
Coxsackievirus B4 Transplacental Infection Severely Disturbs Central Tolerogenic Mechanisms in the Fetal Thymus



 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus
2.2. Mice
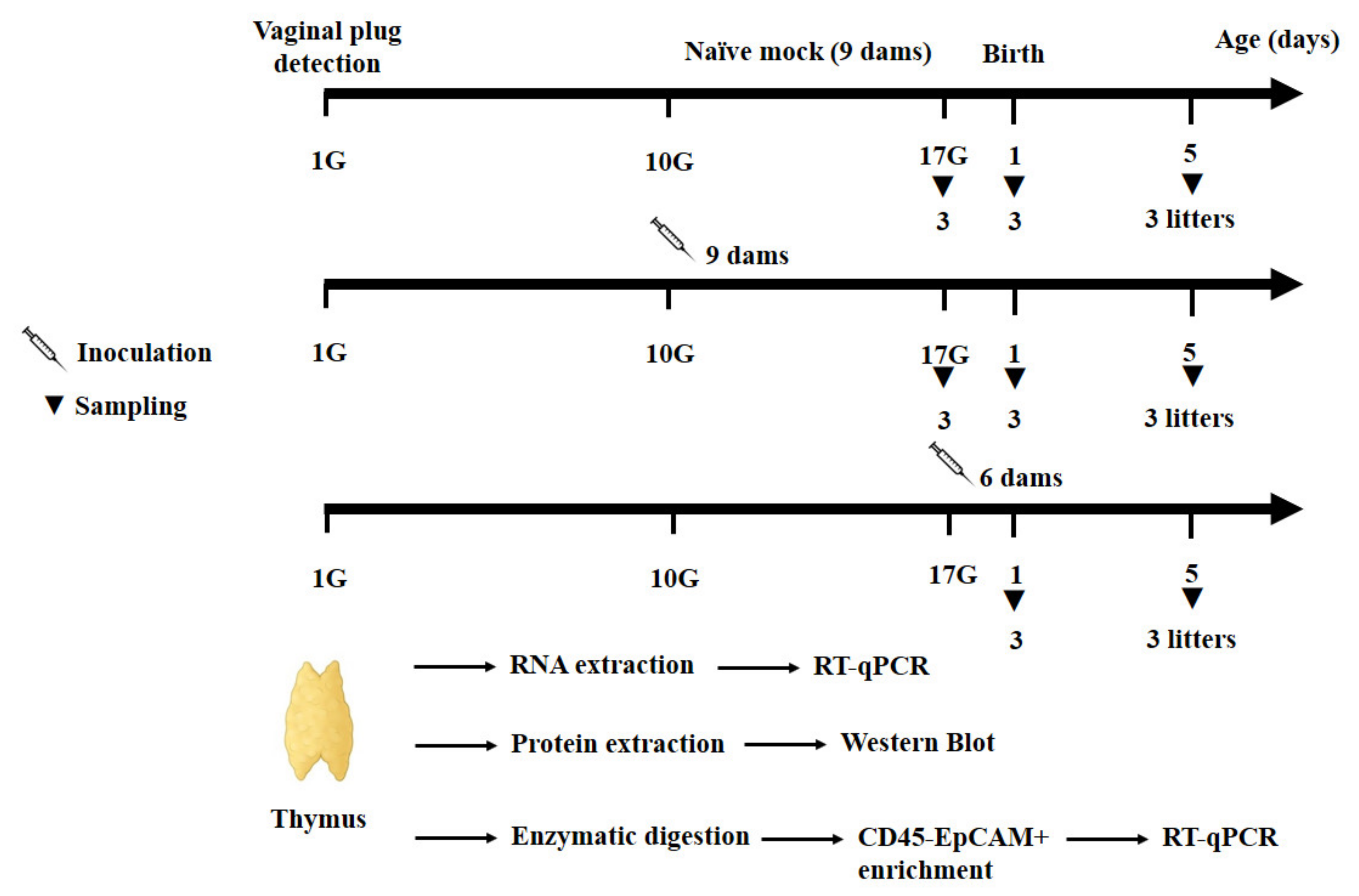
2.3. Mice Inoculation and Follow-Up
2.4. TECs Isolation and Immunostaining
2.5. RNA Extraction
2.6. Complementary DNA (cDNA) Synthesis
2.7. Viral Load Quantification
2.8. Viral Genome Detection in Enriched TECs
2.9. Car Expression
2.10. qPCR Assay
2.11. Western Blotting Analysis
2.12. Statistical Analysis
3. Results
3.1. In Utero CV-B4 Infection of Offspring’s Thymuses
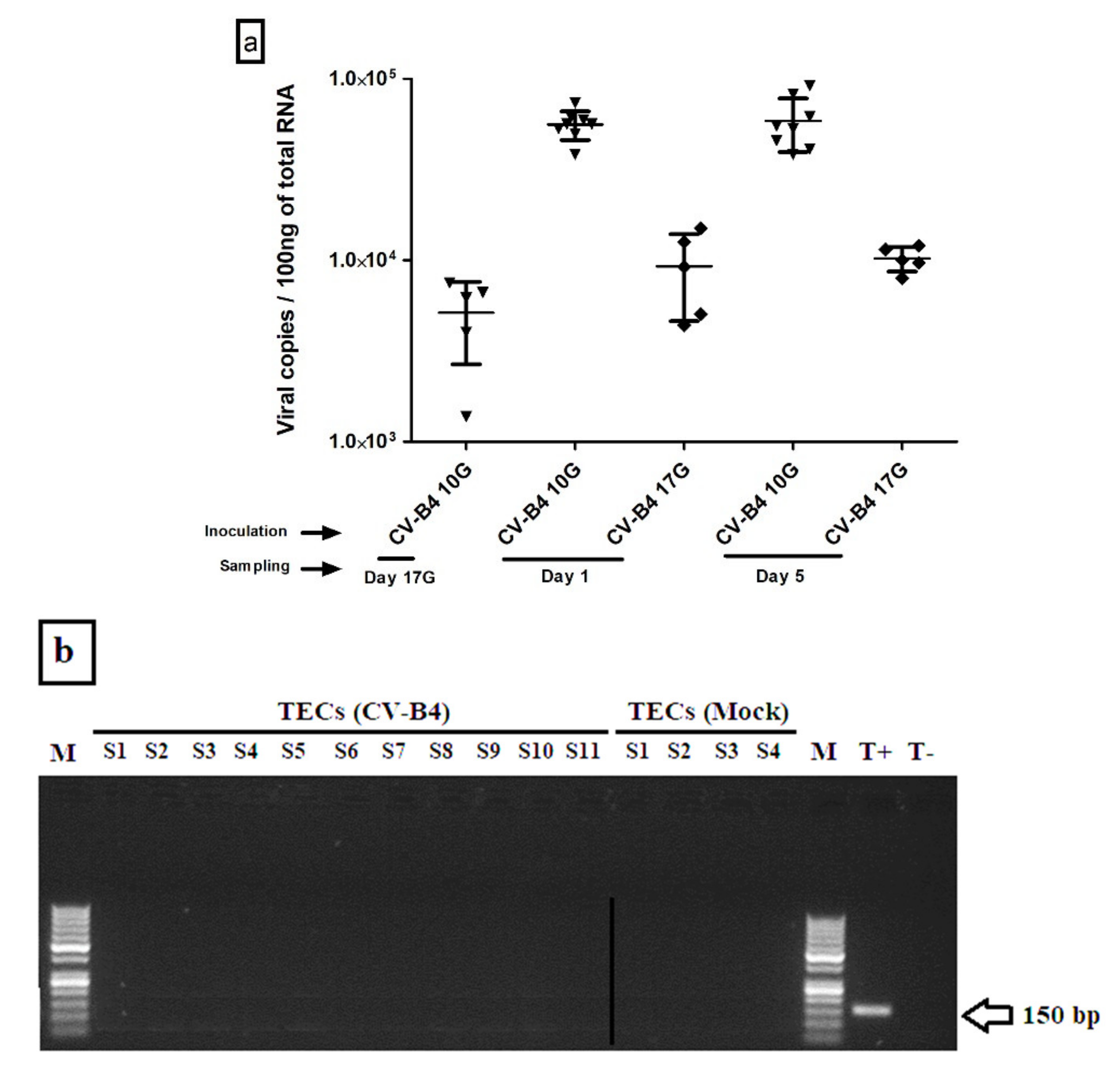
3.1.1. Viral Load in Whole Thymus
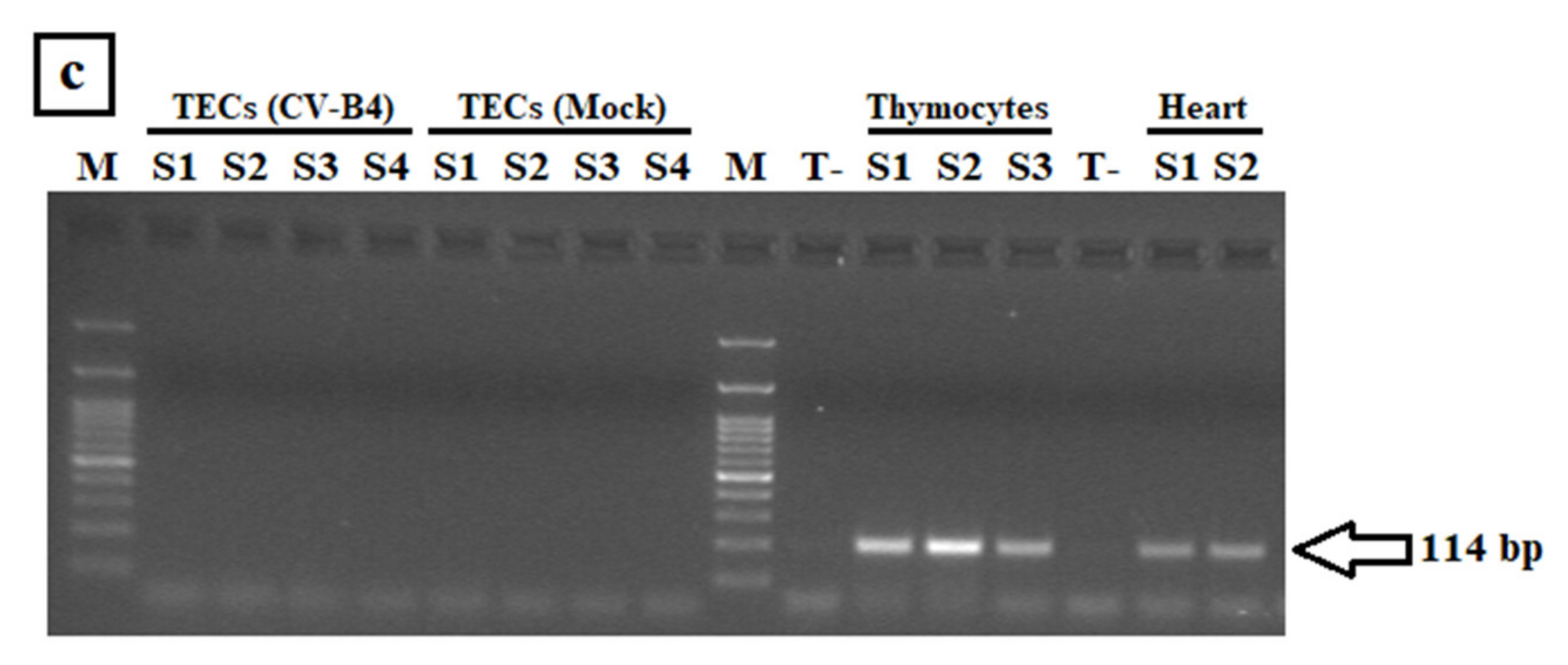
3.1.2. CV-B4 RNA Detection in Enriched TECs
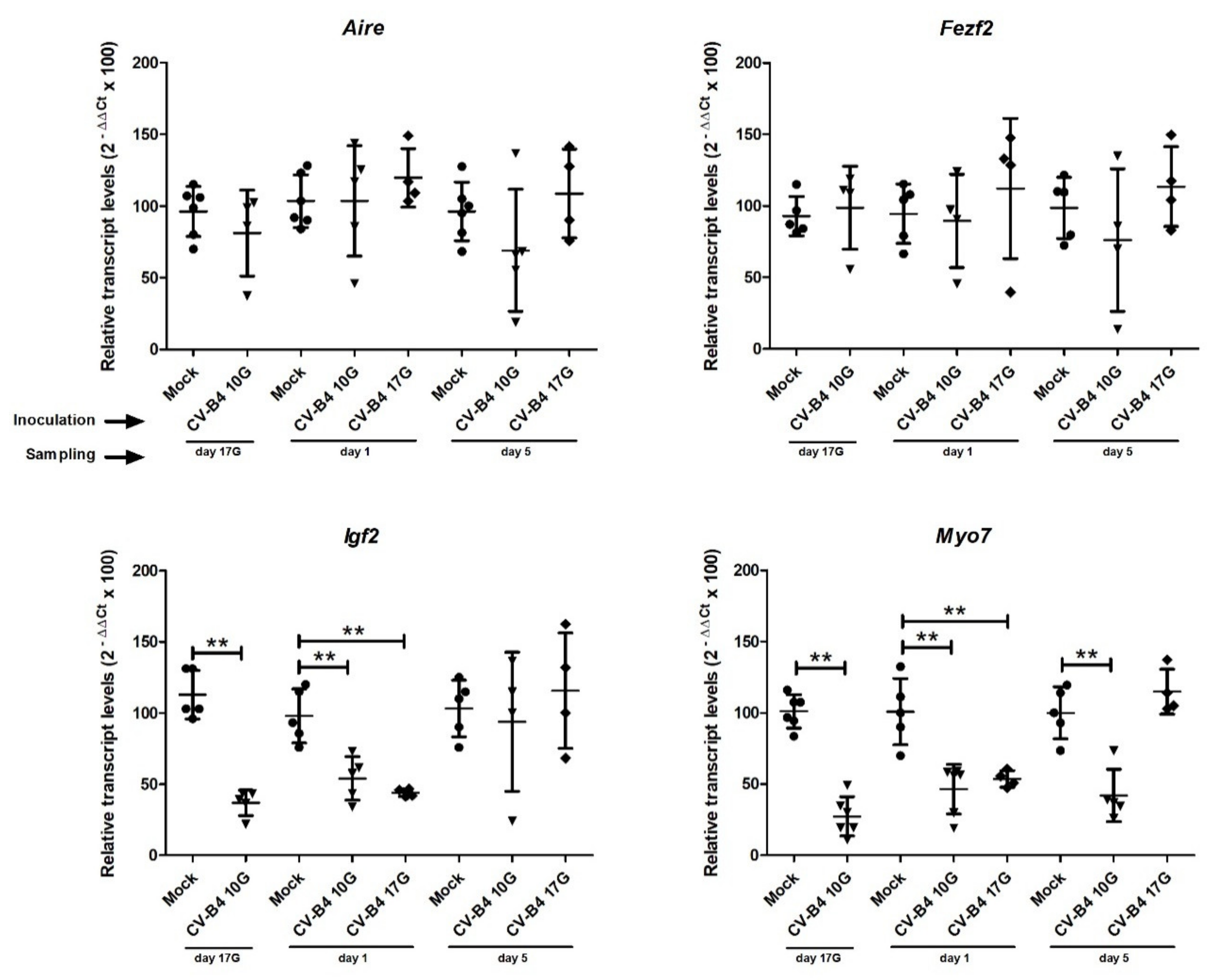
3.2. Transcription Factors and Autoantigens Transcripts in Whole Thymus
3.3. Transcription Factors and Autoantigens Transcripts in Enriched TECs
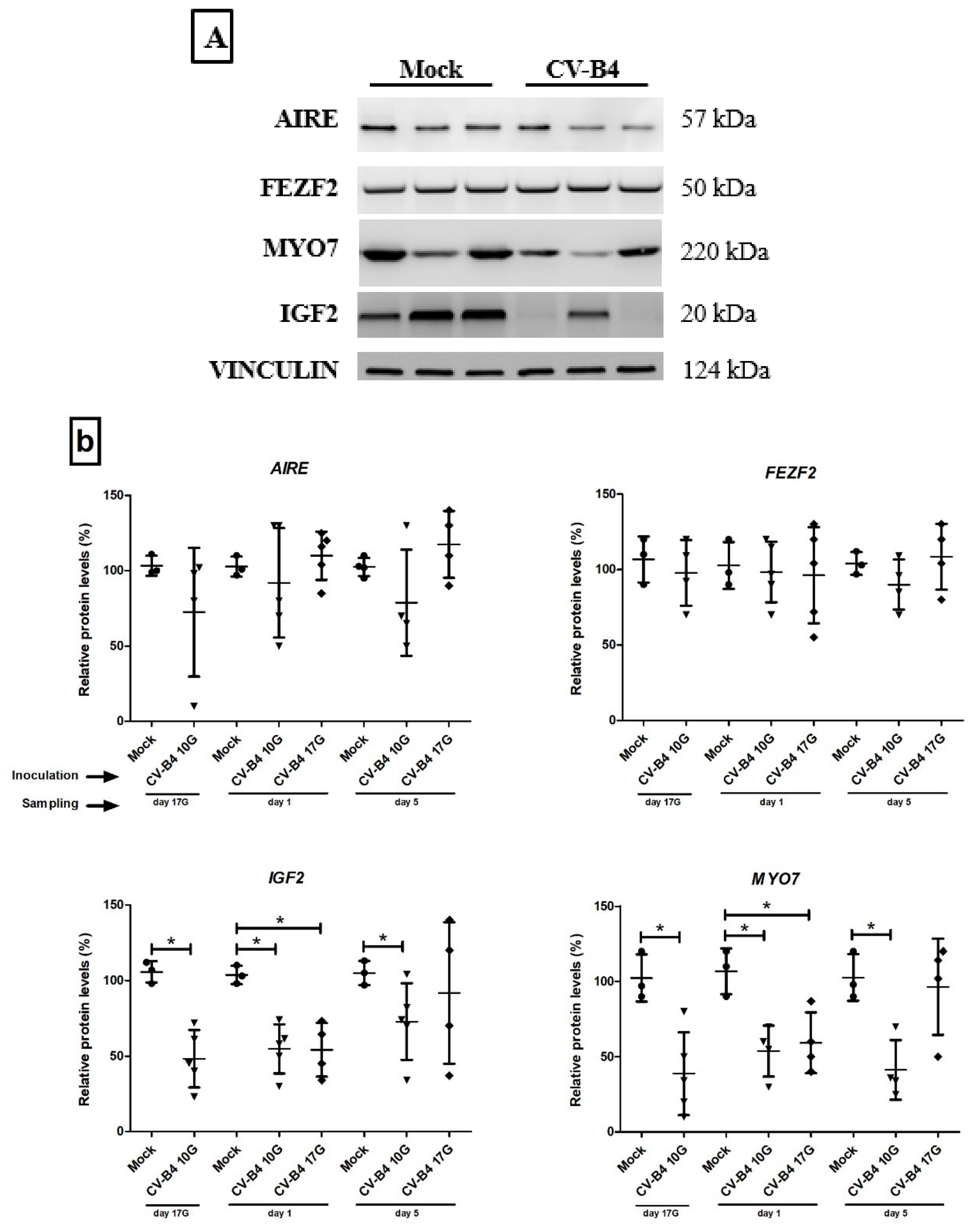
3.4. Western Blot Analysis for Transcription Factors and Autoantigens Proteins
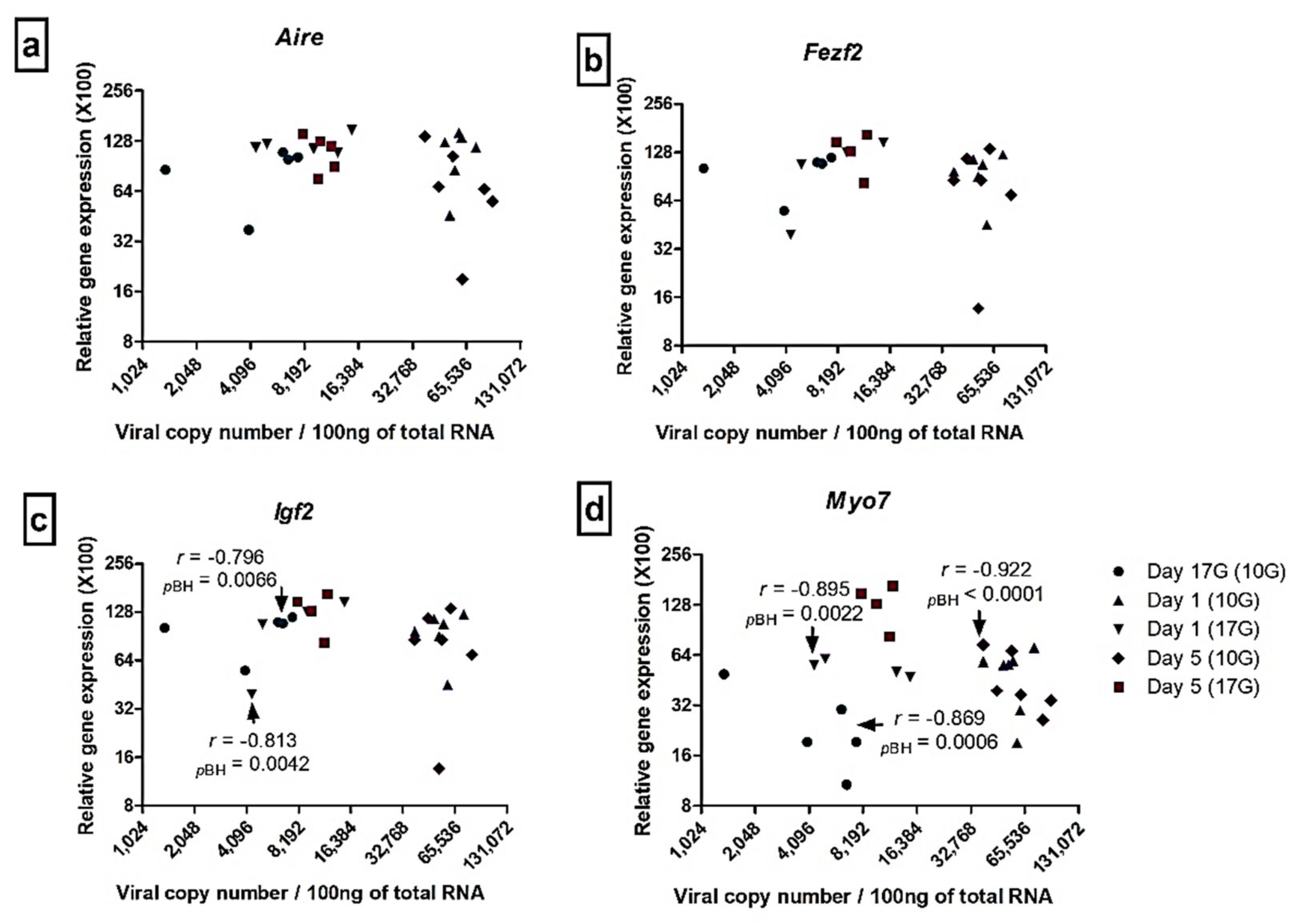
3.5. Correlation between Viral Load and Transcription Factors and Autoantigens Transcripts Levels in the Thymus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, J.F. Burnet oration. The thymus then and now. Immunol. Cell Biol. 1994, 72, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.F. The golden anniversary of the thymus. Nat. Rev. Immunol. 2011, 11, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Sebzda, E.; Mariathasan, S.; Ohteki, T.; Jones, R.; Bachmann, M.F.; Ohashi, P.S. Selection of the T cell repertoire. Annu. Rev. Immunol. 1999, 17, 829–874. [Google Scholar] [CrossRef]
- Takahama, Y. Journey through the thymus, stromal guides for T-cell development and selection. Nat. Rev. Immunol. 2006, 6, 127. [Google Scholar] [CrossRef]
- Benoist, C.; Mathis, D. Positive selection of the T cell repertoire, where and when does it occur? Cell 1989, 58, 1027–1033. [Google Scholar] [CrossRef]
- Van den Berg, H.A.; Molina-París, C. Thymic presentation of autoantigens and the efficiency of negative selection. Comput. Math. Methods Med. 2003, 5, 1–22. [Google Scholar] [CrossRef]
- Klein, L. Aire gets company for immune tolerance. Cell 2015, 163, 794–795. [Google Scholar] [CrossRef][Green Version]
- Fairweather, D.; Stafford, K.A.; Sung, Y.K. Update on coxsackievirus B3 myocarditis. Curr. Opin. Rheumatol. 2012, 24, 401–407. [Google Scholar] [CrossRef]
