Effects of Dietary Fiber Sources during Gestation on Stress Status, Abnormal Behaviors and Reproductive Performance of Sows

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets and Housing
2.2. Sample Collection
2.3. Performance Measurement
2.4. In Vitro Fermentation Analysis
2.5. Chemical Analysis and Calculation
2.6. Behavior Determination
2.7. Statistical Analysis
3. Results
3.1. Physical Properties of Gestation Diets and In Vitro Fermentability of Gestation Diets
3.2. Sow Performance
3.3. Piglet Performance
3.4. Behavior Determination
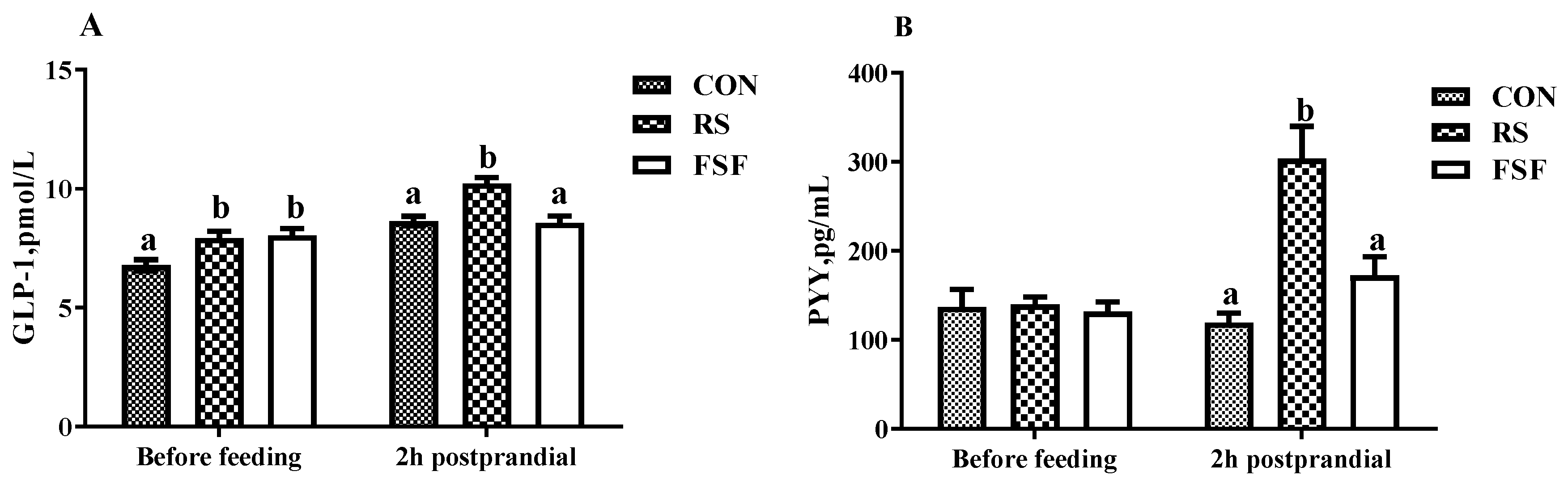
3.5. PYY and GLP-1 Concentration of Serum
3.6. Cortisol Concentration in Plasma and Saliva
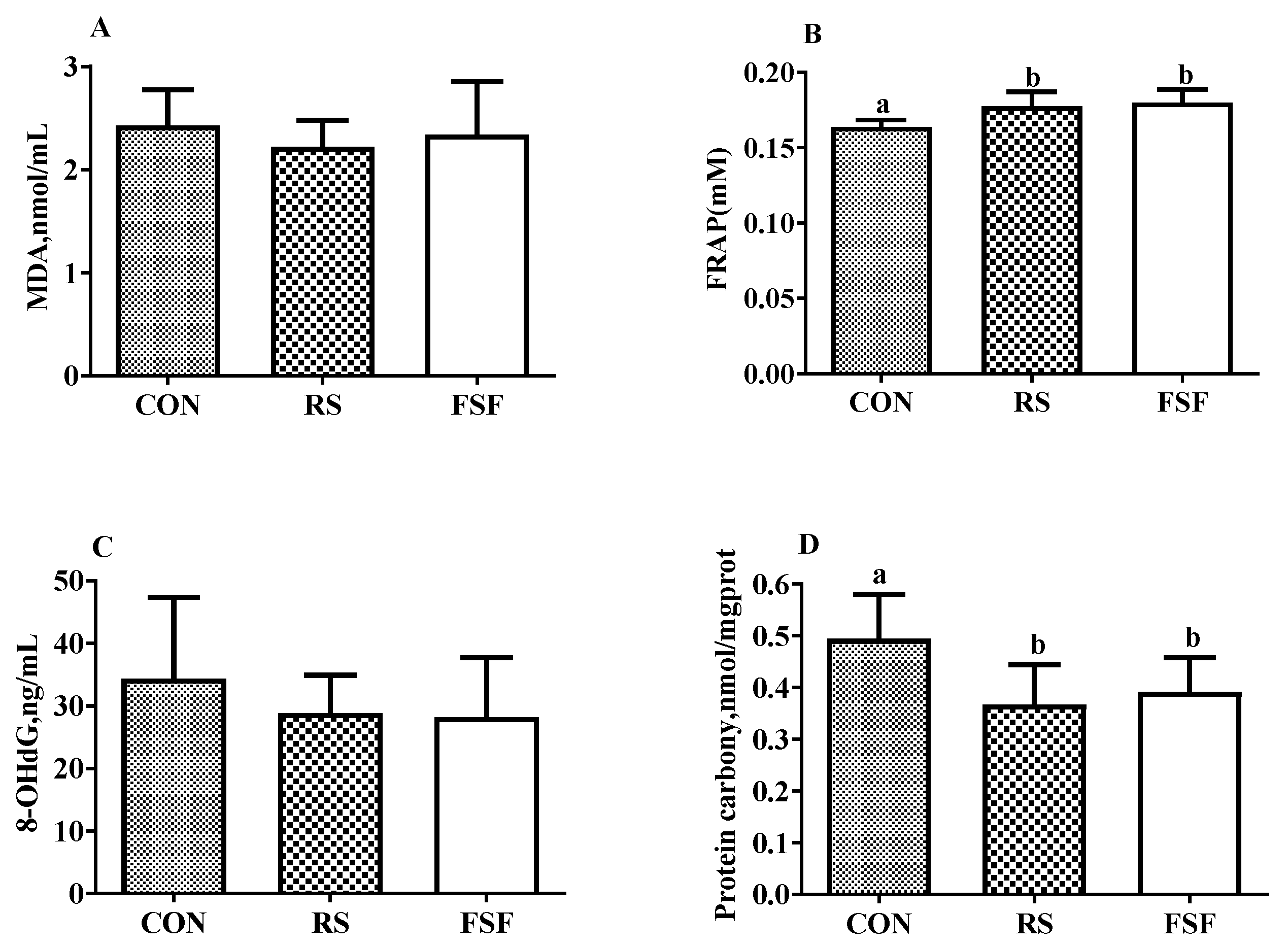
3.7. Oxidative and Antioxidative Indicators
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meunier-Salaün, M.; Edwards, S.; Robert, S. Effect of dietary fibre on the behaviour and health of the restricted fed sow. Anim. Feed Sci. Technol. 2001, 90, 53–69. [Google Scholar]
- Robert, S.; Bergeron, R.; Farmer, C.; Meunier-Salaün, M.C. Does the number of daily meals affect feeding motivation and behaviour of gilts fed high-fibre diets? Appl. Anim. Behav. Sci. 2002, 76, 105–117. [Google Scholar] [CrossRef]
- Renteria-Flores, J.; Johnston, L.; Shurson, G.C.; Moser, R.; Webel, S. Effect of soluble and insoluble dietary fiber on embryo survival and sow performance. J. Anim. Sci. 2008, 86, 2576–2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekiguchi, T.; Koketsu, Y. Behavior and reproductive performance by stalled breeding females on a commercial swine farm. J. Anim. Sci. 2004, 82, 1482–1487. [Google Scholar] [CrossRef] [Green Version]
- Quesnel, H.; Meunier-Salaun, M.-C.; Hamard, A.; Guillemet, R.; Etienne, M.; Farmer, C.; Dourmad, J.-Y.; Père, M.-C. Dietary fiber for pregnant sows: Influence on sow physiology and performance during lactation. J. Anim. Sci. 2009, 87, 532–543. [Google Scholar] [CrossRef] [Green Version]
- Reese, D.; Prosch, A.; Travnicek, D.A.; Eskridge, K.M. Dietary fiber in sow gestation diets-an updated review. Rep. Nebr. Swine 2008, 45, 14–18. [Google Scholar]
- Matte, J.J.; Robert, S.; Girard, C.L.; Farmer, C.; Martineau, G.P. Effect of bulky diets based on wheat bran or oat hulls on reproductive performance of sows during their first two parities. J. Anim. Sci. 1994, 72, 1754–1760. [Google Scholar] [CrossRef]
