Behavioural Patterns and Postnatal Development in Pups of the Asian Parti-Coloured Bat, Vespertilio sinensis

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
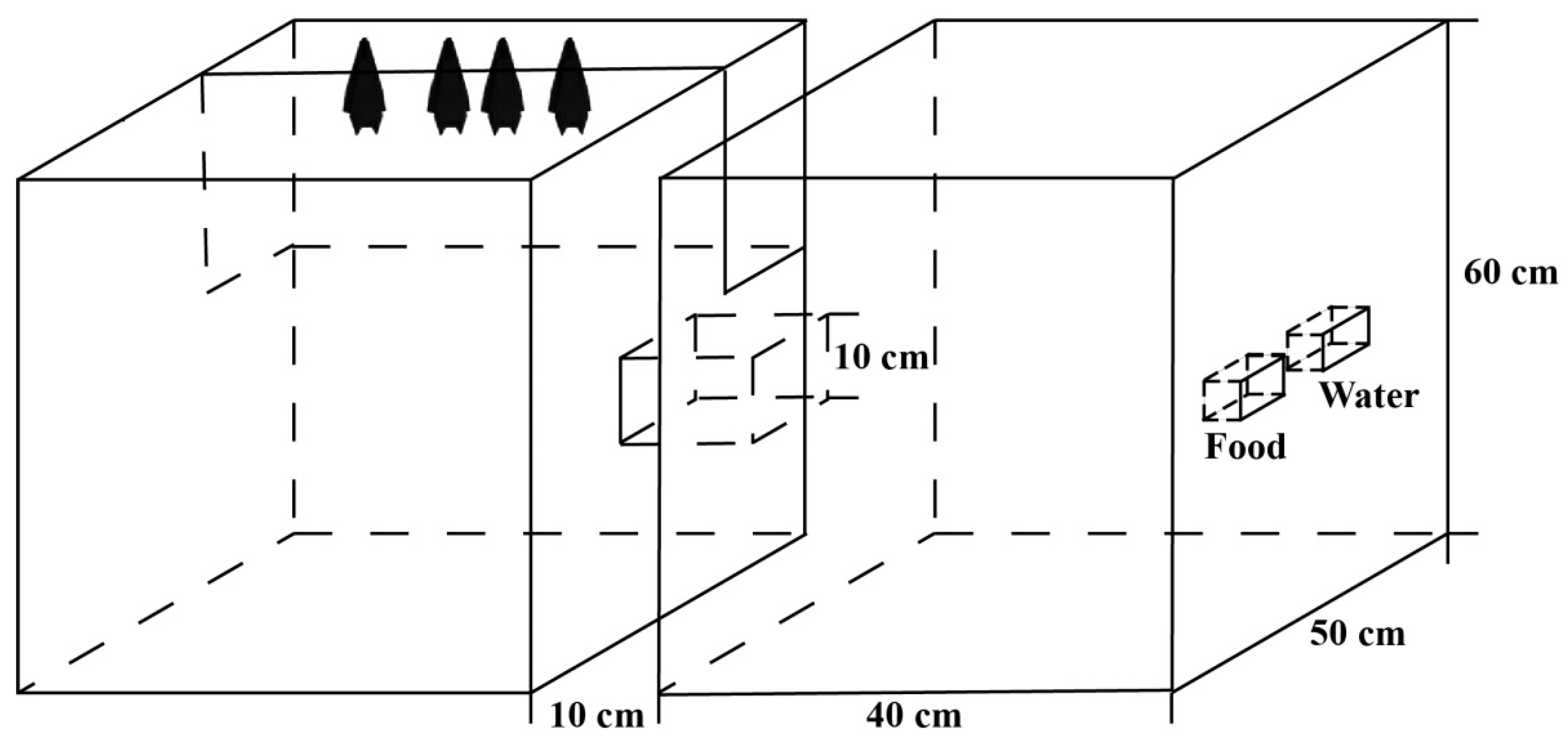
2.1. Sample Collection and Domestication
2.2. Behaviour Observation and Recording
2.3. Scaled Mass Index
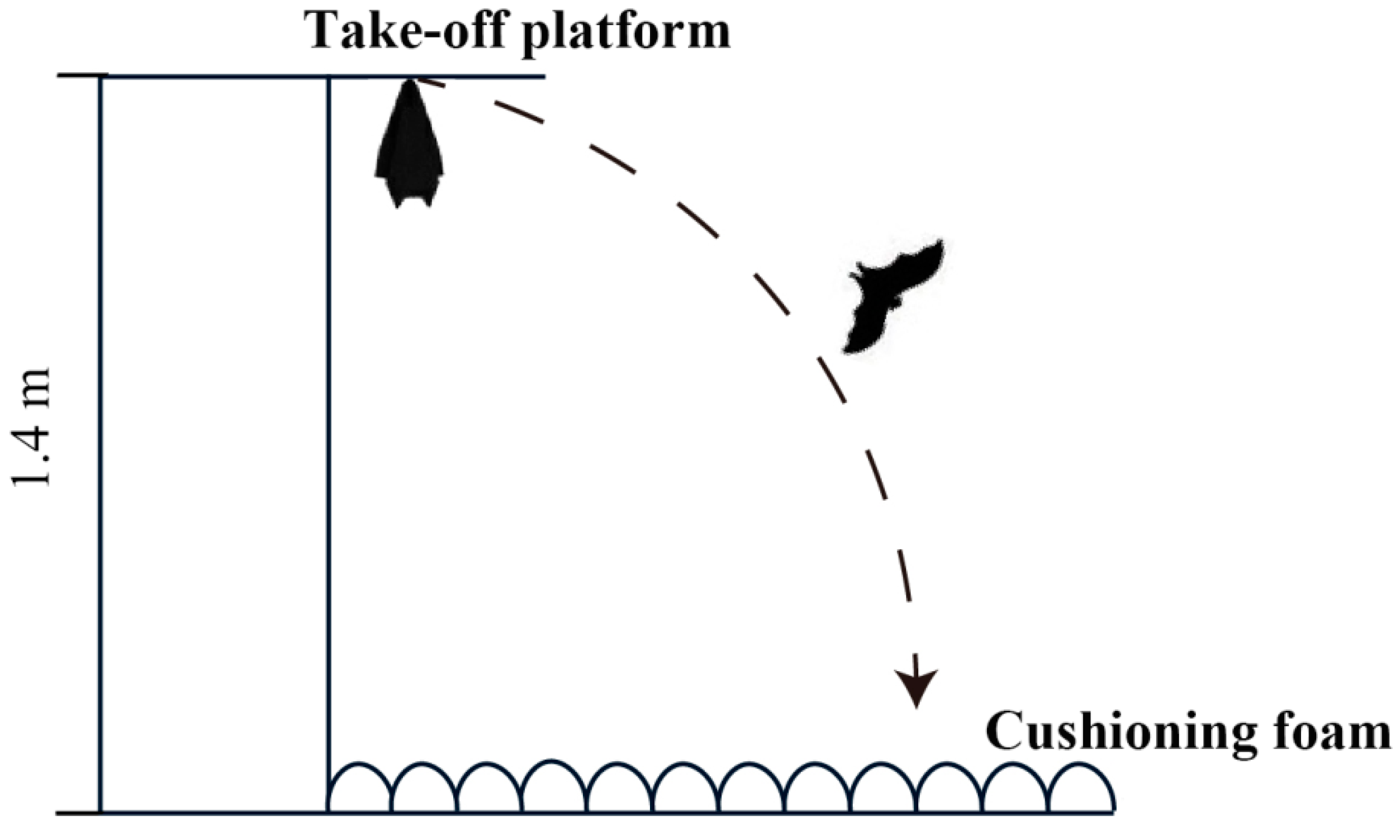
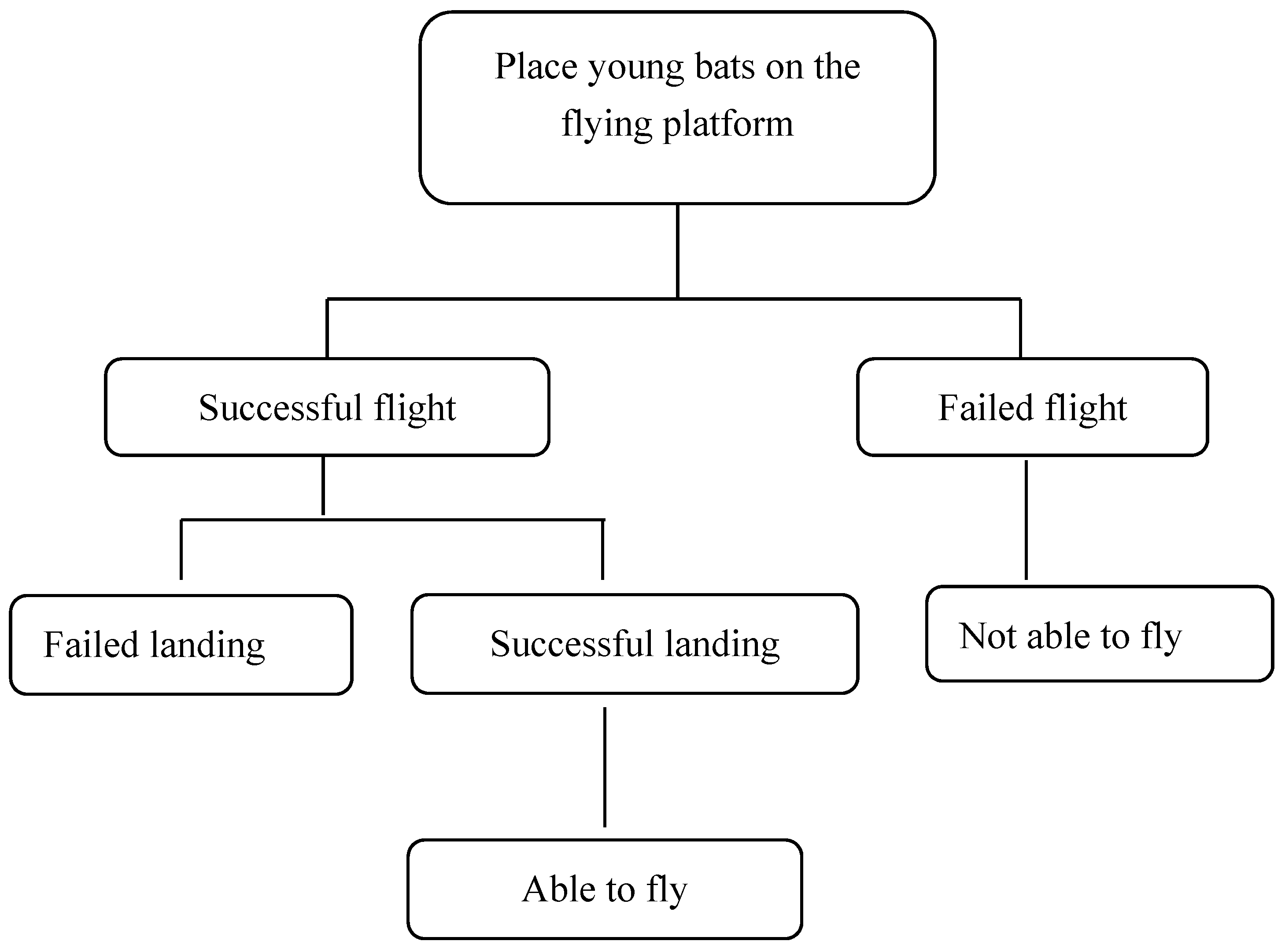
2.4. Flight Ability Experiment
2.5. Statistical Analysis
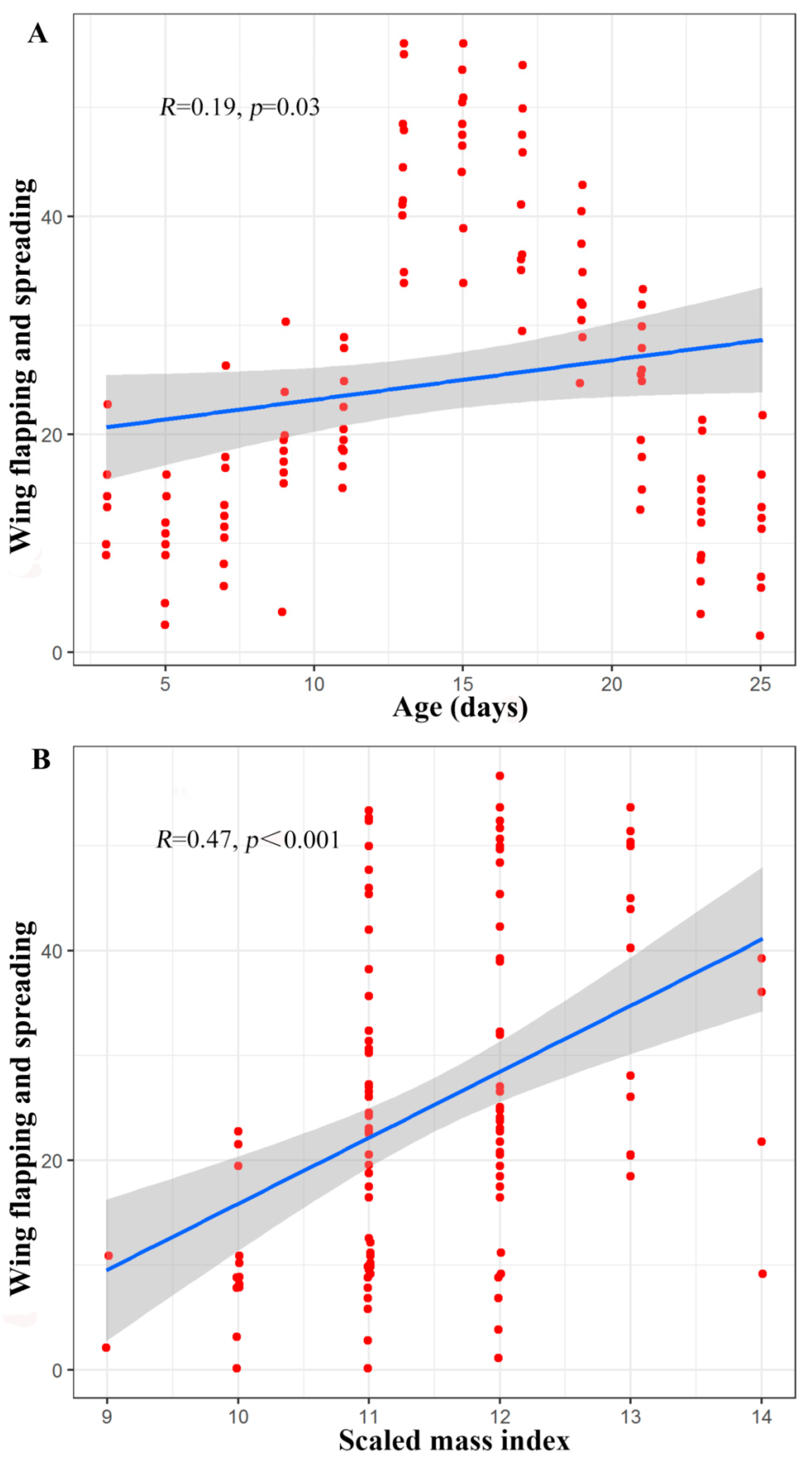
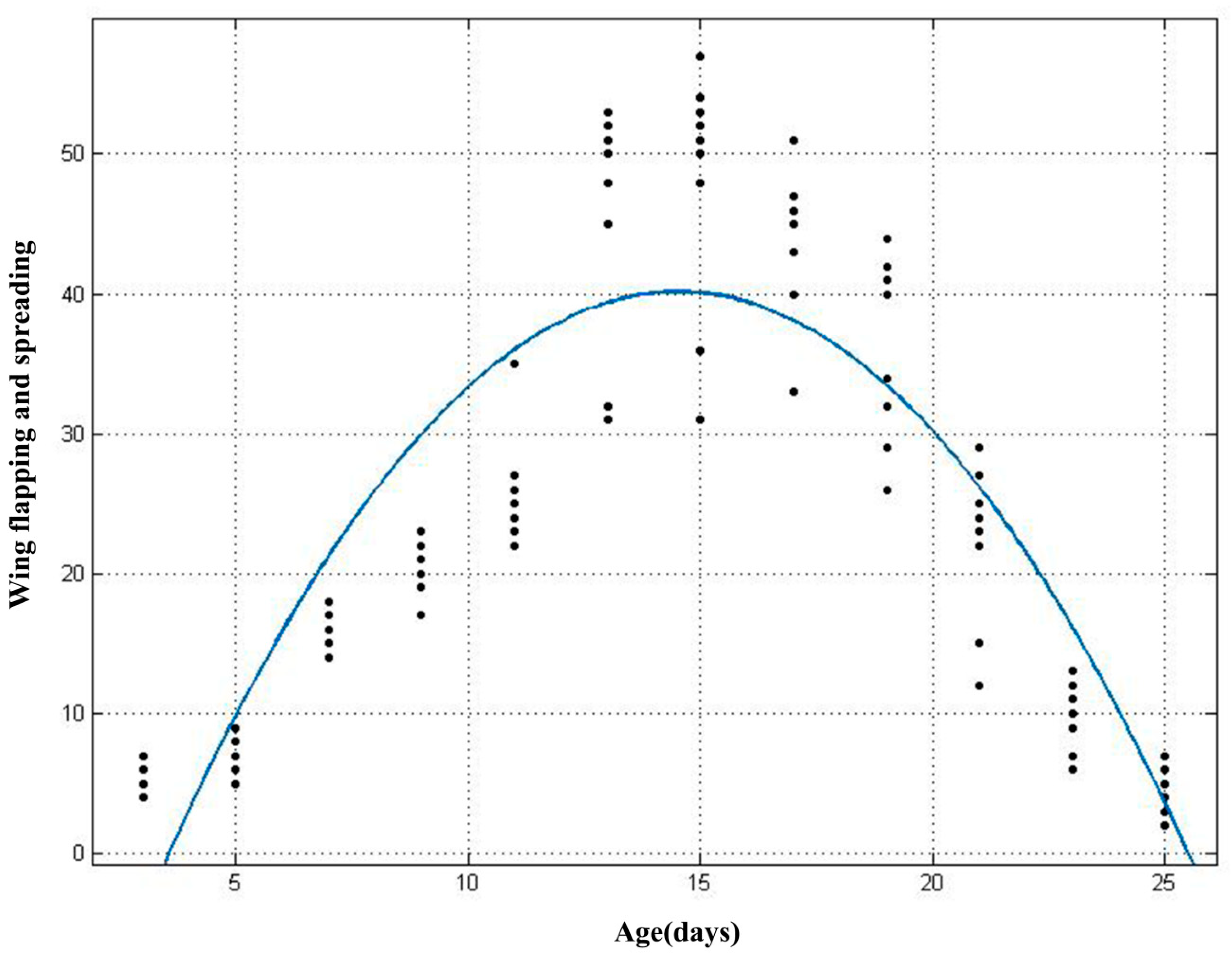
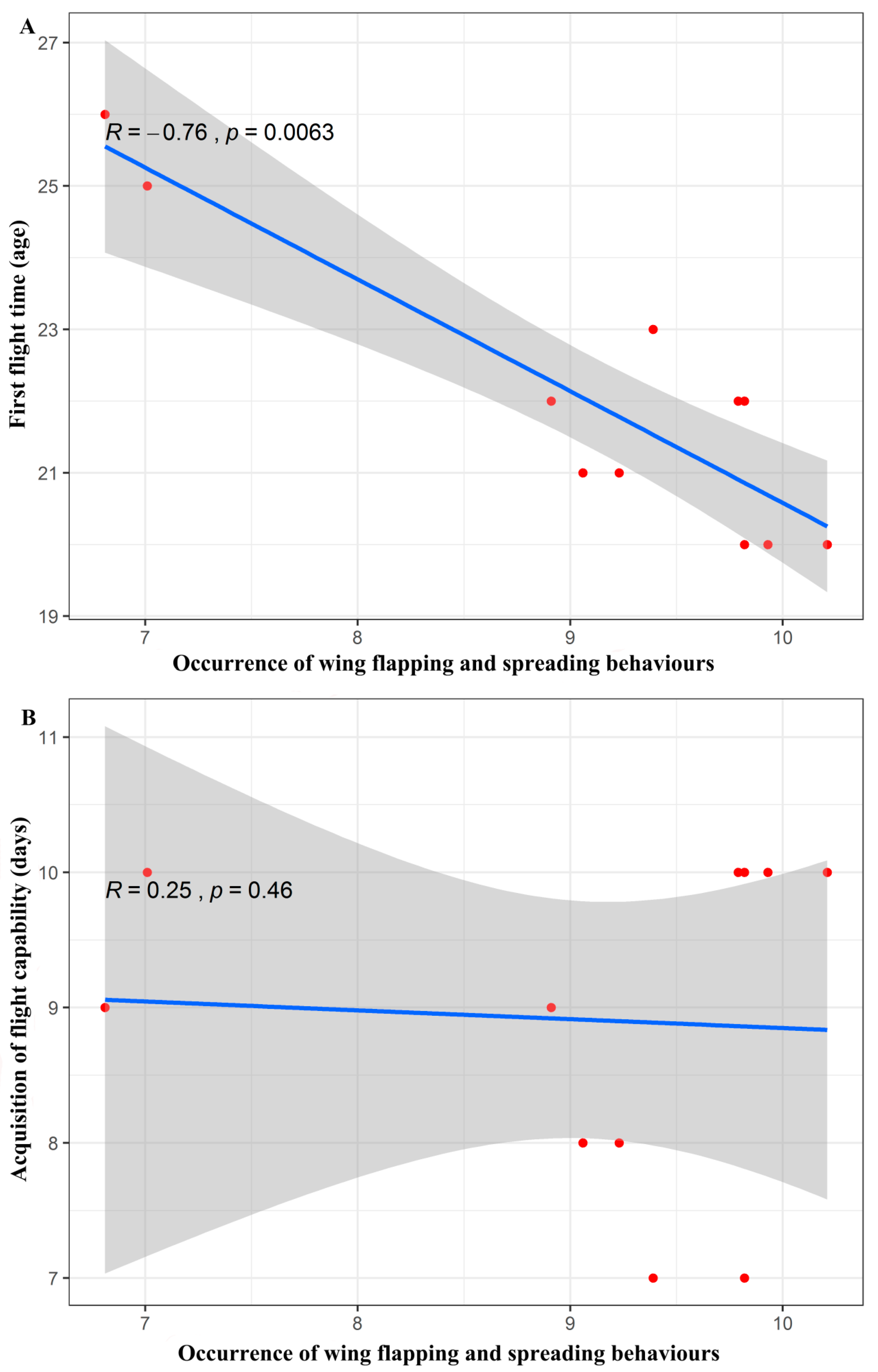
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tinbergen, N. On aims and methods of ethology. Z. Tierpsychol. 1963, 20, 410–433. [Google Scholar] [CrossRef]
- Bolhuis, J.; Hogan, J. The development of behaviour: Trends since tinbergen (1963). Anim. Biol. 2005, 55, 371–398. [Google Scholar]
- Welker, W. Ontogeny of play and exploratory behaviors: A definition of problems and a search for new conceptual solutions. In The Ontogeny of Vertebrate Behavior; Elsevier: Amsterdam, The Netherlands, 1971; pp. 171–228. [Google Scholar]
- Bekoff, M. The development of social interaction, play, and metacommunication in mammals: An ethological perspective. Q. Rev. Biol. 1972, 47, 412–434. [Google Scholar] [CrossRef] [Green Version]
- Nowak, R.M.; Walker, E.P. Walker’s Bats of the World; Johns Hopkins University Press: Baltimore, MD, USA, 1994. [Google Scholar]
- Simmons, N.B. Order Chiroptera. In Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; Wilson, D.E., Reeder, D.M., Eds.; Johns Hopkins University Press: London, UK, 2005; pp. 312–529. [Google Scholar]
