Effect of MSTN Mutation on Growth and Carcass Performance in Duroc × Meishan Hybrid Population

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statement
2.2. Animals and Management
2.3. Traits Measurements
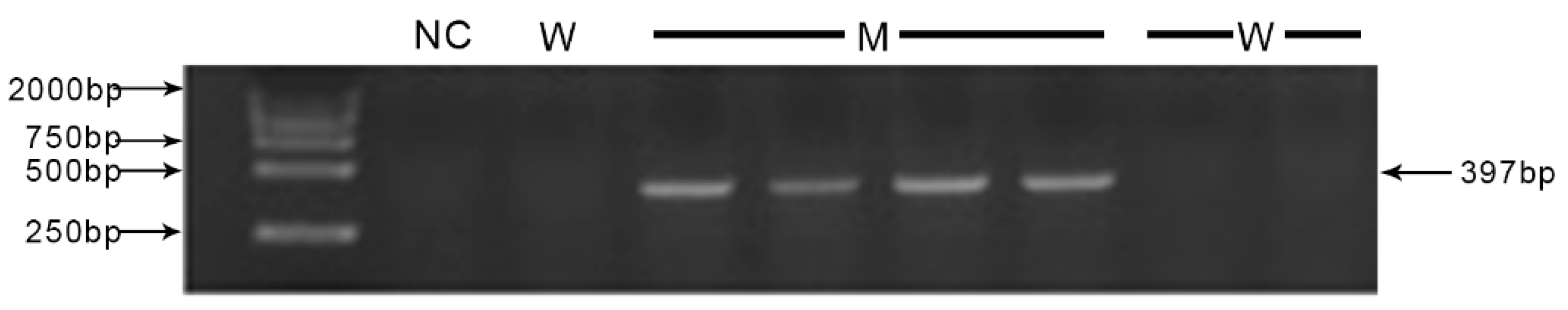
2.4. Genotype Assessments
2.5. RNA Extraction and Real-Time Quantitative Polymerase Chain Reaction
2.6. Statistical Analyses
3. Results
3.1. Reproductive Performance
3.2. Genotyping and MSTN Expression Detection
3.3. Body Size and Growth Performance
3.4. Carcass Performance and Meat Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gjerlaug-Enger, E.; Kongsro, J.; Odegard, J.; Aass, L.; Vangen, O. Genetic parameters between slaughter pig efficiency and growth rate of different body tissues estimated by computed tomography in live boars of Landrace and Duroc. Animal 2012, 6, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.H.; McKeith, F.K.; Novakofski, J.; Carr, T.R. Carcass and muscle characteristics of yorkshire, meishan, yorkshire × meishan, meishan × yorkshire, fengjing × yorkshire, and minzhu × yorkshire pigs. J. Anim. Sci. 1993, 71, 3344–3349. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.Z.; Zhu, L.; Tang, G.Q.; Li, M.Z.; Jiang, A.A.; Cen, W.M.; Xing, S.H.; Chen, J.N.; Wen, A.X.; He, T.; et al. Carcass and meat quality traits of four commercial pig crossbreeds in China. Genet. Mol. Res. 2012, 11, 4447–4455. [Google Scholar] [CrossRef]
- Cameron, N.D. Genetic and phenotypic parameters for carcass traits, meat and eating quality traits in pigs. Livest. Prod. Sci. 1990, 26, 119–135. [Google Scholar] [CrossRef]
- Newcom, D.W.; Stalder, K.J.; Baas, T.J.; Goodwin, R.N.; Parrish, F.C.; Wiegand, B.R. Breed differences and genetic parameters of myoglobin concentration in porcine longissimus muscle. J. Anim. Sci. 2004, 82, 2264–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPherron, A.; Lawler, A.; Lee, S. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lee, S.J. Suppression of body fat accumulation in myostatin-deficient mice. J. Clin. Investig. 2002, 109, 595–601. [Google Scholar] [CrossRef]
- Lee, C.-Y.; Hu, S.-Y.; Gong, H.-Y.; Chen, M.H.-C.; Lu, J.-K.; Wu, J.-L. Suppression of myostatin with vector-based RNA interference causes a double-muscle effect in transgenic zebrafish. Biochem. Biophys. Res. Commun. 2009, 387, 766–771. [Google Scholar] [CrossRef]
- Sonstegard, T.S.; Rohrer, G.A.; Smith, T.P.L. Myostatin maps to porcine chromosome 15 by linkage and physical analyses. Anim. Genet. 1998, 29, 19–22. [Google Scholar] [CrossRef]
- Stratil, A.; Kopecny, M. Genomic organization, sequence and polymorphism of the porcine myostatin (GDF8; MSTN) gene. Anim. Genet. 1999, 30, 468–470. [Google Scholar] [CrossRef]
