A Novel Method for Increasing the Numerousness of Biometrical Parameters Useful for Wildlife Management: Roe Deer Mandible as Bone Model

 ,
,  , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
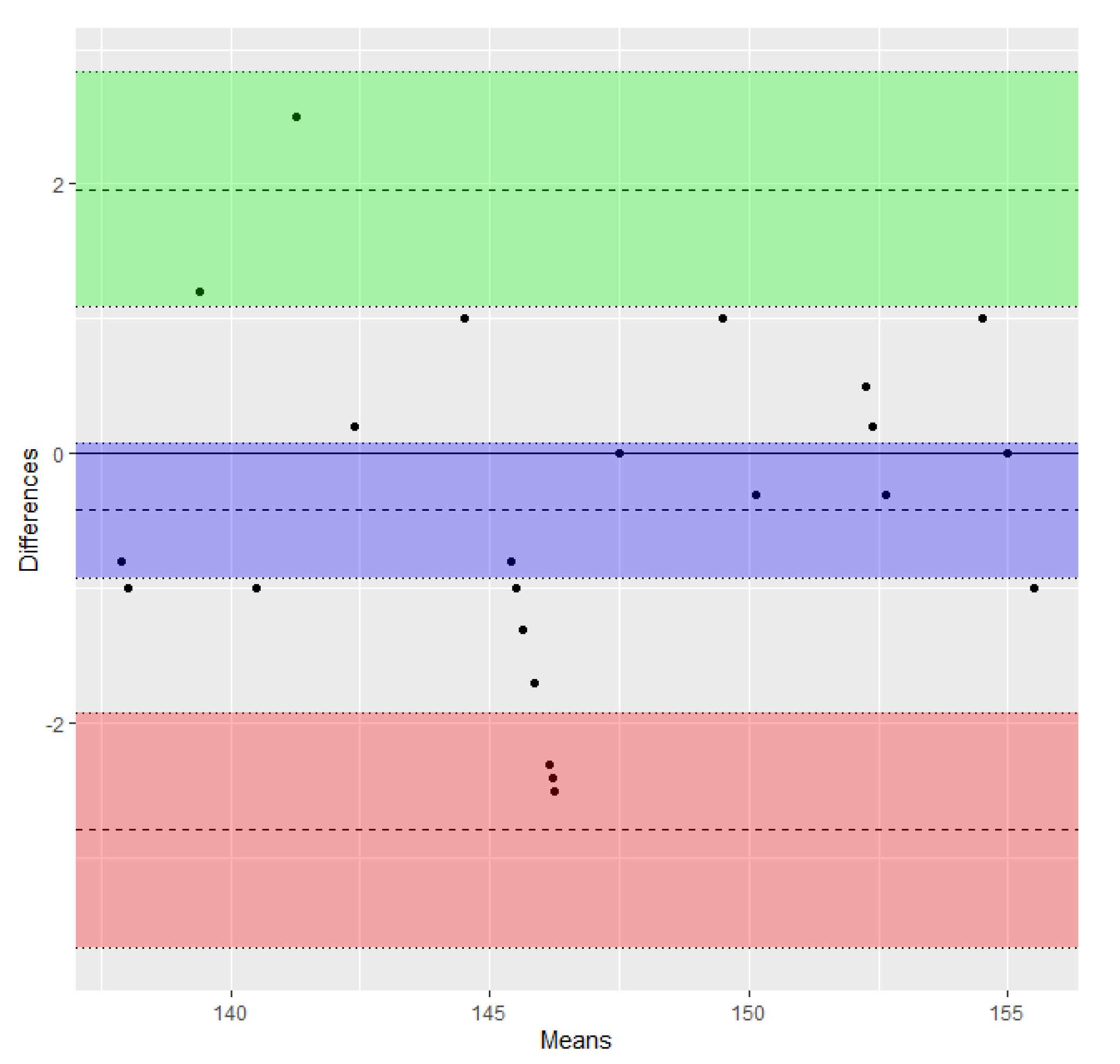
2.1. Method to Define the Conversion Factor
2.2. Conversion and Analysis of Additional Measures
- -
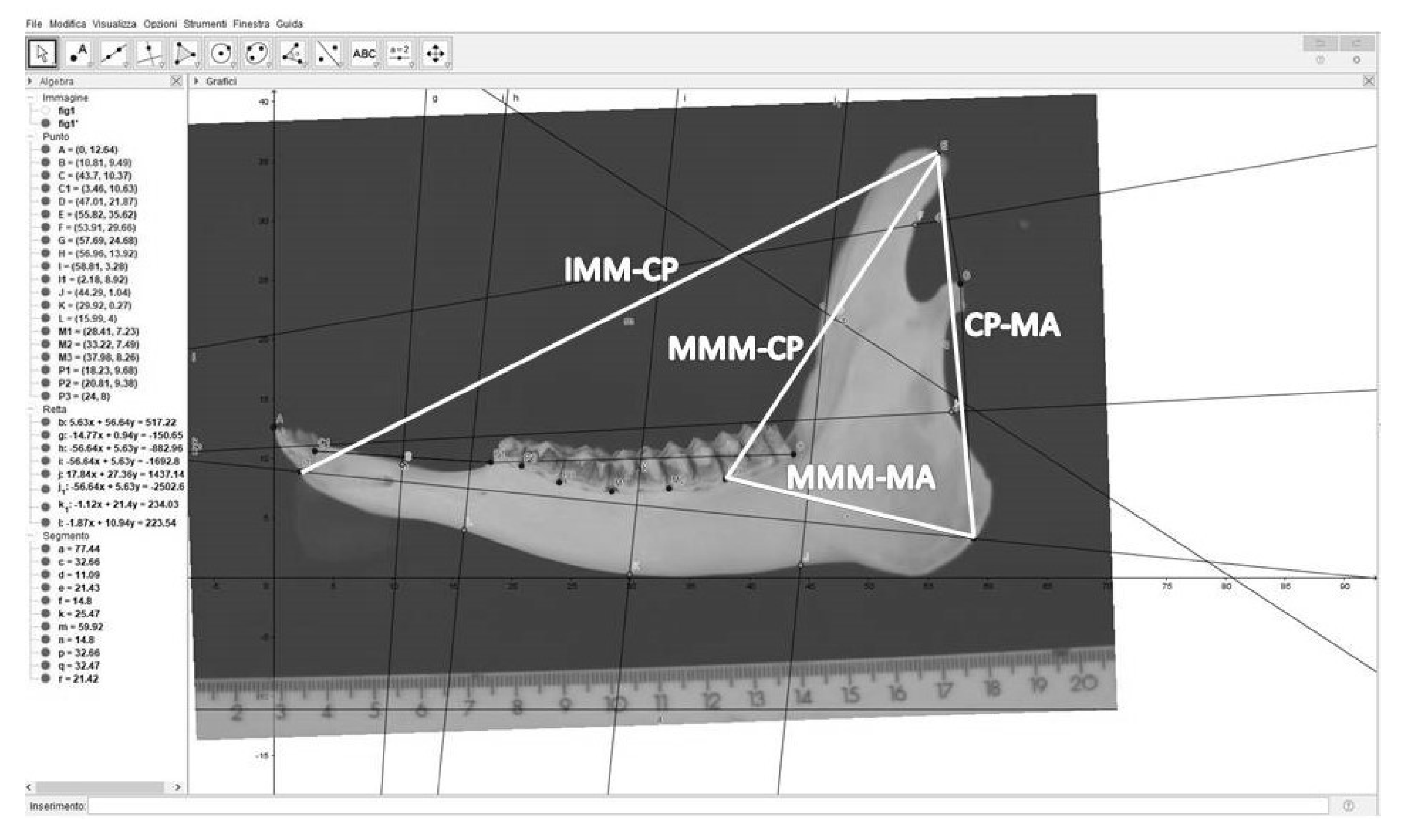
- distance from the mesial margin of the first incisor (IMM) to the coronoid process (CP), hereafter IMM-CP;
- -
- distance from the mesial margin of the 3rd molar tooth (MMM) to the coronoid process (CP), hereafter MMM-CP;
- -
- distance from the coronoid process (CP) to the angle of the mandible (MA), hereafter CP-MA;
- -
- distance from the mesial margin of the 3rd molar tooth (MMM) to the angle of the mandible (MA), hereafter MMM-MA.