- Simmons, N.B.; Seymour, K.L.; Habersetzer, J.R.; Gunnell, G.F. Primitive early eocene bat from wyoming and the evolution of flight and echolocation. Nature 2008, 451, 818–821. [Google Scholar] [CrossRef] [PubMed]
- Norberg, U.M. Allometry of bat wings and legs and comparison with bird wings. Philos. Trans. R. Soc. Lond. 1981, 292, 359–398. [Google Scholar]
- Norberg, U.M.; Rayner, J.M.V. Ecological morphology and flight in bats (Mammalia; Chiroptera): Wing adaptations, flight performance, foraging strategy and echolocation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1987, 316, 335–427. [Google Scholar]
- Neuweiler, G. Foraging ecology and audition in echolocating bats. Trends Ecol. Evol. 1989, 4, 160–166. [Google Scholar] [CrossRef]
- Fenton, M.B. The foraging behaviour and ecology of animal-eating bats. Can. J. Zool. 1990, 68, 411–422. [Google Scholar] [CrossRef]
- Hughes, P.; Rayner, J.; Jones, G. Ontogeny of ‘true’ flight and other aspects of growth in the bat Pipistrellus pipistrellus. J. Zool. 1995, 236, 291–318. [Google Scholar] [CrossRef]
- Brunet-Rossinni, A.K.; Wilkinson, G.S. Methods for age estimation and the study of senescence in bats. In Ecological and Behavioral Methods for the Study of Bats, 2nd ed.; Kunz, T.H., Parsons, S., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2009; pp. 315.–325. [Google Scholar]
- Kunz, T.H.; Hood, W.R. Parental care and postnatal growth in the Chiroptera. In Reproductive Biology of Bats; Crichton, E.G., Krutzsch, P.H., Eds.; Elsevier: Amsterdam, The Netherlands; San Diego Academic Press: San Diego, CA, USA, 2000; pp. 415–468. [Google Scholar]