- Kambadur, R.; Sharma, M.; Smith, T.P.L.; Bass, J.J. Mutations in myostatin (GDF8) in double-muscled Belgian blue and Piedmontese cattle. Genome Res. 1997, 7, 910–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPherron, A.C.; Lee, S.J. Double muscling in cattle due to mutations in the myostatin gene. Proc. Natl. Acad. Sci. USA 1997, 94, 12457–12461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, P.F. Double muscling in cattle: A review. Aust. J. Agric. Res. 1995, 46, 1493–1515. [Google Scholar] [CrossRef]
- Casas, E.; Bennett, G.L.; Smith, T.P.L.; Cundiff, L.V. Association of myostatin on early calf mortality, growth, and carcass composition traits in crossbred cattle. J. Anim. Sci. 2004, 82, 2913–2918. [Google Scholar] [CrossRef] [PubMed]
- Casas, E.; Keele, J.W.; Fahrenkrug, S.C.; Smith, T.P.L.; Cundiff, L.V.; Stone, R.T. Quantitative analysis of birth, weaning, and yearling weights and calving difficulty in Piedmontese crossbreds segregating an inactive myostatin allele. J. Anim. Sci. 1999, 77, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Wiener, P.; Smith, J.A.; Lewis, A.M.; Woolliams, J.A.; Williams, J.L. Muscle-related traits in cattle: The role of the myostatin gene in the South Devon breed. Genet. Sel. Evol. 2002, 34, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Arthur, P.F.; Makarechian, M.; Price, M.A. Incidence of dystocia and perinatal calf mortality resulting from reciprocal crossing of double-muscled and normal cattle. Can. Vet. J. Rev. Vet. Can. 1988, 29, 163–167. [Google Scholar]
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academy Press: Washington, DC, USA, 2012. [Google Scholar]
- AOAC. Official Methods of Analysis, 18th ed.; AOAC: Arlington, VA, USA, 2004; Volume 2. [Google Scholar]
- Xiao, R.J.; Xu, Z.R.; Chen, H.L. Effects of ractopamine at different dietary protein levels on growth performance and carcass characteristics in finishing pigs. Anim. Feed Sci. Technol. 1999, 79, 119–127. [Google Scholar] [CrossRef]
- Alonso, V.; Campo, M.D.M.; Espanol, S.; Roncales, P.; Beltran, J.A. Effect of crossbreeding and gender on meat quality and fatty acid composition in pork. Meat Sci. 2009, 81, 209–217. [Google Scholar] [CrossRef]
- Miao, Z.G.; Wang, L.J.; Xu, Z.R.; Huang, J.F.; Wang, Y.R. Developmental changes of carcass composition, meat quality and organs in the Jinhua pig and Landrace. Animal 2009, 3, 468–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, L.L.; Tang, M.X.; Yang, J.Z.; Wang, Q.Q.; Cai, C.B.; Jiang, S.W.; Li, H.G.; Jiang, K.; Gao, P.F.; Ma, D.Z.; et al. Targeted mutations in myostatin by zinc-finger nucleases result in double-muscled phenotype in Meishan pigs. Sci. Rep. 2015, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.D.; Kim, S.; Zhu, H.Y.; Jin, L.; Guo, Q.; Li, X.C.; Zhang, Y.C.; Xing, X.X.; Xuan, M.F.; Zhang, G.L.; et al. Generation of cloned adult muscular pigs with myostatin gene mutation by genetic engineering. RSC Adv. 2017, 7, 12541–12549. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.K.; Tang, X.C.; Xie, Z.C.; Zou, X.D.; Li, M.J.; Yuan, H.M.; Guo, N.N.; Ouyang, H.S.; Jiao, H.P.; Pang, D.X. CRISPR/Cas9-mediated knockout of myostatin in Chinese indigenous Erhualian pigs. Transgenic Res. 2017, 26, 799–805. [Google Scholar] [CrossRef]