- Holt, J.P.; Johnston, L.J.; Baidoo, S.K.; Shurson, G.C. Effects of a high-fiber diet and frequent feeding on behavior, reproductive performance, and nutrient digestibility in gestating sows. J. Anim. Sci. 2006, 84, 946–955. [Google Scholar] [CrossRef]
- Holzgraefe, D.; Jensen, A.; Fahey, G., Jr.; Grummer, R. Effects of dietary alfalfa-orchardgrass hay and lasalocid on sow reproductive performance. J. Anim. Sci. 1986, 62, 1145–1153. [Google Scholar] [CrossRef]
- Sun, H.; Zhou, Y.; Tan, C.; Zheng, L.; Peng, J.; Jiang, S. Effects of konjac flour inclusion in gestation diets on the nutrient digestibility, lactation feed intake and reproductive performance of sows. Animal 2014, 8, 1089–1094. [Google Scholar] [CrossRef]
- Cummings, J.H.; Englyst, H.N. Measurement of starch fermentation in the human large intestine. Can. J. Physiol. Pharmacol. 1991, 69, 121–129. [Google Scholar] [CrossRef]
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 46, S33–S50. [Google Scholar]
- Giuberti, G.; Gallo, A.; Moschini, M.; Masoero, F. New insight into the role of resistant starch in pig nutrition. Anim. Feed Sci. Technol. 2015, 201, 1–13. [Google Scholar] [CrossRef]
- Sajilata, M.G.; Singhal, R.S.; Kulkarni, P.R. Resistant starch–a review. Compr. Rev. Food Sci. Food Saf. 2006, 5, 1–17. [Google Scholar] [CrossRef]
- Asp, N.G. Nutritional classification and analysis of food carbohydrates. Am. J. Clin. Nutr. 1994, 59, 679S–681S. [Google Scholar] [CrossRef]
- Eerlingen, R.C.; Delcour, J.A. Formation, analysis, structure and properties of Type III enzyme resistant starch. J. Cereal Sci. 1995, 22, 129–138. [Google Scholar] [CrossRef]
- Sapkota, A.; Marchant-Forde, J.; Richert, B.; Lay, D., Jr. Including dietary fiber and resistant starch to increase satiety and reduce aggression in gestating sows. J. Anim. Sci. 2016, 94, 2117–2127. [Google Scholar] [CrossRef]
- Kim, J.H.; Jia, Y.; Lee, J.G.; Nam, B.; Lee, S.J. Hypolipidemic and antiinflammation activities of fermented soybean fibers from meju in C57BL/6 J mice. Phytother. Res. 2014, 28, 1335–1341. [Google Scholar] [CrossRef]
- USA, National Research Council. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Mochida, T.; Take, K.; Maki, T.; Nakakariya, M.; Adachi, R.; Sato, K.; Kitazaki, T.; Takekawa, S. Inhibition of MGAT2 modulates fat-induced gut peptide release and fat intake in normal mice and ameliorates obesity and diabetes in ob/ob mice fed on a high fat diet. FEBS Open Biol. 2019. [Google Scholar] [CrossRef]