- Adams, R.A. Morphogenesis in bat wings: Linking development, evolution and ecology. Cells Tissues Organs 2008, 187, 13–23. [Google Scholar] [CrossRef]
- Jones, C. Growth, development, and wing loading in the evening bat, Nycticeius humeralis (Rafinesque). J. Mammal. 1967, 48, 1–19. [Google Scholar] [CrossRef]
- Davis, R. Growth and development of young pallid bats, Antrozous pallidus. J. Mammal. 1969, 50, 729–736. [Google Scholar] [CrossRef]
- Kunz, T.H.; Robson, S.K. Postnatal growth and development in the Mexican free-tailed bat (Tadarida brasiliensis mexicana): Birth size, growth rates, and age estimation. J. Mammal. 1995, 76, 769–783. [Google Scholar] [CrossRef]
- Isaac, S.S.; Marimuthu, G. Development of wing morphology in the Indian pygmy bat Pipistrellus mimus. J. Biosci. 1997, 22, 193–202. [Google Scholar] [CrossRef]
- Elangovan, V.; Raghuram, H.; Satya Priya, E.Y.; Marimuthu, G. Wing morphology and flight performance in Rousettus leschenaulti. J. Mammal. 2004, 85, 806–812. [Google Scholar] [CrossRef] [Green Version]
- Elangovan, V.; Priya, E.Y.S.; Raghuram, H.; Marimuthu, G. Wing morphology and flight development in the short-nosed fruit bat Cynopterus sphinx. Zoology 2007, 110, 189–196. [Google Scholar] [CrossRef]
- Reiter, G. Postnatal growth and reproductive biology of Rhinolophus hipposideros (Chiroptera: Rhinolophidae). J. Zool. 2004, 262, 231–241. [Google Scholar] [CrossRef]
- Sharifi, M. Postnatal growth and age estimation in the Mehely’s horseshoe bat (Rhinolophus mehelyi). Acta Chiropterol. 2004, 6, 155–161. [Google Scholar] [CrossRef]