- Jaïdane, H.; Sauter, P.; Sané, F.; Goffard, A.; Gharbi, J.; Hober, D. Enteroviruses and type 1 diabetes, towards a better understanding of the relationship. Rev. Med. Virol. 2010, 20, 265–280. [Google Scholar] [CrossRef]
- Triantafyllopoulou, A.; Moutsopoulos, H.M. Autoimmunity and coxsackievirus infection in primary Sjögren’s syndrome. Ann. N. Y. Acad. Sci. 2005, 1050, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Tracy, S.; Gauntt, C. Group B coxsackievirus virulence. In Group B Coxsackieviruses; Springer: Berlin/Heidelberg, Germany, 2008; pp. 49–63. [Google Scholar]
- Molina, V.; Shoenfeld, Y. Infection, vaccines and other environmental triggers of autoimmunity. Autoimmunity 2005, 38, 235–245. [Google Scholar] [CrossRef]
- Tsai, S.; Santamaria, P. MHC class II polymorphisms, autoreactive T-cells, and autoimmunity. Front. Immunol. 2013, 4, 321. [Google Scholar] [CrossRef]
- Geenen, V.; Bodart, G.; Henry, S.; Michaux, H.; Dardenne, O.; Charlet-Renard, C.; Martens, H.; Hober, D. Programming of neuroendocrine self in the thymus and its defect in the development of neuroendocrine autoimmunity. Front. Neurosci. 2013, 7, 187. [Google Scholar] [CrossRef]
- Savino, W. The thymus is a common target organ in infectious diseases. PLoS Pathog. 2006, 2, e62. [Google Scholar] [CrossRef]
- Nunes-Alves, C.; Nobrega, C.; Behar, S.M.; Correia-Neves, M. Tolerance has its limits, how the thymus copes with infection. Trends Immunol. 2013, 34, 502–510. [Google Scholar] [CrossRef]
- Jaïdane, H.; Sané, F.; Hiar, R.; Goffard, A.; Gharbi, J.; Geenen, V.; Hober, D. Immunology in the clinic review series, focus on type 1 diabetes and viruses, enterovirus, thymus and type 1 diabetes pathogenesis. Clin. Exp. Immunol. 2012, 168, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Jaïdane, H.; Sané, F.; Gharbi, J.; Aouni, M.; Romond, M.B.; Hober, D. Coxsackievirus B4 and type 1 diabetes pathogenesis: Contribution of animal models. Diabetes Metab. Res. Rev. 2009, 25, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Jaïdane, H.; Gharbi, J.; Lobert, P.E.; Lucas, B.; Hiar, R.; M’Hadheb, M.B.; Brilot, F.; Geenen, V.; Aouni, M.; Hober, D. Prolonged viral RNA detection in blood and lymphoid tissues from coxsackievirus B4 E2 orally-inoculated Swiss mice. Microbiol. Immunol. 