- Gibbons, C.; Caudwell, P.; Finlayson, G.; Webb, D.-L.; Hellström, P.M.; Näslund, E.; Blundell, J.E. Comparison of postprandial profiles of ghrelin, active GLP-1, and total PYY to meals varying in fat and carbohydrate and their association with hunger and the phases of satiety. J. Clin. Endocrinol. Metab. 2013, 98, E847–E855. [Google Scholar] [CrossRef] [Green Version]
- Kiros, T.K.; Agyekum, A.K.; Wang, J.; D’inca, R.; Beaulieu, D.A.; Auclair, E.; Van Kessel, A.G. Supplementation with live yeast increases rate and extent of in vitro fermentation of nondigested feed ingredients by fecal microbiota. J. Anim. Sci. 2019, 97, 1806–1818. [Google Scholar] [CrossRef] [PubMed]
- Menke, K.H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Bindelle, J.; Buldgen, A.; Boudry, C.; Leterme, P. Effect of inoculum and pepsin–pancreatin hydrolysis on fibre fermentation measured by the gas production technique in pigs. Anim. Feed Sci. Technol. 2007, 132, 111–122. [Google Scholar] [CrossRef]
- Crittenden, R.; Karppinen, S.; Ojanen, S.; Tenkanen, M.; Fagerström, R.; Mättö, J.; Saarela, M.; Mattila-Sandholm, T.; Poutanen, K. In vitro fermentation of cereal dietary fibre carbohydrates by probiotic and intestinal bacteria. J. Sci. Food Agric. 2002, 82, 781–789. [Google Scholar] [CrossRef]
- Jha, R.; Bindelle, J.; Van Kessel, A.; Leterme, P. In vitro fibre fermentation of feed ingredients with varying fermentable carbohydrate and protein levels and protein synthesis by colonic bacteria isolated from pigs. Anim. Feed Sci. Technol. 2011, 165, 191–200. [Google Scholar] [CrossRef]
- Van, P.Y.; Hamilton, G.J.; Kremenevskiy, I.V.; Sambasivan, C.; Spoerke, N.J.; Differding, J.A.; Watters, J.M.; Schreiber, M.A. Lyophilized plasma reconstituted with ascorbic acid suppresses inflammation and oxidative DNA damage. J. Trauma Acute Care Surg. 2011, 71, 20–25. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Grotto, D.; Santa Maria, L.; Boeira, S.; Valentini, J.; Charão, M.; Moro, A.; Nascimento, P.; Pomblum, V.; Garcia, S. Rapid quantification of malondialdehyde in plasma by high performance liquid chromatography–visible detection. J. Pharm. Biomed. Anal. 2007, 43, 619–624. [Google Scholar] [CrossRef]
- Ganhão, R.; Morcuende, D.; Estévez, M. Protein oxidation in emulsified cooked burger patties with added fruit extracts: Influence on colour and texture deterioration during chill storage. Meat Sci. 2010, 85, 402–409. [Google Scholar] [CrossRef]