- Jin, L.; Lin, A.; Sun, K.; Liu, Y.; Feng, J. Postnatal growth and age estimation in the ashy leaf-nosed bat, Hipposideros cineraceus. Acta Chiropterol. 2010, 12, 155–160. [Google Scholar] [CrossRef]
- Lin, A.; Jin, L.; Liu, Y.; Sun, K.; Feng, J. Postnatal growth and age estimation in Horsfield’s leaf-nosed bat Hipposideros larvatus. Zool. Stud. 2010, 49, 789–796. [Google Scholar]
- Aldridge, H.D. Turning flight of bats. J. Exp. Biol. 1987, 128, 419. [Google Scholar] [PubMed]
- Aldridge, H.D.J.N.; Brigham, R.M. Load carrying and maneuverability in an insectivorous bat: A test of the 5% “rule” of radio-telemetry. J. Mammal. 1988, 69, 379–382. [Google Scholar] [CrossRef]
- Huges, P.M.; Rayner, J.M.V. Addition of artificial loads to long-eared bats Plecotus auritus: Handicapping flight performance. J. Exp. Biol. 1991, 161, 285–298. [Google Scholar]
- Strauss, M.; Knörnschild, M.; von Helversen, O. The ontogeny of courtship behaviours in bat pups (Saccopteryx bilineata). Behaviour 2010, 147, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Gould, E. Studies of maternal-infant communication and development of vocalizations in the bats Myotis and Eptesicus. Commun. Behav. Biol. 1971, 5, 263–313. [Google Scholar]
- Kunz, T.H. Population studies of the cave bat (Myotis velifer): Reproduction, growth and development. Counc. Nat. Acad. Award. 1973, 15, 1–43. [Google Scholar] [CrossRef]
- Yokoyama, K.; Ohtsu, R.; Uchida, T.A. Growth and LDH isozyme patterns in the pectoral and cardiac muscles of the Japanese Lesser horseshoe bat, Rhinolophus cornutus cornutus from the standpoint of adaptation for flight. J. Zool. 1979, 187, 85–96. [Google Scholar] [CrossRef]