- Han, S.Z.; Jin, S.S.; Xuan, M.F.; Guo, Q.; Luo, Z.B.; Wang, J.X.; Kang, J.D.; Yin, X.J. Semen quality and fertilization ability of myostatin-knockout boars. Theriogenology 2019, 135, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.J.; Haley, C.S. Comparative farrowing to weaning performance in meishan and large white-pigs and their crosses. Anim. Sci. 1995, 60, 269–280. [Google Scholar] [CrossRef]
- Haley, C.S.; Lee, G.J.; Ritchie, M. Comparative reproductive-performance in meishan and large white-pigs and their crosses. Anim. Sci. 1995, 60, 259–267. [Google Scholar] [CrossRef]
- Dunshea, F.R.; King, R.H.; Campbell, R.G.; Sainz, R.D.; Kim, Y.S. Interrelationships between sex and ractopamine on protein and lipid deposition in rapidly growing pigs. J. Anim. Sci. 1993, 71, 2919–2930. [Google Scholar] [CrossRef]
- Campbell, R.G.; Steele, N.C.; Caperna, T.J.; McMurtry, J.P.; Solomon, M.B.; Mitchell, A.D. Interrelationships between sex and exogenous growth-hormone administration on performance, body-composition and protein and fat accretion of growing-pigs. J. Anim. Sci. 1989, 67, 177–186. [Google Scholar] [CrossRef]
- Latorre, M.A.; Lazaro, R.; Valencia, D.G.; Medel, P.; Mateos, G.G. The effects of gender and slaughter weight on the growth performance, carcass traits, and meat quality characteristics of heavy pigs. J. Anim. Sci. 2004, 82, 526–533. [Google Scholar] [CrossRef]
- Renaudeau, D.; Giorgi, M.; Silou, F.; Weisbecker, J.L. Effect of breed (lean or fat pigs) and sex on performance and feeding behaviour of group housed growing pigs in a tropical climate. Asian Australas. J. Anim. Sci. 2006, 19, 593–600. [Google Scholar] [CrossRef]
- Latorre, M.A.; Lazaro, R.; Gracia, M.I.; Nieto, M.; Mateos, G.G. Effect of sex and terminal sire genotype on performance, carcass characteristics, and meat quality of pigs slaughtered at 117 kg body weight. Meat Sci. 2003, 65, 1369–1377. [Google Scholar] [CrossRef]
- Peinado, J.; Medel, P.; Fuentetaja, A.; Mateos, G.G. Influence of sex and castration of females on growth performance and carcass and meat quality of heavy pigs destined for the dry-cured industry. J. Anim. Sci. 2008, 86, 1410–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; McFarlane, C.; Lokireddy, S.; Masuda, S.; Ge, X.; Gluckman, P.D.; Sharma, M.; Kambadur, R. Inhibition of myostatin protects against diet-induced obesity by enhancing fatty acid oxidation and promoting a brown adipose phenotype in mice. Diabetologia 2012, 55, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.B.; Qian, L.L.; Jiang, S.W.; Sun, Y.D.; Wang, Q.Q.; Ma, D.Z.; Xiao, G.J.; Li, B.; Xie, S.S.; Gao, T.; et al. Loss-of-function myostatin mutation increases insulin sensitivity and browning of white fat in Meishan pigs. Oncotarget 2017, 8, 34911–34922. [Google Scholar] [CrossRef] [Green Version]