- Tan, C.; Sun, H.; Wei, H.; Tan, J.; Long, G.; Jiang, S.; Peng, J. Effects of soluble fiber inclusion in gestation diets with varying fermentation characteristics on lactational feed intake of sows over two successive parities. Animal 2018, 12, 1388–1395. [Google Scholar] [CrossRef]
- De Leeuw, J.; Bolhuis, J.; Bosch, G.; Gerrits, W. Effects of dietary fibre on behaviour and satiety in pigs: Symposium on ‘Behavioural nutrition and energy balance in the young’. Proc. Nutr. Soc. 2008, 67, 334–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchant-Forde, R.; Marchant-Forde, J. Pregnancy-related changes in behavior and cardiac activity in primiparous pigs. Physiol. Behav. 2004, 82, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Wenk, C. The role of dietary fibre in the digestive physiology of the pig. Anim. Feed Sci. Technol. 2001, 90, 21–33. [Google Scholar] [CrossRef]
- Howarth, N.C.; Saltzman, E.; Roberts, S.B. Dietary Fiber and Weight Regulation. Nutr. Rev. 2001, 59, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Zijlstra, N.; De Wijk, R.A.; Mars, M.; Stafleu, A.; De, G.C. Effect of bite size and oral processing time of a semisolid food on satiation. Am. J. Clin. Nutr. 2008, 51, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schueler, J.L.; Alexander, B.M.; Hart, A.M.; Austin, K.J.; Larson-Meyer, D.E. Presence and Dynamics of Leptin, GLP-1, and PYY in Human Breast Milk at Early Postpartum. Obesity 2013, 21, 1451–1458. [Google Scholar] [CrossRef]
- Blundell, J.E.; Naslund, E. Glucagon-like peptide-1, satiety and appetite control. Br. J. Nutr. 1999, 81, 259–260. [Google Scholar] [CrossRef] [Green Version]
- Sleeth, M.L.; Thompson, E.L.; Ford, H.E.; Zac-Varghese, S.E.; Frost, G. Free fatty acid receptor 2 and nutrient sensing: A proposed role for fibre, fermentable carbohydrates and short-chain fatty acids in appetite regulation. Nutr. Rev. 2010, 23, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Keenan, M.J.; Raggio, A.M.; Tripathy, S.; Shen, L.; Mccutcheon, K.L.; Hegsted, M.; Tulley, R.T.; Martin, R.J. Feeding resistant starch maintains elevated plasma levels of GLP-1 and PYY throughout the day and is associated with decreased body fat in rats. FASEB J. 2007, 21, A158. [Google Scholar]