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanzal, V.; Janiszewski, P.; Tajchman, K.; Košinová, K. The correlation between mandibular length versus body mass and age in the european roe deer (Capreolus capreolus L.). Appl. Ecol. Env. Res. 2017, 15, 1623–1632. [Google Scholar] [CrossRef]
- Becciolini, V.; Bozzi, R.; Viliani, M.; Biffani, S.; Ponzetta, M.P. Body measurements from selective hunting: Biometric features of red deer (Cervus elaphus) from Northern Apennine, Italy. Ital. J. Anim. Sci. 2016, 15, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Moretti, M. Biometric data and growth rates of a mountain population of wild boar (Sus scrofa L.), Ticino, Switzerland. J. Mountain Ecol. 2014, 3, 56–59. [Google Scholar]
- Zannèse, A.; Baïsse, A.; Gaillard, J.M.; Hewison, A.J.M.; Saint-Hilaire, K.; Toïgo, C.; VanLaere, G.; Morellet, N. Hind foot length: A new biological indicator for monitoring roe deer populations at a landscape scale. Wildlife Soc. B 2006, 34, 351–358. [Google Scholar] [CrossRef]
- Blant, M.; Gaillard, J.M. Use of biometric body variables as indicators of roe deer (Capreolus capreolus) population density changes. Game Wildlife Sci. 2004, 21, 21–40. [Google Scholar]
- Aragon, S.; Braza, F.; Jose, C.S.; Fandos, P. Variation in skull morphology of roe deer (Capreolus capreolus) in western and central Europe. J. Mammal. 1998, 79, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Carnevali, L.; Pedrotti, L.; Riga, F.; Toso, S. Banca dati ungulati: Status, distribuzione, consistenza, gestione e prelievo venatorio delle popolazioni di ungulati in Italia: Rapporto 2001–2005. Biol. Cons. Fauna. 2009, 117, 1–168. [Google Scholar]
- Mattioli, S.; De Marinis, A.M.; Armaroli, E. Guida al rilevamento biometrico degli Ungulati. ISPRA Documenti Tecnici 2009, 28, 1–216. [Google Scholar]
- Horcajada-Sánchez, F.; Barja, I. Local ecotypes of roe deer populations (Capreolus capreolus L.) in relation to morphometric features and fur colouration in the centre of the Iberian Peninsula. Pol. J. Ecol. 2016, 64, 113–124. [Google Scholar] [CrossRef]
- Toïgo, C.; Gaillard, J.M.; Van Laere, G.; Hewison, M.; Morellet, N. How does environmental variation influence body mass, body size, and body condition? Roe deer as a case study. Ecography 2006, 29, 301–308. [Google Scholar]
- Pettorelli, N.; Gaillard, J.M.; van Laere, G.; Duncan, P.; Kjellander, P.; Liberg, O.; Delorme, D.; Maillard, D. Variations in adult body mass in roe deer: The effects of population density at birth and of habitat quality. Proc. R. Soc. Lond. Ser. B 2002, 269, 747–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewison, A.J.M.; Vincent, J.P.; Bideau, E.; Angibault, J.M.; Putman, R.J. Variation in cohort mandible size as an index of roe deer (Capreolus capreolus) densities and population trends. J. Zool. 1996, 239, 573–581. [Google Scholar] [CrossRef]
- De Felice, E.; Mercati, F.; Pacioni, C.; Catorci, A.; Tardella, F.M.; Brusaferro, A.; Scocco, P. Relation between biometric parameters and autumn-winter food availability in roe deer (Capreolus capreolus) population in central Italy. Eur. Zool. J. 2020, 87, 82–93. [Google Scholar] [CrossRef] [Green Version]
- Azorit, C.; Analla, M.; Muñoz-Cobo, J. Variation of mandible size in red deer Cervus elaphus hispanicus from southern Spain. Acta Theriol. 2003, 48, 221–228. [Google Scholar] [CrossRef]
