Betaine Supplementation Improves the Production Performance, Rumen Fermentation, and Antioxidant Profile of Dairy Cows in Heat Stress

 ,
,  ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethical Statement
2.2. Experimental Design and Diets
2.3. Measurement of Temperature–Humidity Index (THI)
2.4. Sample Collection
2.5. Chemical Analysis and Calculations
2.6. Statistical Analysis
3. Results
3.1. Lactation Performance
3.2. Rumen Fermentation
3.3. Nutrient Apparent Digestibility
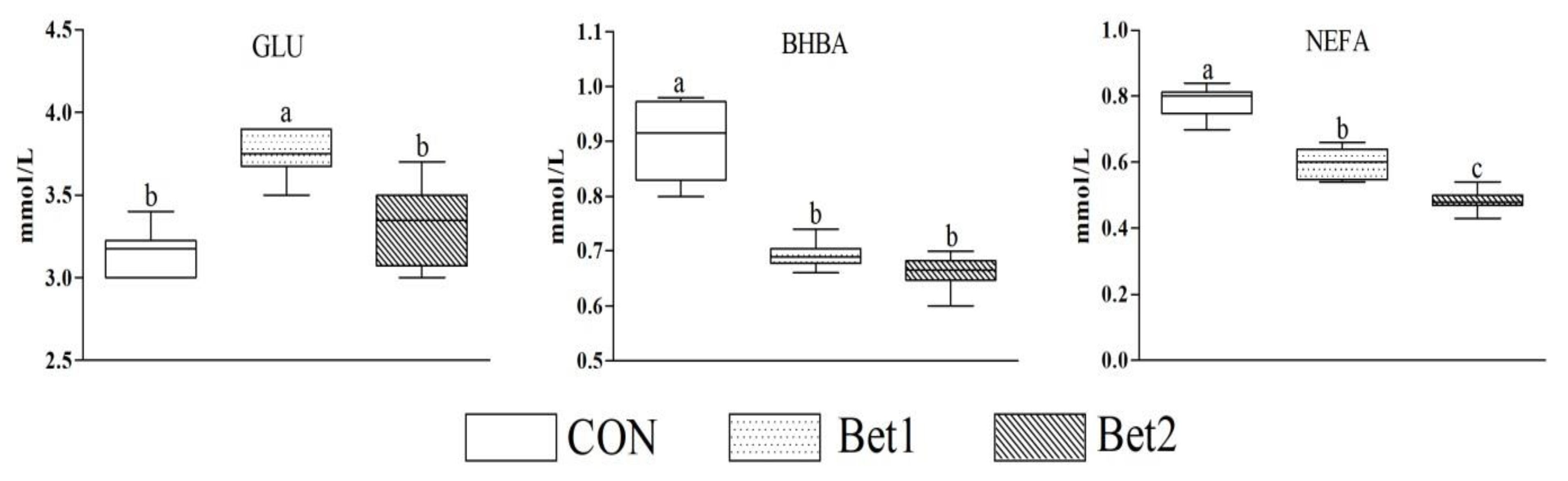
3.4. Serum Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dunshea, F.R.; Oluboyede, K.; DiGiacomo, K.; Leury, B.J.; Cottrell, J.J. Betaine Improves Milk Yield in Grazing Dairy Cows Supplemented with Concentrates at High Temperatures. Animal 2019, 9, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Ying, S.; An, W.; Lian, H.; Zhou, G.; Han, Z. Effects of dietary betaine supplementation subjected to heat stress on milk performances and physiology indices in dairy cow. Gen. and Mol. Res. 2014, 13, 7577–7586. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D. Heat stress interaction with shade and cooling. J. Dairy Sci. 1994, 77, 2044–2050. [Google Scholar] [CrossRef]
- Al-Saiady, M.; Al-Shaikh, M.; Al-Mufarrej, S.; Al-Showeimi, T.; Mogawer, H.; Dirrar, A. Effect of chelated chromium supplementation on lactation performance and blood parameters of Holstein cows under heat stress. Anim. Feed Sci. and Tech. 2004, 117, 223–233. [Google Scholar] [CrossRef]
- Zimbelman, R.; Baumgard, L.; Collier, R. Effects of encapsulated niacin on evaporative heat loss and body temperature in moderately heat-stressed lactating Holstein cows. J. Dairy Sci. 2010, 93, 2387–2394. [Google Scholar] [CrossRef] [PubMed]
- DiGiacomo, K.; Simpson, S.; Leury, B.; Dunshea, F. Dietary betaine impacts the physiological responses to moderate heat conditions in a dose dependent manner in sheep. Animal 2016, 6, 51. [Google Scholar] [CrossRef] [Green Version]
- Hammer, M.A.; Baltz, J.M. Betaine is a highly effective organic osmolyte but does not appear to be transported by established organic osmolyte transporters in mouse embryos. Mol. Rep. and Develop. Incorpor. Gam. Res. 2002, 62, 195–202. [Google Scholar] [CrossRef]
- Cronje, P. Essential role of methyl donors in animal productivity. Anim. Prod. Sci. 2018, 58, 655–665. [Google Scholar] [CrossRef]
- Lai, S.-J.; Lai, M.-C. Characterization and regulation of the osmolyte betaine synthesizing enzymes GSMT and SDMT from halophilic methanogen Methanohalophilus portucalensis. PLoS. ONE 2011, 6, e25090. [Google Scholar] [CrossRef]