- Zhou, J.; Martin, R.J.; Tulley, R.T.; Raggio, A.M.; Mccutcheon, K.L.; Shen, L.; Danna, S.C.; Tripathy, S.; Hegsted, M.; Keenan, M.J. Dietary resistant starch upregulates total GLP-1 and PYY in a sustained day-long manner through fermentation in rodents. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1160–E1166. [Google Scholar] [CrossRef] [Green Version]
- Byrne, C.; Chambers, E.; Morrison, D.; Frost, G. The role of short chain fatty acids in appetite regulation and energy homeostasis. Int. J. Obes. 2015, 39, 1331–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coles, L.; Moughan, P.; Darragh, A. In vitro digestion and fermentation methods, including gas production techniques, as applied to nutritive evaluation of foods in the hindgut of humans and other simple-stomached animals. Anim. Feed Sci. Technol. 2005, 123, 421–444. [Google Scholar] [CrossRef]
- Bindelle, J.; Buldgen, A.; Lambotte, D.; Wavreille, J.; Leterme, P. Effect of pig faecal donor and of pig diet composition on in vitro fermentation of sugar beet pulp. Anim. Feed Sci. Technol. 2007, 132, 212–226. [Google Scholar] [CrossRef]
- Farmer, C.; Lapointe, J.; Palin, M.-F. Effects of the plant extract silymarin on prolactin concentrations, mammary gland development, and oxidative stress in gestating gilts. J. Anim. Sci. 2014, 92, 2922–2930. [Google Scholar] [CrossRef]
- Tan, C.; Hongkui, W.; Haiqing, S.; Jiangtao, A.; Guang, L.; Siwen, J.; Jian, P. Effects of Dietary Supplementation of Oregano Essential Oil to Sows on Oxidative Stress Status, Lactation Feed Intake of Sows, and Piglet Performance. Biomed. Res. Int. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tao, S.; Bubolz, J.; Do Amaral, B.; Thompson, I.; Hayen, M.; Johnson, S.; Dahl, G. Effect of heat stress during the dry period on mammary gland development. J. Dairy Sci. 2011, 94, 5976–5986. [Google Scholar] [CrossRef]
- Kim, S.W.; Weaver, A.C.; Shen, Y.B.; Zhao, Y. Improving efficiency of sow productivity: Nutrition and health. J. Anim. Sci. Biotechnol. 2013, 4, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Tan, C.; Wei, H.; Zou, Y.; Long, G.; Ao, J.; Xue, H.; Jiang, S.; Peng, J. Effects of different amounts of konjac flour inclusion in gestation diets on physio-chemical properties of diets, postprandial satiety in pregnant sows, lactation feed intake of sows and piglet performance. Anim. Reprod. Sci. 2015, 152, 55–64. [Google Scholar] [CrossRef]