- Legault, C. Selection of breeds, strains and individual pigs for prolificacy. J. Reprod. Fertil. Suppl. 1985, 33, 151–166. [Google Scholar]
- White, B.R.; Lan, Y.H.; McKeith, F.K.; Novakofski, J.; Wheeler, M.B.; McLaren, D.G. Growth and body-composition of meishan and yorkshire barrows and gilts. J. Anim. Sci. 1995, 73, 738–749. [Google Scholar] [CrossRef]
- Bidanel, J.P.; Caritez, J.C.; Gruand, J.; Legault, C. Growth, carcass and meat quality performance of crossbred pigs with graded proportions of meishan genes. Genet. Sel. Evol. 1993, 25, 83–99. [Google Scholar] [CrossRef]
- Muller, E.; Moser, G.; Bartenschlager, H.; Geldermann, H. Trait values of growth, carcass and meat quality in Wild Boar, Meishan and Pietrain pigs as well as their crossbred generations. J. Anim. Breed. Genet. Z. Tierz. Zucht. 2000, 117, 189–202. [Google Scholar] [CrossRef]
- Verbeke, W.; Van Oeckel, M.J.; Warnants, N.; Viaene, J.; Boucque, C.V. Consumer perception, facts and possibilities to improve acceptability of health and sensory characteristics of pork. Meat Sci. 1999, 53, 77–99. [Google Scholar] [CrossRef]
- Fortin, A.; Robertson, W.M.; Tong, A.K.W. The eating quality of Canadian pork and its relationship with intramuscular fat. Meat Sci. 2005, 69, 297–305. [Google Scholar] [CrossRef]
- Schwab, C.R.; Baas, T.J.; Stalder, K.J.; Nettleton, D. Results from six generations of selection for intramuscular fat in Duroc swine using real-time ultrasound. I. Direct and correlated phenotypic responses to selection. J. Anim. Sci. 2009, 87, 2774–2780. [Google Scholar] [CrossRef] [PubMed]
- Devol, D.L.; McKeith, F.K.; Bechtel, P.J.; Novakofski, J.; Shanks, R.D.; Carr, T.R. Variation in composition and palatability traits and relationships between muscle characteristics and palatability in a random sample of pork carcasses. J. Anim. Sci. 1988, 66, 385–395. [Google Scholar] [CrossRef]
- Newcom, D.W.; Baas, T.J.; Schwab, C.R.; Stalder, K.J. Genetic and phenotypic relationships between individual subcutaneous backfat layers and percentage of longissimus intramuscular fat in Duroc swine. J. Anim. Sci. 2005, 83, 316–323. [Google Scholar] [CrossRef]
- Vanderwal, P.G.; Devries, A.G.; Eikelenboom, G. Predictive value of slaughterhouse measurements of ultimate pork quality in 7 halothane negative yorkshire populations. Meat Sci. 1995, 40, 183–191. [Google Scholar] [CrossRef]
- Sellier, P. Genetics of meat and carcass traits. In The Genetics of the Pig; Rothschild, M.F., Ruvinsky, A., Eds.; CABI: Wallingford, UK, 1998. [Google Scholar]