- Yokoyama, K.; Uchida, T. Functional morphology of wings from the standpoint of adaptation for flight in chiroptera: II. growth and changes in mode of life during the young period in Rhinolophus Cornutus Cornutus. J. Fac. Agric. Kyushu Univ. 1979, 23, 185–198. [Google Scholar]
- Kleiman, D.G. Maternal care, growth rate, and development in the noctule (Nyctalus noctula), pipistrelle (Pipistrellus pipistrellus), and serotine (Eptesicus serotinus) bats. J. Zool. 1969, 157, 187–211. [Google Scholar] [CrossRef]
- Powers, L.V.; Kandarian, S.C.; Kunz, T.H. Ontogeny of flight in the little brown bat, Myotis lucifugus: Behavior, morphology, and muscle histochemistry. J. Comp. Physiol. A 1991, 168, 675–685. [Google Scholar] [CrossRef]
- Yokoyama, K.; Uchida, T.; Shiraishi, S. Functional morphology of wings from the standpoint of adaptation for flight in Chiroptera.I. Relative growth and ossification in forelimb, wing loading and aspect ratio. Zool. Mag. 1975, 84, 233–247. [Google Scholar]
- Jin, L.R.; Wang, J.; Zhang, Z.Z.; Sun, K.P.; Kanwal, J.S.; Feng, J. Postnatal development of morphological and vocal features in Asian parti-colored bat, Vespertilio sinensis. Mamm. Biol. 2012, 77, 339–344. [Google Scholar] [CrossRef]
- Peig, J.; Green, A.J. New perspectives for estimating body condition from mass/length data: The scaled mass index as an alternative method. Oikos 2009, 118, 1883–1891. [Google Scholar] [CrossRef]
- Maina, J.N. What it takes to fly: The structural and functional respiratory refinements in birds and bats. J. Exp. Biol. 2000, 203, 3045–3064. [Google Scholar]