- Suttie, J.M.; Mitchell, B. Jaw length and hind foot length as measures of skeletal development of red deer (Cervus elaphus). J. Zool. 1983, 200, 431–434. [Google Scholar] [CrossRef]
- Avdić, R.; Hadžiomerović, N.; Tandir, F.; Bejdić, P.; Ćutahija, V. Analysis of morphometric parameters of the Roe deer mandible (Capreolus capreolus) and mandible of the sheep (Ovis aries). Veterinaria 2013, 62, 1–9. [Google Scholar]
- Bland, J.M.; Altman, D. Statistical methods for assessing agreement between two methods of clinical measurement. Lancet 1986, 327, 307–310. [Google Scholar] [CrossRef]
- Bland, J.M.; Altman, D.G. Statistical methods for assessing agreement between two methods of clinical measurement. Int. J. Nurs. Stud. 2010, 47, 931–936. [Google Scholar] [CrossRef]
- Giavarina, D. Understanding Bland Altman analysis. Biochem. Med. 2015, 25, 141–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- And, P.F.; Reig, S. Craniometric variability in two populations of roe deer (Capreolus capreolus) from Spain. J. Zool. 1993, 231, 39–49. [Google Scholar] [CrossRef]
- Langvatn, R. Criteria of Physical Condition, Growth and Development in Cervidae, Suitable for Routine Studies; Nordic Council for Wildlife Research: Stockholm, Sweden, 1977; pp. 1–27. [Google Scholar]
- Blagojević, M.; Milošević-Zlatanović, S. Sexual shape dimorphism in Serbian roe deer (Capreolus capreolus L.). Mamm. Biol. 2011, 76, 735–740. [Google Scholar] [CrossRef]
- Górecki, G.; Komosa, M.; Skubis, J.; Jakubowski, M.; Skorupski, M. Interrelations between mandibular parameters, age and carcass in female fallow deer (Dama dama). Acta Zool. Hungarica 2014, 60, 271–279. [Google Scholar]
- Nugent, G.; Frampton, C. Microgeographic and temporal variation in mandible size within a New Zealand fallow deer (Dama dama) population. J. Appl. Ecol. 1994, 31, 253–262. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | N | Measures Mean and SEM | MLc (mm) | TRLc (mm) | MLG | TRLG | tMLG (mm) | tTRLG (mm) |
|---|---|---|---|---|---|---|---|---|
| Female | 11 | Mean | 147.7 | 69.3 | 5.9 | 2.8 | 147.0 | 69.3 |
| SEM | 1.7 | 0.9 | 0.1 | 0.0 | 1.8 | 1.0 | ||
| Male | 14 | Mean | 146.2 | 69.7 | 5.8 | 2.8 | 145.9 | 69.6 |
| SEM | 1.3 | 0.9 | 0.1 | 0.0 | 1.3 | 0.9 |
| Parameters 1 | Male (Mean ± SD) | Female (Mean ± SD) | p |
|---|---|---|---|
| IMM-CP | 159.5 ± 7.4 | 156.4 ± 15.4 | 0.612 |
| MMM-CP | 86.5 ± 3.9 | 85.7 ± 5.2 | 0.692 |
| CP-MA | 85.8 ± 5.7 | 84.3 ± 6.3 | 0.692 |
| MMM-MA | 52.7 ± 6.4 | 54.3 ± 5.3 | 0.040 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Felice, E.; Pacioni, C.; Tardella, F.M.; Dall’Aglio, C.; Palladino, A.; Scocco, P. A Novel Method for Increasing the Numerousness of Biometrical Parameters Useful for Wildlife Management: Roe Deer Mandible as Bone Model. Animals 2020, 10, 465. https://doi.org/10.3390/ani10030465
De Felice E, Pacioni C, Tardella FM, Dall’Aglio C, Palladino A, Scocco P. A Novel Method for Increasing the Numerousness of Biometrical Parameters Useful for Wildlife Management: Roe Deer Mandible as Bone Model. Animals. 2020; 10(3):465. https://doi.org/10.3390/ani10030465
Chicago/Turabian StyleDe Felice, Elena, Cesare Pacioni, Federico Maria Tardella, Cecilia Dall’Aglio, Antonio Palladino, and Paola Scocco. 2020. "A Novel Method for Increasing the Numerousness of Biometrical Parameters Useful for Wildlife Management: Roe Deer Mandible as Bone Model" Animals 10, no. 3: 465. https://doi.org/10.3390/ani10030465