- DiGiacomo, K.; Warner, R.; Leury, B.; Gaughan, J.; Dunshea, F. Dietary betaine supplementation has energy-sparing effects in feedlot cattle during summer, particularly in those without access to shade. Anim. Prod. Sci. 2014, 54, 450–458. [Google Scholar] [CrossRef] [Green Version]
- Dunshea, F.; Cadogan, D.; Partridge, G. Dietary betaine and ractopamine combine to increase lean tissue deposition in finisher pigs, particularly gilts. Anim. Prod. Sci. 2008, 49, 65–70. [Google Scholar] [CrossRef]
- Council, N.R. Nutrient requirements of dairy cattle: 2001; National Academies Press: Washington, WA, USA, 2001. [Google Scholar]
- McDowell, R.; Hooven, N.; Camoens, J. Effect of climate on performance of Holsteins in first lactation. J. Dairy Sci. 1976, 59, 965–971. [Google Scholar] [CrossRef]
- Zeng, B.; Sun, J.; Chen, T.; Sun, B.; He, Q.; Chen, X.; Zhang, Y.; Xi, Q. Effects of Moringa oleifera silage on milk yield, nutrient digestibility and serum biochemical indexes of lactating dairy cows. J. Anim. Phys. Anim. Nutri. 2018, 102, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chen, Z.; Zhou, S.; Song, X.; Ouyang, K.; Pan, K.; Xu, L.; Liu, C.; Qu, M. Effects of daidzein on performance, serum metabolites, nutrient digestibility, and fecal bacterial community in bull calves. Anim. Feed Sci. Tech. 2017, 225, 87–96. [Google Scholar] [CrossRef]
- Ramos-Morales, E.; Arco-Pérez, A.; Martín-García, A.I.; Yáñez-Ruiz, D.R.; Frutos, P.; Hervás, G. Use of stomach tubing as an alternative to rumen cannulation to study ruminal fermentation and microbiota in sheep and goats. Anim. Feed Sci. and Tech. 2014, 198, 57–66. [Google Scholar] [CrossRef] [Green Version]
- AOAC International. O_cial Methods of Analysis of AOAC International, 16th ed.; AOAC International: Arlington, VA, USA, 1998; pp. 71–90. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Van Keulen, J.; Young, B. Evaluation of acid-insoluble ash as a natural marker in ruminant digestibility studies. J. Anim. Sci. 1977, 44, 282–287. [Google Scholar] [CrossRef]
- Erwin, E.; Marco, G.; Emery, E. Volatile fatty acid analyses of blood and rumen fluid by gas chromatography. J. Dairy Sci. 1961, 44, 1768–1771. [Google Scholar] [CrossRef]
- Makkar, H.; Sharma, O.; Dawra, R.; Negi, S. Simple determination of microbial protein in rumen liquor. J. Dairy Sci. 1982, 65, 2170–2173. [Google Scholar] [CrossRef]
- Broderick, G.; Kang, J. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Gaughan, J.; Cadogan, D.; Cawdell-Smith, A.; Croft, I.; Van Barneveld, R.; Bryden, W. Improved heat tolerance of cattle by dietary supplementation with osmolytes. Asia Pacific J. Clin. Nutr. 2005, 14. [Google Scholar]