- Bell, E.; Price, E.; Balthes, S.; Cordon, M.; Wormell, D. Flight patterns in zoo-housed fruit bats (Pteropus spp.). Zoo Biol. 2019, 38, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Harrison, X.A. Using observation-level random effects to model overdispersion in count data in ecology and evolution. PeerJ 2014, 2, e616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppenheim, R.W. Ontogenetic adaptations and retrogressive processes in the development of the nervous system and behavior: A neuroembryological perspective. In Maturation and Development: Biological and Psychological Perspectives; Cambridge University Press: Cambridge, UK, 1981; pp. 73–109. [Google Scholar]
- Patrick, P.G.B. Ontogeny of behaviour. Br. Med. Bull. 1981, 37, 159–164. [Google Scholar]
- Bolhuis, J.J. The development of animal behavior: From lorenz to neural nets. Naturwissenschaften 1999, 86, 101–111. [Google Scholar] [CrossRef]
- Fox, W.M. Reflex-ontogeny and behavioral development of the mouse. Anim Behav. 1965, 13, 234–241. [Google Scholar] [CrossRef]
- Graham, K.; Burghardt, G. Current perspectives on the biological study of play: Signs of progress. Q. Rev. Biol. 2010, 85, 393–418. [Google Scholar] [CrossRef] [Green Version]
- Bracke, M.B.M.; Hopster, H. Assessing the importance of natural behavior for animal welfare. J. Agric. Environ. Ethics 2006, 19, 77–89. [Google Scholar] [CrossRef]
- Heers, A.M.; Dial, K.P. Wings versus legs in the avian bauplan: Development and evolution of alternative locomotor strategies. Evolution 2014, 69, 305–320. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.A.; Snode, E.R.; Shaw, J.B.; Fenton, B. Flapping tail membrane in bats produces potentially important thrust during horizontal takeoffs and very slow flight. PLoS ONE 2012, 7, e32074. [Google Scholar] [CrossRef] [Green Version]
