. Nanomed. 2020, 15, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Quinn, C.P.; Semenova, V.A.; Elie, C.M.; Romero-Steiner, S.; Greene, C.; Li, H.; Stamey, K.; Steward-Clark, E.; Schmidt, D.S.; Mothershed, E.; et al. Specific, sensitive, and quantitative enzyme-linked immunosorbent assay for human immunoglobulin G antibodies to anthrax toxin protective antigen. Emerg. Infect. Dis. 2002, 8, 1103–1110. [Google Scholar] [CrossRef]
- Li, H.; Soroka, S.D.; Taylor, T.H., Jr.; Stamey, K.L.; Stinson, K.W.; Freeman, A.E.; Abramson, D.R.; Desai, R.; Cronin, L.X.; Oxford, J.W.; et al. Strandarized, mathmatical model-based and validate in vitro analysis of anthrax lethal toxin neutralization. J. Immunol. Methods 2008, 333, 89–106. [Google Scholar] [CrossRef]
- Soroka, S.D.; Schiffer, J.M.; Semenova, V.A.; Li, H.; Foster, L.; Quinn, C.P. A two-strage, multilevel quality control system for serological assays in anthrax vaccine clinical trials. Biologicals 2010, 38, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Flick-Smith, H.C.; Walker, N.J.; Gibson, P.; Bullifent, J.; Hayward, S.; Miller, J.; Titball, R.W.; Williamson, E.D. A recombinant carboxy-terminal domain of the protective antigen of Bacilius anthracis protects mice against anthrax infection. Infect. Immun. 2002, 70, 1653–1656. [Google Scholar] [CrossRef] [Green Version]
- Quinn, C.P.; Sabourin, C.L.; Schiffer, J.M.; Niemuth, N.A.; Semenova, V.A.; Li, H.; Rudge, T.L.; Brys, A.M.; Mittler, R.S.; Ibegbu, C.C.; et al. Humoral and cell-mediated immune responses to alternate booster schedules of anthrax vaccine adsorbed in humans. Clin. Vaccine Immunol. 2016, 23, 326–338. [Google Scholar] [CrossRef] [Green Version]
- Garman, L.; Smith, K.; Farris, A.D.; Nelson, M.R.; Engler, R.J.M.; James, J.A. Protective antigen-specific memory B cells persist years after anthrax vaccination and correlate with humoral immunity. Toxins 2014, 6, 2424–2431. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, E.; Coban, C.; Ishii, K.J. Particulate adjuvant and innate immunity: Past achievements, present findings, and future prospects. Int. Rev. Immunol. 2013, 32, 209–220. [Google Scholar] [CrossRef]
- James, L.K.; Till, S.J. Potential mechanisms for IgG4 inhibition of immediate hypersensitivity reactions. Curr. Allergy Asthma Rep. 2016, 16, 23. [Google Scholar] [CrossRef] [Green Version]
- Akdis, M. Healthy immune response to allergens: T regulatory cells and more. Curr. Opin. Immunol. 2006, 18, 738–744. [Google Scholar] [CrossRef]
- Garman, L.; Smith, K.; Muns, E.E.; Velte, C.A.; Spooner, C.E.; Munroe, M.E.; Farris, A.D.; Nelson, M.R.; Engler, R.J.M.; James, J.A. Unique inflammatory mediators and specific IgE levels distinguish local from system reactions after anthrax vaccine adsorbed vaccination. Clin. Vaccine Immunol. 2016, 23, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Victora, G.D.; Nussenzweig, M.C. Germinal centers. Annu. Rev. Immunol. 2012, 30, 429–457. [Google Scholar] [CrossRef]
- Zhang, Y.; Meyer-Hermann, M.; George, L.A.; Figge, M.T.; Khan, M.; Goodall, M.; Young, S.P.; Reynolds, A.; Falciani, F.; Waisman, A.; et al. Germinal center B cells govern their own fate via antibody feedback. J. Exp. Med. 2013, 210, 457–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfeld, R.; Alacalay, R.; Mechaly, A.; Lapidoth, G.; Epstein, E.; Kronman, C.; Fleishman, S.J.; Mazor, M.O. Improved antibody-based ricin neutralization by affinity maturation is correlated with slower off-rate values. Protein Eng. Des. Sel. 2017, 30, 611–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brossier, F.; Weber-Levy, M.; Mock, M.; Sirard, J.C. Role of toxin functional domains in anthrax pathogenesis. Infect. Immun. 2000, 68, 1781–1786. [Google Scholar] [CrossRef] [Green Version]
- Kelly-Cirino, C.D.; Mantis, N.J. Neutralizing monoclonal antibodies directed against defined linear epitopes on domain 4 of anthrax protective antigen. Infect. Immun. 2009, 77, 4859–4867. [Google Scholar] [CrossRef] [Green Version]