- Grochowska, E.; Borys, B.; Lisiak, D.; Mroczkowski, S. Genotypic and allelic effects of the myostatin gene (MSTN) on carcass, meat quality, and biometric traits in Colored Polish. Merino sheep. Meat Sci. 2019, 151, 4–17. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, B.A.; Dennis, J.A.; Healy, P.J.; McKiernan, W.A.; Greenwood, P.L.; Cafe, L.M.; Perry, D.; Walker, K.H.; Marsh, I.; Parnell, P.F.; et al. Quantitative analysis of performance, carcass and meat quality traits in cattle from two Australian beef herds in which a null myostatin allele is segregating. Anim. Prod. Sci. 2009, 49, 297–305. [Google Scholar] [CrossRef]
- den Hertog-Meischke, M.J.A.; van Laack, R.; Smulders, F.J.M. The water-holding capacity of fresh meat. Vet. Q. 1997, 19, 175–181. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Item | 20–50 kg | 50 kg-Slaughter |
|---|---|---|
| Ingredients (g/kg) | ||
| Corn | 715 | 787 |
| Soybean meal | 250 | 180 |
| Dicalcium phosphate | 11 | 10 |
| Limestone | 8 | 8 |
| Lysine-HCl | 2 | 1 |
| Salt | 4 | 4 |
| Premix 1 | 10 | 10 |
| Total | 1000 | 1000 |
| Calculated nutrient levels | ||
| Digestible energy (MJ/kg) | 13.77 | 13.80 |
| Crude protein (%) | 17.27 | 14.80 |
| Calcium (%) | 0.71 | 0.66 |
| Total phosphorus (%) | 0.60 | 0.55 |
| Lysine (%) | 1.02 | 0.80 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| PZFN1/PZFN2 | TACAAGGTATACTGGAATCCGATCT | GCAAAGTAAAAGTATCAAGAGGGTA |
| MSTN-intact | TGAGAATGGTCATGATCTTGCTG | TCCAGTCCCATCCAAAAGCT |
| MSTN-total GAPDH | AGTGATGGCTCCTTGGAAGAGTGAAGGTCGGAGTGAACG | TGTAGGAGTCTTGACGGGTCTCGCTCCTGGAAGATGGTG |
| BSN | SSN | LS | Alive Litter Size | LW (kg) | BW (kg) | ||
|---|---|---|---|---|---|---|---|
| Male | Female | Subtotal | |||||
| 452 | 678 | 10 | 4 | 5 | 9 | 9.26 | 1.03 |
| 452 | 8978 | 9 | 3 | 6 | 9 | 10.64 | 1.18 |
| 452 | 3624 | 15 | 6 | 7 | 13 | 13.03 | 1.00 |
| 452 | 8834 | 12 | 6 | 5 | 11 | 10.51 | 0.96 |
| 452 | 2228 | 12 | 5 | 3 | 8 | 9.29 | 1.16 |
| 452 | 1178 | 15 | 8 | 5 | 13 | 13.34 | 1.03 |
| Total | 73 | 32 | 31 | 63 | |||
| Item | Barrow | Female | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| MSTN+/+ (n = 8) | MSTN+/− (n = 13) | MSTN+/+ (n = 7) | MSTN+/− (n = 15) | S | G | S × G | BW | ||
| BW (kg) | 102.29 | 101.25 | 103.57 | 99.65 | 3.37 | 0.173 | 0.283 | 0.814 | - |
| ADG (g) | 655.11 | 647.96 | 674.41 | 643.27 | 44.31 | 0.327 | 0.460 | 0.478 | 0.001 |
| LBF (mm) | 22.13 | 20.83 | 21.77 | 19.57 | 2.71 | 0.639 | 0.300 | 0.662 | 0.001 |
| Item (cm) | Barrow | Female | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| MSTN+/+ (n = 8) | MSTN+/− (n = 13) | MSTN+/+ (n = 7) | MSTN+/− (n = 15) | S | G | S × G | BW | ||
| BL | 117.57 | 114.36 | 114.25 | 111.31 | 4.69 | 0.021 | 0.102 | 0.994 | 0.002 |