- Winter, Y.; von Helvesen, O. The energy cost of flight: Do small bats fly more cheaply than small birds? J. Comp. Physiol. B 1998, 168, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Spinka, M.; Newberry, R.C.; Bekoff, M. Mammalian play: Training for the unexpected. Q. Rev. Biol. 2001, 76, 141–168. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, G.M. The Genesis of Animal Play: Testing the Limits; Mit Press: Cambrige, MA, USA, 2005. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behaviour | Standard | Description |
|---|---|---|
| Flight | Success | Young bats take the initiative to leave the flight platform and fly smoothly through the air. |
| Failure | Young bats stay on the flight platform or fall off it uncontrolled. | |
| Landing | Success | Young bats stabilize their limbs and body immediately before landing in a smooth and confident movement. |
| Failure | Young bats land uncontrolled, usually accompanied by extending their patagium or making an additional flapping motion. |
| Type | Number of Occurrences | Percentage | Description |
|---|---|---|---|
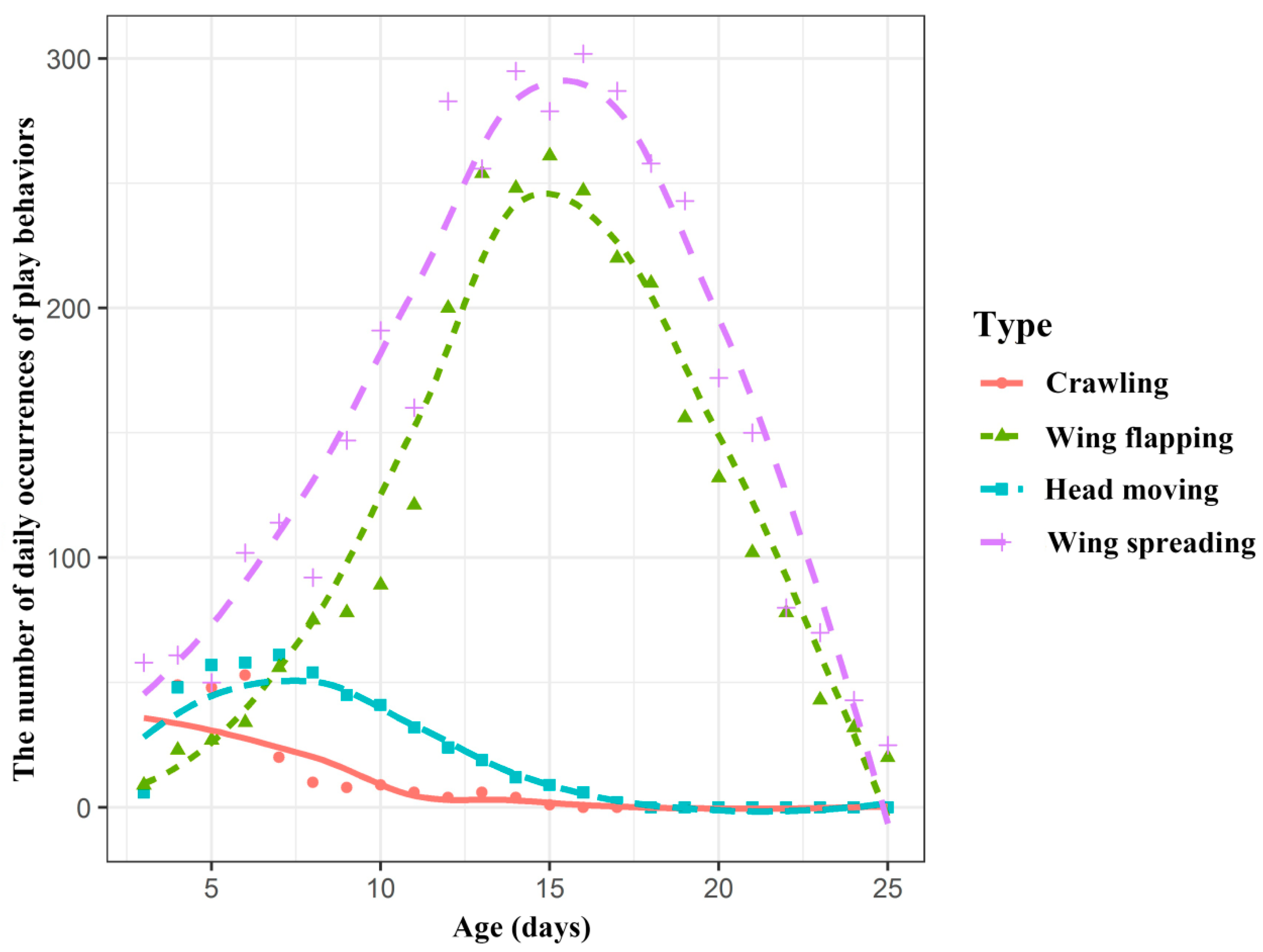
| Wing flapping | 2715 | 38% | Infants open one or both wings and then flap them at a high frequency. The flapping of the wings occurs continuously, with intervals between each flap. This is occasionally accompanied by head movements. |