- Gonzalez-Rivas, P.A.; Sullivan, M.; Cottrell, J.J.; Leury, B.J.; Gaughan, J.B.; Dunshea, F.R. Effect of feeding slowly fermentable grains on productive variables and amelioration of heat stress in lactating dairy cows in a sub-tropical summer. Trop. Anim. Heal. Prod. 2018, 50, 1763–1769. [Google Scholar] [CrossRef] [PubMed]
- Hall, L.; Dunshea, F.; Allen, J.; Rungruang, S.; Collier, J.; Long, N.; Collier, R.J. Evaluation of dietary betaine in lactating Holstein cows subjected to heat stress. J. Dairy Sci. 2016, 99, 9745–9753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loest, C.; Titgemeyer, E.; Drouillard, J.; Coetzer, C.; Hunter, R.; Bindel, D.; Lambert, B. Supplemental betaine and peroxide-treated feather meal for finishing cattle. J. Anim. Sci. 2002, 80, 2234–2240. [Google Scholar] [CrossRef]
- Wdowiak-Wróbel, S.; Leszcz, A.; Małek, W. Salt tolerance in Astragalus cicer microsymbionts: The role of glycine betaine in osmoprotection. Curr. Microbiol. 2013, 66, 428–436. [Google Scholar] [CrossRef]
- Peterson, S.E.; Rezamand, P.; Williams, J.; Price, W.; Chahine, M.; McGuire, M. Effects of dietary betaine on milk yield and milk composition of mid-lactation Holstein dairy cows. J. Dairy Sci. 2012, 95, 6557–6562. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, Q.; Yang, W.; Wu, J.; Zhang, W.; Zhang, P.; Dong, K.; Huang, Y. Effects of betaine supplementation on rumen fermentation, lactation performance, feed digestibilities and plasma characteristics in dairy cows. The J. Agric. Sci. 2010, 148, 487–495. [Google Scholar] [CrossRef]
- Eklund, M.; Mosenthin, R.; Tafaj, M.; Wamatu, J. Effects of betaine and condensed molasses solubles on nitrogen balance and nutrient digestibility in piglets fed diets deficient in methionine and low in compatible osmolytes. Arch. Anim. Nutr. 2006, 60, 289–300. [Google Scholar] [CrossRef]
- Davidson, S.; Hopkins, B.; Odle, J.; Brownie, C.; Fellner, V.; Whitlow, L. Supplementing limited methionine diets with rumen-protected methionine, betaine, and choline in early lactation Holstein cows. J. Dairy Sci. 2008, 91, 1552–1559. [Google Scholar] [CrossRef]
- Lu, X.; Wang, J.; Wang, H.; Qu, S. Effects of betaine on body weight and biochemical index in obese rats fed high-fat diet. Acta Nutri. Sini. 2008, 3, 023. [Google Scholar]


{kind=link}
| Ingredient | Contents | Item | Contents |
|---|---|---|---|
| Ingredients | Nutrient Levels | ||
| Corn silage | 36.80 | NEg (MJ/kg) 2 | 5.48 |
| Alfalfa hay | 3.45 | CP | 14.88 |
| Chinese wildrye | 6.01 | NDF | 43.15 |
| Corn | 16.67 | ADF | 25.20 |
| Soybean meal | 3.85 | Ca | 0.89 |
| Beet pulp | 4.28 | P | 0.58 |
| Brewer’s grains | 16.55 | ||
| DDGS | 3.44 | ||
| Cottonseed meal | 3.48 | ||
| Molasses | 3.66 | ||
| Limestone | 0.35 | ||
| CaHPO4 | 0.40 | ||
| Na2CO3 | 0.53 | ||
| NaCl | 0.33 | ||
| Premix 1 | 0.20 | ||
| Total | 100 | ||