2006, 50, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Kibrick, S.; Benirschke, K. Acute aseptic myocarditis and meningoencephalitis in the newborn child infected with Coxsackie virus group B, type 3. N. Engl. J. Med. 1956, 255, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Jaïdane, H.; Gharbi, J.; Lobert, P.E.; Caloone, D.; Lucas, B.; Sané, F.; Dziorek, T.; Romond, M.B.; Aouni, M.; Hober, D. Infection of primary cultures of murine splenic and thymic cells with coxsackievirus B4. Microbiol. Immunol. 2008, 52, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Brilot, F.; Chehadeh, W.; Charlet-Renard, C.; Martens, H.; Geenen, V.; Hober, D. Persistent infection of human thymic epithelial cells by coxsackievirus B4. J. Virol. 2002, 76, 5260–5265. [Google Scholar] [CrossRef]
- Jaïdane, H.; Caloone, D.; Lobert, P.E.; Sané, F.; Dardenne, O.; Naquet, P.; Gharbi, J.; Aouni, M.; Geenen, V.; Hober, D. Persistent infection of thymic epithelial cells with coxsackievirus B4 results in decreased expression of type 2 insulin-like growth factor. J. Virol. 2012, 86, 11151–11162. [Google Scholar] [CrossRef]
- Brilot, F.; Geenen, V.; Hober, D.; Stoddart, C.A. Coxsackievirus B4 infection of human fetal thymus cells. J. Virol. 2004, 78, 9854–9861. [Google Scholar] [CrossRef][Green Version]
- Brilot, F.; Jaïdane, H.; Geenen, V.; Hober, D. Coxsackievirus B4 infection of murine foetal thymus organ cultures. J. Med. Virol. 2008, 80, 659–666. [Google Scholar] [CrossRef]
- Jaïdane, H.; Halouani, A.; Jmii, H.; Elmastour, F.; Abdelkefi, S.; Bodart, G.; Michaux, H.; Chakroun, T.; Sane, F.; Mokni, M.; et al. In-utero coxsackievirus B4 infection of the mouse thymus. Clin. Exp. Immunol. 2017, 187, 399–407. [Google Scholar] [CrossRef]
- Iwasaki, T.; Monma, N.; Satodate, R.; Kawana, R.; Kueata, T. An immunofluorescent study of generalized Coxsackievirus B3 infection in a newborn infant. Pathol. Int. 1985, 35, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Lozovskaia, L.S.; Osipov, S.M.; Zubkova, I.V.; Soboleva, V.D. Study of vertical transmission of coxsackie group enteroviruses in the etiology of congenital immunodeficiencies. Vopr. Virusol. 1997, 42, 175–179. [Google Scholar] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Halouani, A.; Jmii, H.; Michaux, H.; Renard, C.; Martens, H.; Pirottin, D.; Mastouri, M.; Aouni, M.; Geenen, V.; Jaïdane, H. Housekeeping Gene Expression in the Fetal and Neonatal Murine Thymus Following Coxsackievirus B4 Infection. Genes 2020, 11, 279. [Google Scholar] [CrossRef]