| BH | 63.57 | 61.08 | 58.87 | 59.58 | 2.63 | 0.001 | 0.489 | 0.022 | 0.001 |
| ChG | 110.00 | 109.00 | 107.38 | 106.85 | 4.43 | 0.839 | 0.300 | 0.147 | <0.001 |
| AG | 126.43 | 123.25 | 126.50 | 122.77 | 2.80 | 0.440 | 0.088 | 0.826 | <0.001 |
| CaG | 16.71 | 16.25 | 16.13 | 16.31 | 1.12 | 0.349 | 0.966 | 0.457 | 0.038 |
| HG | 82.50 | 83.83 | 86.57 | 84.62 | 3.94 | 0.045 | 0.716 | 0.184 | 0.007 |
| Item | Barrow | Female | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| MSTN+/+ (n = 8) | MSTN+/− (n = 13) | MSTN+/+ (n = 7) | MSTN+/− (n = 15) | S | G | S × G | BW | ||
| Dressing yield(%) | 71.65 | 73.27 | 72.83 | 73.82 | 1.67 | 0.043 | 0.001 | 0.479 | 0.029 |
| % of carcass | |||||||||
| Lean (%) | 46.37 | 55.41 | 48.42 | 54.86 | 3.97 | 0.449 | <0.001 | 0.204 | 0.017 |
| Fat (%) | 37.66 | 29.18 | 35.98 | 30.27 | 4.36 | 0.718 | <0.001 | 0.195 | 0.002 |
| Skeleton (%) | 7.57 | 7.39 | 7.75 | 7.51 | 0.90 | 0.308 | 0.048 | 0.894 | 0.001 |
| Skin (%) | 8.40 | 8.08 | 7.85 | 7.37 | 0.89 | 0.005 | 0.016 | 0.603 | 0.009 |
| ABT(mm) | 33.98 | 27.69 | 30.39 | 26.61 | 5.74 | 0.124 | 0.043 | 0.352 | <0.001 |
| ST(mm) | 3.55 | 3.69 | 3.88 | 3.56 | 0.64 | 0.600 | 0.635 | 0.228 | 0.249 |
| LMA (cm2) | 24.31 | 26.75 | 25.18 | 28.91 | 4.65 | 0.328 | 0.003 | 0.519 | <0.001 |
| Item | Barrow | Female | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| MSTN+/+ (n = 8) | MSTN+/− (n = 13) | MSTN+/+ (n = 7) | MSTN+/− (n = 15) | S | G | S × G | BW | ||
| pH1 (45min) | 6.58 | 6.65 | 6.46 | 6.42 | 0.08 | 0.018 | 0.851 | 0.588 | 0.950 |
| pH2 (24h) | 5.76 | 5.73 | 5.73 | 5.68 | 0.09 | 0.339 | 0.720 | 0.422 | 0.692 |
| Color parameters | |||||||||
| Lightness (L*) | 41.71 | 42.13 | 41.95 | 42.24 | 4.36 | 0.444 | 0.089 | 0.229 | 0.517 |
| Redness (a*) | 10.87 | 11.12 | 10.72 | 11.68 | 0.94 | 0.272 | 0.066 | 0.606 | 0.936 |
| Yellowness (b*) | 4.75 | 4.34 | 4.36 | 4.32 | 0.51 | 0.365 | 0.626 | 0.157 | 0.385 |
| Drip loss (%) | 1.32 | 1.08 | 1.53 | 1.37 | 0.24 | 0.087 | 0.168 | 0.084 | 0.069 |
| IMF (%) | 4.59 | 3.05 | 4.36 | 2.66 | 0.64 | 0.497 | 0.010 | 0.942 | 0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Li, R.; Wei, Y.; Meng, X.; Wang, B.; Zhang, Z.; Wu, W.; Liu, H. Effect of MSTN Mutation on Growth and Carcass Performance in Duroc × Meishan Hybrid Population. Animals 2020, 10, 932. https://doi.org/10.3390/ani10060932
Li W, Li R, Wei Y, Meng X, Wang B, Zhang Z, Wu W, Liu H. Effect of MSTN Mutation on Growth and Carcass Performance in Duroc × Meishan Hybrid Population. Animals. 2020; 10(6):932. https://doi.org/10.3390/ani10060932
Chicago/Turabian StyleLi, Weijian, Rongyang Li, Yinghui Wei, Xueqing Meng, Binbin Wang, Zengkai Zhang, Wangjun Wu, and Honglin Liu. 2020. "Effect of MSTN Mutation on Growth and Carcass Performance in Duroc × Meishan Hybrid Population" Animals 10, no. 6: 932. https://doi.org/10.3390/ani10060932