- Little, S.F.; Novak, J.M.; Lowe, J.R.; Leppla, S.H.; Singh, Y.; Klimpel, K.R.; Lidgerding, B.C.; Friedlander, A.M. Characterization of lethal factor binding and cell receptor binding domains of protective antigen of Bacillus anthracis using monoclonal antibodies. Microbiology 1996, 142 Pt 3, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Rosovitz, M.J.; Schuck, P.; Varughese, M.; Chopra, A.P.; Mehra, V.; Singh, Y.; McGinnis, L.M.; Leppla, S.H. Alanine-scanning mutations in domain 4 of anthrax toxin protective antigen reveal residues important for binding to the cellular receptor and to a neutralizing monoclonal antibody. J. Biol. Chem. 2003, 278, 30936–30944. [Google Scholar] [CrossRef] [Green Version]
- Abboud, N.; Casadevall, A. Immunogenicity of Bacillus anthracis protective antigen domains and efficacy of elicited antibody responses depend on host genetic background. Clin. Vaccine Immunol. 2008, 15, 1115–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| High LTNA | Low LTNA | |
|---|---|---|
| Gender: | ||
| M (%) | 73.0% | 100% |
| Race: | ||
| European American (%) | 93.3% (1 A) | 86.7% (1 AA, 1 AI) |
| Age at collection: | ||
| Average (SEM) | 31.7 (2.29) | 31.1 (1.37) |
| Median | 31 | 31 |
| Range | 20–47 | 22–40 |
| Number of vaccinations: | ||
| Average (SEM) | 6.33 (0.48) | 6.4 (0.43) |
| Median | 6 | 6 |
| Range | 4–9 | 4–9 |
| Years since last vaccination: | ||
| Average (SEM) | 1.12 (0.27) | 1.09 (0.28) |
| Median | 0.88 | 0.73 |
| Range | 0.18–3.67 | 0.09–3.57 |
| Anti-PA: | ||
| Average (SEM) | 117.2 (9.35) | 105.9 (6.01) |
| Median | 114.3 | 106.2 |
| Range | 73.37–197.4 | 70.5–154.0 |
| Donor | Anti-PA (µg/mL) | ED50 | Age | Total # of AVA Vaccinations | Years Post Last Boost | Possible Reason for Low LTNA | ||
|---|---|---|---|---|---|---|---|---|
| High IgG4/IgG1 Ratio (>0.5) | Low Avidity (<1.5 AI) | Low Anti- Domain 4 (<1.1 µg/mL) | ||||||
| Vaccinee 1 | 106.2 | 89.0 | 28 | 5 | 0.79 | 0.03 | 2.04 | 2.14 |
| Vaccinee 2 | 108.8 | 3.9 | 23 | 4 | 0.56 | 0.62 | 1.54 | 0.55 |
| Vaccinee 3 | 126.4 | 58.4 | 33 | 4 | 3.57 | 0.36 | 1.24 | 0.61 |
| Vaccinee 4 | 112.6 | 6.4 | 25 | 5 | 0.21 | 0.59 | 1.63 | 0.29 |
| Vaccinee 5 | 93.3 | 97.7 | 30 | 6 | 2.42 | 0.01 | 2.76 | 1.81 |
| Vaccinee 6 | 134.8 | 78.8 | 32 | 7 | 1.89 | 0.36 | 2.43 | 0.95 |
| Vaccinee 7 | 124.0 | 53.5 | 40 | 6 | 0.18 | 2.04 | 2.04 | 1.50 |
| Vaccinee 8 | 98.2 | 89.7 | 22 | 5 | 0.14 | 0.63 | 1.21 | 1.42 |
| Vaccinee 9 | 102.0 | 53.5 | 31 | 6 | 0.13 | 0.04 | 0.74 | 0.64 |
| Vaccinee 10 | 112.5 | 32.6 | 37 | 6 | 0.09 | 0.68 | 1.34 | 1.00 |
| Vaccinee 11 | 96.4 | 96.9 | 30 | 8 | 0.73 | 0.01 | 1.29 | 2.04 |
| Vaccinee 12 | 72.0 | 59.5 | 39 | 8 | 1.60 | 0.54 | 1.12 | 0.42 |
| Vaccinee 13 | 70.5 | 7.3 | 33 | 9 | 1.38 | 0.15 | 2.30 | 1.55 |
| Vaccinee 14 | 154.0 | 74.7 | 34 | 8 | 0.36 | 0.57 | 2.32 | 3.42 |
| Vaccinee 15 | 76.5 | 2.2 | 30 | 9 | 2.37 | 0.50 | 1.74 | 0.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, K.; Garman, L.; Norris, K.; Muther, J.; Duke, A.; Engler, R.J.M.; Nelson, M.R.; Collins, L.C.; Spooner, C.; Guthridge, C.; et al. Insufficient Anthrax Lethal Toxin Neutralization Is Associated with Antibody Subclass and Domain Specificity in the Plasma of Anthrax-Vaccinated Individuals. Microorganisms 2021, 9, 1204. https://doi.org/10.3390/microorganisms9061204
Smith K, Garman L, Norris K, Muther J, Duke A, Engler RJM, Nelson MR, Collins LC, Spooner C, Guthridge C, et al. Insufficient Anthrax Lethal Toxin Neutralization Is Associated with Antibody Subclass and Domain Specificity in the Plasma of Anthrax-Vaccinated Individuals. Microorganisms. 2021; 9(6):1204. https://doi.org/10.3390/microorganisms9061204
Chicago/Turabian StyleSmith, Kenneth, Lori Garman, Kathleen Norris, Jennifer Muther, Angie Duke, Renata J. M. Engler, Michael R. Nelson, Limone C. Collins, Christina Spooner, Carla Guthridge, and et al. 2021. "Insufficient Anthrax Lethal Toxin Neutralization Is Associated with Antibody Subclass and Domain Specificity in the Plasma of Anthrax-Vaccinated Individuals" Microorganisms 9, no. 6: 1204. https://doi.org/10.3390/microorganisms9061204