- Stoeckle, C.; Rota, I.A.; Tolosa, E.; Haller, C.; Melms, A.; Adamopoulou, E. Isolation of myeloid dendritic cells and epithelial cells from human thymus. JoVE 2013, 79, e50951. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Dotti, I.; Bonin, S. DNase treatment of RNA. In Guidelines for Molecular Analysis in Archive Tissues; Springer: Berlin/Heidelberg, Germany, 2011; pp. 87–90. [Google Scholar]
- Leparc, I.; Aymard, M.; Fuchs, F. Acute, chronic and persistent enterovirus and poliovirus infections, detection of viral genome by semi-nested PCR amplification in culture-negative samples. Mol. Cell. Probes 1994, 8, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Zoll, G.J.; Melchers, W.J.; Kopecka, H.; Jambroes, G.; van der Poel, H.J.; Galama, J.M. General primer-mediated polymerase chain reaction for detection of enteroviruses, application for diagnostic routine and persistent infections. J. Clin. Microbiol. 1992, 30, 160–165. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jaïdane, H.; Halouani, A.; Jmii, H.; Elmastour, F.; Mokni, M.; Aouni, M. Coxsackievirus B4 vertical transmission in a murine model. Virol. J. 2017, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Michaux, H.; Halouani, A.; Trussart, C.; Renard, C.; Jaïdane, H.; Martens, H.; Geenen, V.; Hober, D. Modulation of IGF2 Expression in the Murine Thymus and Thymic Epithelial Cells Following Coxsackievirus-B4 Infection. Microorganisms 2021, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef]
- Kyewski, B.; Klein, L. A central role for central tolerance. Annu. Rev. Immunol. 2006, 24, 571–606. [Google Scholar] [CrossRef]
- Kermani, H.; Goffinet, L.; Mottet, M.; Bodart, G.; Morrhaye, G.; Dardenne, O.; Renard, C.; Overbergh, L.; Baron, F.; Beguin, Y.; et al. Expression of the growth hormone/insulin-like growth factor axis during Balb/c thymus ontogeny and effects of growth hormone upon ex vivo T cell differentiation. Neuroimmunomodulation 2012, 19, 137–147. [Google Scholar] [CrossRef]
- Yang, S.; Fujikado, N.; Kolodin, D.; Benoist, C.; Mathis, D. Regulatory T cells generated early in life play a distinct role in maintaining self-tolerance. Science 2015, 348, 589–594. [Google Scholar] [CrossRef]
- Mathis, D.; Benoist, C. Aire. Annu. Rev. Immunol. 2009, 27, 287–312. [Google Scholar] [CrossRef] [PubMed]