| Wing spreading | 3718 | 52% | Infants open one or both wings without flapping, sometimes opening and closing the wings and sometimes leaving them open for a period of time. When they spread their wings, the infants are usually not performing other behaviours. |
| Crawling | 225 | 3% | Infants move away from the other individuals and crawl around the cage. This behaviour would happen when the female had left to find food. The infants did not stray very far from their starting point, and then they returned to the group. |
| Head moving | 474 | 7% | Infants push their heads up and look around. The range of movement frequency during head moving play was much wider than that of adults when perceiving their surroundings. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, D.; Li, Y.; Yin, Z.; Zhang, K.; Liu, H.; Liu, Y.; Feng, J. Behavioural Patterns and Postnatal Development in Pups of the Asian Parti-Coloured Bat, Vespertilio sinensis. Animals 2020, 10, 1325. https://doi.org/10.3390/ani10081325
Sun D, Li Y, Yin Z, Zhang K, Liu H, Liu Y, Feng J. Behavioural Patterns and Postnatal Development in Pups of the Asian Parti-Coloured Bat, Vespertilio sinensis. Animals. 2020; 10(8):1325. https://doi.org/10.3390/ani10081325
Chicago/Turabian StyleSun, Deyi, Yu Li, Zhongwei Yin, Kangkang Zhang, Heng Liu, Ying Liu, and Jiang Feng. 2020. "Behavioural Patterns and Postnatal Development in Pups of the Asian Parti-Coloured Bat, Vespertilio sinensis" Animals 10, no. 8: 1325. https://doi.org/10.3390/ani10081325
APA StyleSun, D., Li, Y., Yin, Z., Zhang, K., Liu, H., Liu, Y., & Feng, J. (2020). Behavioural Patterns and Postnatal Development in Pups of the Asian Parti-Coloured Bat, Vespertilio sinensis. Animals, 10(8), 1325. https://doi.org/10.3390/ani10081325