- Jensen, E.; Gallaher, B.; Breier, B.; Harding, J. The effect of a chronic maternal cortisol infusion on the late-gestation fetal sheep. J. Endocrinol. 2002, 174, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Renteria-Flores, J.A.; Johnston, L.J.; Shurson, G.C.; Gallaher, D.D. Effect of soluble and insoluble fiber on energy digestibility, nitrogen retention, and fiber digestibility of diets fed to gestating sows. J. Anim. Sci. 2008, 86, 2568–2575. [Google Scholar] [CrossRef]
- Grieshop, C.M.; Reese, D.E.; Fahey, G.C., Jr. Nonstarch polysaccharides and oligosaccharides in swine nutrition. In Swine Nutrition, 2nd ed.; Lewis, A.J., Southern, L.L., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 107–131. [Google Scholar]
- Ewan, R.; Crenshaw, J.; Crenshaw, T.; Cromwell, G.; Easter, R.; Nelssen, J.; Miller, E.; Pettigrew, J.; Veum, T. Effect of addition of fiber to gestation diets on reproductive performance of sows. J. Anim. Sci. 1996, 74, 190. [Google Scholar]
- Allee, G.L. Alfalfa haylage for sows during gestation. J. Anim. Sci. 1981, 55, 38–41. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Gestation Diet | Lactation Diet | ||
|---|---|---|---|---|
| CON 1 | RS 1 | FSF 1 | ||
| Ingredient, % | ||||
| Corn | 61.60 | 53.60 | 61.30 | 61.00 |
| Soybean meal | 11.00 | 12.00 | 9.20 | 19.50 |
| Wheat bran | 20.00 | 20.00 | 18.00 | - |
| Soybean hull | 3.00 | 5.00 | 2.00 | 5.00 |
| Wheat middlings | - | - | - | 1.50 |
| Corn meal | 0.35 | |||
| Fish meal, 67%CP | 1.00 | |||
| Extruded soybean | 5.50 | |||
| Soybean oil | 0.60 | |||
| Glucose | 1.25 | |||
| Resistant starch 2 | - | 5.00 | - | - |
| Fermented soybean fiber 3 | - | - | 5.00 | - |
| Dicalcium phosphate | 1.70 | 1.70 | 1.70 | 1.55 |
| Limestone | 1.00 | 1.00 | 1.00 | 0.90 |
| Salt | 0.50 | 0.50 | 0.50 | 0.50 |
| Lysine sulfate (70%) | 0.20 | 0.20 | 0.30 | 0.30 |
| Threonine | 0.10 | |||
| Methionine | 0.05 | |||
| Choline chloride | 0.10 | 0.10 | 0.10 | 0.10 |
| Sodium bicarbonate | 0.40 | 0.40 | 0.40 | 0.40 |
| Mildewcide | 0.10 | 0.10 | 0.10 | - |
| Premix 4 | 0.40 | 0.40 | 0.40 | 0.40 |
| Calculated compositon 5 | ||||
| DE (Mcal/kg) | 2.99 | 2.98 | 3.00 | 3.30 |
| CP (%) | 13.30 | 13.30 | 13.30 | 17.61 |
| EE (%) | 3.10 | 2.90 | 3.10 | 4.05 |
| CF (%) | 5.40 | 6.00 | 6.33 | 3.80 |
| NDF (%) | 17.00 | 17.80 | 17.90 | 12.26 |
| Ca (%) | 0.80 | 0.90 | 0.80 | 0.85 |
| Total phosphorus (%) | 0.70 | 0.70 | 0.70 | 0.64 |
| Lys (%) | 0.70 | 0.70 | 0.70 | 1.15 |
| Met (%) | 0.20 | 0.20 | 0.20 | 0.34 |
| Thr (%) | 0.50 | 0.50 | 0.50 | 0.78 |
| Trp (%) | 0.20 | 0.20 | 0.20 | 0.19 |