- Takaba, H.; Morishita, Y.; Tomofuji, Y.; Danks, L.; Nitta, T.; Komatsu, N.; Kodama, T.; Takayanagi, H. Fezf2 orchestrates a thymic program of self-antigen expression for immune tolerance. Cell 2015, 163, 975–987. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.S.; Venanzi, E.S.; Klein, L.; Chen, Z.; Berzins, S.P.; Turley, S.J.; Von Boehmer, H.; Bronson, R.; Dierich, A.; Benoist, C.; et al. Projection of an immunological self shadow within the thymus by the aire protein. Science 2002, 298, 1395–1401. [Google Scholar] [CrossRef]
- Akirav, E.M.; Ruddle, N.H.; Herold, K.C. The role of AIRE in human autoimmune disease. Nat. Rev. Endocrinol. 2011, 7, 25. [Google Scholar] [CrossRef]
- Sanders, S.J.; Murtha, M.T.; Gupta, A.R.; Murdoch, J.D.; Raubeson, M.J.; Willsey, A.J.; Ercan-Sencicek, A.G.; DiLullo, N.M.; Parikshak, N.N.; Stein, J.L.; et al. De novo mutations revealed by whole-exome sequencing are strongly associated with autism. Nature 2012, 485, 237–241. [Google Scholar] [CrossRef]
- Kwan, K.Y. Transcriptional dysregulation of neocortical circuit assembly in ASD. Int. Rev. Neurobiol. 2013, 113, 167–205. [Google Scholar]
- Shu, X.S.; Li, L.; Ji, M.; Cheng, Y.; Ying, J.; Fan, Y.; Zhong, L.; Liu, X.; Tsao, S.W.; Chan, A.T.; et al. FEZF2, a novel 3p14 tumor suppressor gene, represses oncogene EZH2 and MDM2 expression and is frequently methylated in nasopharyngeal carcinoma. Carcinogenesis 2013, 34, 1984–1993. [Google Scholar] [CrossRef]
- Geenen, V.; Achour, I.; Robert, F.; Vandersmissen, E.; Sodoyez, J.C.; Defresne, M.P.; Boniver, J.; Lefebvre, P.; Franchimont, P. Evidence that insulin-like growth factor 2 (IGF2), is the dominant thymic peptide of the insulin superfamily. Thymus 1993, 21, 115–127. [Google Scholar] [PubMed]
- Kecha-Kamoun, O.; Achour, I.; Martens, H.; Collette, J.; Lefebvre, P.J.; Greiner, D.L.; Geenen, V. Thymic expression of insulin-related genes in an animal model of autoimmune type 1 diabetes. Diabetes Metab. Res. Rev. 2001, 17, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Michaux, H.; Martens, H.; Jaïdane, H.; Halouani, A.; Hober, D.; Geenen, V. How does thymus infection by coxsackievirus contribute to the pathogenesis of type 1 diabetes? Front. Immunol. 2015, 6, 338. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Havari, E.; Pinto, S.; Gottumukkala, R.V.; Cornivelli, L.; Raddassi, K.; Matsui, T.; Rosenzweig, A.; Bronson, R.T.; Smith, R.; et al. Impaired thymic tolerance to α-myosin directs autoimmunity to the heart in mice and humans. J. Clin. Investig. 2011, 121, 1561–1573. [Google Scholar] [CrossRef]