| Item | CON | Bet1 | Bet2 | SEM | p |
|---|---|---|---|---|---|
| DMI (kg/d) | 18.83 | 19.99 | 18.90 | 0.158 | 0.081 |
| Milk yield (kg/d) | 21.98 | 23.24 | 22.77 | 0.114 | <0.001 |
| Milk fat (%) | 4.02 | 4.11 | 4.03 | 0.016 | 0.036 |
| Milk protein (%) | 3.12 | 3.15 | 3.13 | 0.006 | 0.041 |
| Lactose (%) | 4.54 | 4.57 | 4.56 | 0.011 | 0.562 |
| SCC (×103/mL) | 392.20 | 289.80 | 297.20b | 7.808 | <0.001 |
| Item | CON | Bet1 | Bet2 | SEM | p |
|---|---|---|---|---|---|
| pH | 6.55 | 6.54 | 6.42 | 0.028 | 0.346 |
| VFA (mmol/L) | 117.20 | 127.00 | 122.40 | 2.208 | 0.034 |
| Acetate (mmol/L) | 68.40 | 71.00 | 72.10 | 0.386 | <0.001 |
| Propionate (mmol/L) | 22.90 | 20.60 | 20.80 | 0.298 | 0.003 |
| Butyrate (mmol/L) | 11.69 | 15.33 | 13.47 | 0.282 | <0.001 |
| Acetate-to-propionate ratio | 3.00 | 3.46 | 3.48 | 0.057 | 0.012 |
| NH3-N (mg/dL) | 15.44 | 11.20 | 10.77 | 0.395 | <0.001 |
| MCP (mg/mL) | 6.55 | 8.69 | 7.21 | 0.469 | <0.001 |
| Item | CON | Bet1 | Bet2 | SEM | p |
|---|---|---|---|---|---|
| DM (%) | 72.44 | 75.00 | 73.40 | 0.213 | <0.001 |
| OM (%) | 62.15 | 65.30 | 64.21 | 0.948 | 0.035 |
| CP (%) | 65.82 | 72.25 | 69.55 | 0.498 | <0.001 |
| EE (%) | 71.04 | 70.14 | 70.05 | 1.072 | 0.358 |
| NDF (%) | 51.65 | 54.41 | 53.84 | 0.866 | 0.033 |
| ADF (%) | 44.50 | 48.24 | 45.28 | 0.303 | <0.001 |
| Item | CON | Bet1 | Bet2 | SEM | p |
|---|---|---|---|---|---|
| T-AOC (U/L) | 1.06 | 2.32 | 2.39 | 0.613 | <0.001 |
| GSH-Px (U/L) | 116.06 | 153.74 | 93.24 | 4.64 | <0.001 |
| MDA (U/L) | 1.26 | 2.09 | 2.39 | 0.089 | <0.001 |
| SOD (U/L) | 13.55 | 14.40 | 16.43 | 0.235 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, A.M.; Ma, J.; Wang, Z.; Zou, H.; Hu, R.; Peng, Q. Betaine Supplementation Improves the Production Performance, Rumen Fermentation, and Antioxidant Profile of Dairy Cows in Heat Stress. Animals 2020, 10, 634. https://doi.org/10.3390/ani10040634
Shah AM, Ma J, Wang Z, Zou H, Hu R, Peng Q. Betaine Supplementation Improves the Production Performance, Rumen Fermentation, and Antioxidant Profile of Dairy Cows in Heat Stress. Animals. 2020; 10(4):634. https://doi.org/10.3390/ani10040634
Chicago/Turabian StyleShah, Ali Mujtaba, Jian Ma, Zhisheng Wang, Huawei Zou, Rui Hu, and Quanhui Peng. 2020. "Betaine Supplementation Improves the Production Performance, Rumen Fermentation, and Antioxidant Profile of Dairy Cows in Heat Stress" Animals 10, no. 4: 634. https://doi.org/10.3390/ani10040634
APA StyleShah, A. M., Ma, J., Wang, Z., Zou, H., Hu, R., & Peng, Q. (2020). Betaine Supplementation Improves the Production Performance, Rumen Fermentation, and Antioxidant Profile of Dairy Cows in Heat Stress. Animals, 10(4), 634. https://doi.org/10.3390/ani10040634