| Analyzed composition | ||||
| CF (%) | 5.34 | 5.69 | 6.30 | |
| NDF (%) | 19.00 | 19.15 | 19.50 | |
| Physio-chemical properties | ||||
| Viscosity (mPa·s) | 1.83 | 1.88 | 1.82 | - |
| Water-blinding capacity (g/g) | 0.09 | 0.09 | 0.10 | - |
| Swelling capacity (mL/g) | 1.63 | 2.28 | 1.89 | - |
| Item | Diet | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON 1 | RS 1 | FSF 1 | |||
| Fermented Fraction | |||||
| V 2 | 187.17 | 197.60 | 169.99 | 5.60 | 0.07 |
| VF 3 | 210.41 | 222.77 | 197.96 | 5.08 | 0.09 |
| (FRD0) 4 ×100 | 1.86 | 2.00 | 2.31 | 0.16 | 0.61 |
| K 5 | 0.10 | 0.08 | 0.06 | 0.01 | 0.65 |
| (T1/2) 6 | 20.75 | 20.52 | 20.93 | 0.24 | 0.86 |
| Concentrations of SCFA | |||||
| Acetic acid, mmol/L | 20.08 | 19.65 | 16.15 | 1.15 | 0.39 |
| Propionic acid, mmol/L | 9.17 | 8.57 | 6.67 | 0.64 | 0.30 |
| Butyric acid, mmol/L | 1.59 | 1.35 | 1.18 | 1.10 | 0.32 |
| Iso-butyric acid, mmol/L | 0.49 | 0.30 | 0.26 | 0.06 | 0.31 |
| Valeric acid, mmol/L | 1.50 | 1.09 | 0.97 | 0.13 | 0.22 |
| Iso-valeric acid, mmol/L | 0.83 | 0.46 | 0.40 | 0.12 | 0.33 |
| Total SCFA 7, mmol/L | 33.66 | 31.43 | 25.63 | 2.07 | 0.32 |
| Item | Diet | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON 1 | RS 1 | FSF 1 | |||
| No. of sows | 22 | 21 | 24 | ||
| Average sow parity | 5.1 | 4.9 | 4.9 | ||
| BW of sows, kg | |||||
| Mating | 229.2 | 228.9 | 229.9 | 2.89 | 0.99 |
| D109 of gestation 2 | 285.5 | 279.1 | 282.8 | 2.84 | 0.62 |
| Gain during gestation | 57.6 | 51.0 | 51.2 | 1.64 | 0.19 |
| Parturition 2 | 258.5 | 255.6 | 257.1 | 2.82 | 0.96 |
| Weaning | 243.9 | 243.8 | 240.6 | 2.97 | 0.83 |
| Loss during lactation 2 | 16.4 | 12.4 | 18.3 | 1.64 | 0.45 |
| Sow backfat thickness, mm | |||||
| Mating | 17.5 | 17.6 | 17.1 | 0.52 | 0.91 |
| D109 of gestation | 17.6 | 16.9 | 17.4 | 0.55 | 0.83 |
| Weaning 2 | 16.2 | 15.6 | 15.2 | 0.42 | 0.52 |
| Weaning to estrus interval 2, d | 4.2 | 4.3 | 4.8 | 0.13 | 0.21 |
| Average daily feed intake 3, kg | |||||
| 1st week of lactation | 5.3 | 5.6 | 5.5 | 0.14 | 0.89 |
| 2nd week of lactation 2 | 7.2 | 7.3 | 7.3 | 0.11 | 0.92 |
| 3rd week of lactation | 7.8 | 7.3 | 7.5 | 0.12 | 0.27 |
| Mean of 1st week to 3rd week | 6.8 | 6.7 | 6.7 | 0.09 | 0.98 |
| Item | Diet | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON 1 | RS 1 | FSF 1 | |||
| No. of sows | 19 | 21 | 22 | ||
| Average sow parity | 4.5 | 4.8 | 5.0 | ||
| No. of pigs per litter | |||||
| Total piglets born 2 | 15.6 | 14.0 | 15.0 | 0.38 | 0.24 |