- Gotter, J.; Brors, B.; Hergenhahn, M.; Kyewski, B. Medullary epithelial cells of the human thymus express a highly diverse selection of tissue-specific genes colocalized in chromosomal clusters. J. Exp. Med. 2004, 199, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Derbinski, J.; Schulte, A.; Kyewski, B.; Klein, L. Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self. Nat. Immunol. 2001, 2, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.A.; Carthy, C.M.; Yang, D.; Saad, K.; Wong, D.; Schreiner, G.; Stanton, L.W.; McManus, B.M. Host gene regulation during coxsackievirus B3 infection in mice, assessment by microarrays. Circ. Res. 2000, 87, 328–334. [Google Scholar] [CrossRef]
- Ylipaasto, P.; Kutlu, B.; Rasilainen, S.; Rasschaert, J.; Salmela, K.; Teerijoki, H.; Korsgren, O.; Lahesmaa, R.; Hovi, T.; Eizirik, D.L.; et al. Global profiling of coxsackievirus-and cytokine-induced gene expression in human pancreatic islets. Diabetologia 2005, 48, 1510–1522. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Seah, J.B.K.; Chow, V.T.K.; Poh, C.L.; Tan, E.L. Comparative proteome analyses of host protein expression in response to Enterovirus 71 and Coxsackievirus A16 infections. J. Proteom. 2011, 74, 2018–2024. [Google Scholar] [CrossRef]
- Lepesant, H.; Pierres, M.; Naquet, P. Deficient antigen presentation by thymic epithelial cells reveals differential induction of T cell clone effector functions by CD28-mediated costimulation. Cell. Immunol. 1995, 161, 279–287. [Google Scholar] [CrossRef]
- Ucar, O.; Rattay, K. Promiscuous gene expression in the thymus, a matter of epigenetics, miRNA, and more? Front. Immunol. 2015, 6, 93. [Google Scholar] [CrossRef] [PubMed]
- Derbinski, J.; Gäbler, J.; Brors, B.; Tierling, S.; Jonnakuty, S.; Hergenhahn, M.; Peltonen, L.; Walter, J.; Kyewski, B. Promiscuous gene expression in thymic epithelial cells is regulated at multiple levels. J. Exp. Med. 2005, 202, 33–45. [Google Scholar] [CrossRef]
- Tomofuji, Y.; Takaba, H.; Suzuki, H.I.; Benlaribi, R.; Martinez, C.D.P.; Abe, Y.; Morishita, Y.; Okamura, T.; Taguchi, A.; Kodama, T.; et al. Chd4 choreographs self-antigen expression for central immune tolerance. Nat. Immunol. 2020, 21, 892–901. [Google Scholar] [CrossRef]
- Akiyama, T.; Shinzawa, M.; Qin, J.; Akiyama, N. Regulations of gene expression in medullary thymic epithelial cells required for preventing the onset of autoimmune diseases. Front. Immunol. 2013, 4, 249. [Google Scholar] [CrossRef] [PubMed]
- Takaba, H.; Takayanagi, H. The mechanisms of T cell selection in the thymus. Trends Immunol. 2017, 38, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Tedford, E.; McConkey, G. Neurophysiological changes induced by chronic Toxoplasma gondii infection. Pathogens 2017, 6, 19. [Google Scholar] [CrossRef]
- Yang, K.; Puel, A.; Zhang, S.; Eidenschenk, C.; Ku, C.L.; Casrouge, A.; Picard, C.; Von Bernuth, H.; Senechal, B.; Plancoulaine, S. Human TLR-7-,-8-, and-9-mediated induction of IFN-α/β and-λ is IRAK-4 dependent and redundant for protective immunity to viruses. Immunity 2005, 23, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Der, S.D.; Zhou, A.; Williams, B.R.; Silverman, R.H. Identification of genes differentially regulated by interferon α, β, or γ using oligonucleotide arrays. Proc. Natl. Acad. Sci. USA 1998, 95, 15623–15628. [Google Scholar] [CrossRef] [PubMed]
- Vitale, G.; Van Koetsveld, P.M.; De Herder, W.W.; Van Der Wansem, K.; Janssen, J.A.; Colao, A.; Lombardi, G.; Lamberts, S.W.; Hofland, L.J. Effects of type I interferons on IGF-mediated autocrine/paracrine growth of human neuroendocrine tumor cells. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E559–E566. [Google Scholar] [CrossRef]
- Van Koetsveld, P.M.; Vitale, G.; Feelders, R.A.; Waaijers, M.; Sprij-Mooij, D.M.; De Krijger, R.R.; Speel, E.J.; Hofland, J.; Lamberts, S.W.; De Herder, W.W.; et al. Interferon-beta is a potent inhibitor of cell growth and cortisol production In Vitro and sensitizes human adrenocortical carcinoma cells to mitotane. Endocr.-Relat. Cancer 2013, 20, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Theofilopoulos, A.N.; Kono, D.H.; Baccala, R. The multiple pathways to autoimmunity. Nat. Immunol. 2017, 18, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Qi, Y.; Li, H.; Ge, Y.; Zhao, K.; Qi, X.; Guo, X.; Shi, Z.; Zhou, M.; Zhu, B.; et al. Serum microRNA expression profile distinguishes enterovirus 71 and coxsackievirus 16 infections in patients with hand-foot-and-mouth disease. PLoS ONE 2011, 6, e27071. [Google Scholar] [CrossRef]
- Passos, G.A.; Mendes-da-Cruz, D.A.; Oliveira, E.H. The Role of Aire, microRNAs and Cell–Cell Interactions on Thymic Architecture and Induction of Tolerance. Front. Immunol. 2015, 6, 615. [Google Scholar] [CrossRef]
- Halouani, A.; Jmii, H.; Bodart, G.; Michaux, H.; Renard, C.; Martens, H.; Aouni, M.; Hober, D.; Geenen, V.; Jaïdane, H. Assessment of Thymic Output Dynamics After in utero Infection of Mice with Coxsackievirus B4. Front. Immunol. 2020, 11, 481. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Gene ID | Amplicon Length (bp) |