| Piglets born alive 2,3 | 13.3 | 13.0 | 13.0 | 0.33 | 0.77 |
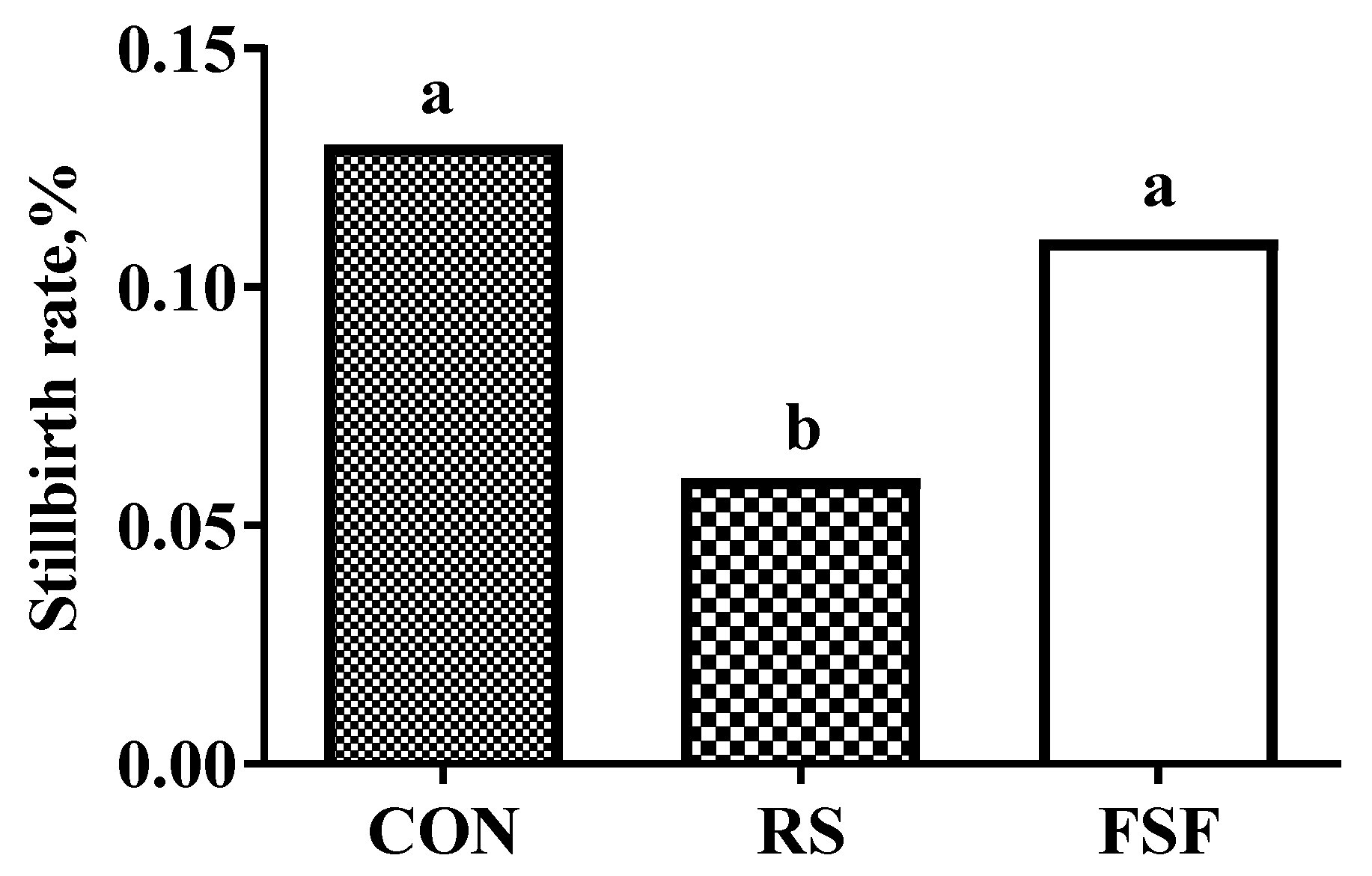
| Stillbirth 2,3 | 2.0 a | 0.9 b | 1.8 ab | 0.20 | 0.04 |
| After cross-foster 3 | 12.1 | 11.6 | 11.5 | 0.11 | 0.12 |
| Pigs weaned 3 | 10.6 | 10.0 | 10.3 | 0.18 | 0.20 |
| Piglet mean BW, kg | |||||
| At Birth 2,3 | 1.4 | 1.4 | 1.4 | 0.03 | 0.99 |
| After cross-foster 3 | 1.6 | 1.6 | 1.6 | 0.03 | 0.43 |
| On day 7 | 3.1 | 3.2 | 3.1 | 0.06 | 0.59 |
| On day 14 | 5.1 | 5.1 | 5.1 | 0.05 | 0.67 |
| On day 21 | 7.0 | 7.1 | 7.0 | 0.07 | 0.73 |
| Average daily gain, g/d | |||||
| Day 1 to 7 | 209.7 | 220.3 | 207.0 | 7.90 | 0.45 |
| Day 7 to 14 | 277.9 | 283.9 | 274.4 | 7.54 | 0.67 |
| Day 14 to 21 | 268.0 | 276.4 | 261.8 | 11.00 | 0.65 |
| Day 1 to 21 | 253.6 | 265.8 | 249.5 | 6.80 | 0.22 |
| Item | Diet | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON 1 | RS 1 | FSF 1 | |||
| No. of sows | 8 | 8 | 8 | ||
| Lying | 91.88 | 96.46 | 91.20 | 1.45 | 0.13 |
| Standing | 7.66 a | 1.77 b | 7.24 a | 1.31 | 0.05 |
| Sitting | 0.47 | 1.77 | 1.62 | 1.38 | 0.74 |
| Drinking, times | 2.00 | 1.25 | 1.63 | 0.31 | 0.45 |
| Sham-chewing | 13.07 | 4.11 | 14.32 | 3.41 | 0.24 |
| Sniffing, licking | 1.93 | 0.68 | 0.63 | 0.27 | 0.19 |
| Position change 2, times | 3.63 | 1.75 | 3.38 | 0.44 | 0.17 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.; Wei, J.; Yu, H.; Hao, X.; Zuo, J.; Tan, C.; Deng, J. Effects of Dietary Fiber Sources during Gestation on Stress Status, Abnormal Behaviors and Reproductive Performance of Sows. Animals 2020, 10, 141. https://doi.org/10.3390/ani10010141
Huang S, Wei J, Yu H, Hao X, Zuo J, Tan C, Deng J. Effects of Dietary Fiber Sources during Gestation on Stress Status, Abnormal Behaviors and Reproductive Performance of Sows. Animals. 2020; 10(1):141. https://doi.org/10.3390/ani10010141
Chicago/Turabian StyleHuang, Shuangbo, Jianfu Wei, Haoyuan Yu, Xiangyu Hao, Jianjun Zuo, Chengquan Tan, and Jinping Deng. 2020. "Effects of Dietary Fiber Sources during Gestation on Stress Status, Abnormal Behaviors and Reproductive Performance of Sows" Animals 10, no. 1: 141. https://doi.org/10.3390/ani10010141
APA StyleHuang, S., Wei, J., Yu, H., Hao, X., Zuo, J., Tan, C., & Deng, J. (2020). Effects of Dietary Fiber Sources during Gestation on Stress Status, Abnormal Behaviors and Reproductive Performance of Sows. Animals, 10(1), 141. https://doi.org/10.3390/ani10010141