|---|---|---|---|---|
| Oaz1 | 5’-GCCAATGAACGAGATCACTT-3′ | 5′-GCTGTTTAAGATGGTCAGGTGA-3′ | 18245 | 110 |
| Aire | 5′-GGTTCTGTTGGACTCTGCCCTG-3′ | 5′-TGTGCCACGACGGAGGTGAG-3′ | 11634 | 144 |
| Fezf2 | 5′-GTGGCTCCCACCTTTGTACATTCA-3′ | 5′-TCACGGTGACAGGCTGGGATTAAA-3′ | 54713 | 121 |
| Igf2 | 5′-GGGAGCTTGTGGACACGC-3′ | 5′-GCACTCTTCCACGATGCCA-3′ | 16002 | 107 |
| Myo7 | 5′-TGCAAAGGCTCCAGGTCTGAGG-3′ | 5′-GCCAACACCAACCTGTCCAAGT-3′ | 140781 | 203 |
| Car | 5′-GGTTTGAGCATCACTACACCCG-3′ | 5′-TTCAATGTCCAGTGGTCCCTGG-3′ | 13052 | 114 |
| Antibody | Host Species | Dilution | Clone | Reference | Supplier | Tagged Protein Size (kDa) |
|---|---|---|---|---|---|---|
| VINCULIN | Rabbit | 1/500 | 42H89L44 | AB-2532280 | Thermofisher Scientific | 124 |
| AIRE | Rabbit | 1/1000 | Polyclonal | PA5-24554 | Thermofisher Scientific | 57 |
| FEZF2 | Rabbit | 2/1000 | Polyclonal | A05051 | Thermofisher Scientific | 50 |
| IGF2 | Rabbit | 1/500 | OAAB07463 | OAAB07463 | BosterBio | 20 |
| MYO7 | Rabbit | 4/1000 | Polyclonal | PA1-936 | Aviva Systems Technology | 220 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halouani, A.; Michaux, H.; Jmii, H.; Trussart, C.; Chahbi, A.; Martens, H.; Renard, C.; Aouni, M.; Hober, D.; Geenen, V.; et al. Coxsackievirus B4 Transplacental Infection Severely Disturbs Central Tolerogenic Mechanisms in the Fetal Thymus. Microorganisms 2021, 9, 1537. https://doi.org/10.3390/microorganisms9071537
Halouani A, Michaux H, Jmii H, Trussart C, Chahbi A, Martens H, Renard C, Aouni M, Hober D, Geenen V, et al. Coxsackievirus B4 Transplacental Infection Severely Disturbs Central Tolerogenic Mechanisms in the Fetal Thymus. Microorganisms. 2021; 9(7):1537. https://doi.org/10.3390/microorganisms9071537
Chicago/Turabian StyleHalouani, Aymen, Hélène Michaux, Habib Jmii, Charlotte Trussart, Ahlem Chahbi, Henri Martens, Chantal Renard, Mahjoub Aouni, Didier Hober, Vincent Geenen, and et al. 2021. "Coxsackievirus B4 Transplacental Infection Severely Disturbs Central Tolerogenic Mechanisms in the Fetal Thymus" Microorganisms 9, no. 7: 1537. https://doi.org/10.3390/microorganisms9071537
APA StyleHalouani, A., Michaux, H., Jmii, H., Trussart, C., Chahbi, A., Martens, H., Renard, C., Aouni, M., Hober, D., Geenen, V., & Jaïdane, H. (2021). Coxsackievirus B4 Transplacental Infection Severely Disturbs Central Tolerogenic Mechanisms in the Fetal Thymus. Microorganisms, 9(7), 1537. https://doi.org/10.3390/microorganisms9071537