Hydrolyzed Yeast Supplementation in Calf Starter Promotes Innate Immune Responses in Holstein Calves under Weaning Stress Condition

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Weaning and Blood Sampling
2.3. BW Gain and Feed Intake
2.4. General Health Monitoring
2.5. Hematology
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Flow Cytometry
2.8. Statistical Analysis
3. Results
3.1. Growth Performance and Feed Intake
3.2. Stress Hormone
3.3. Pro-Inflammatory Cytokines
3.4. Acute-Phase Proteins
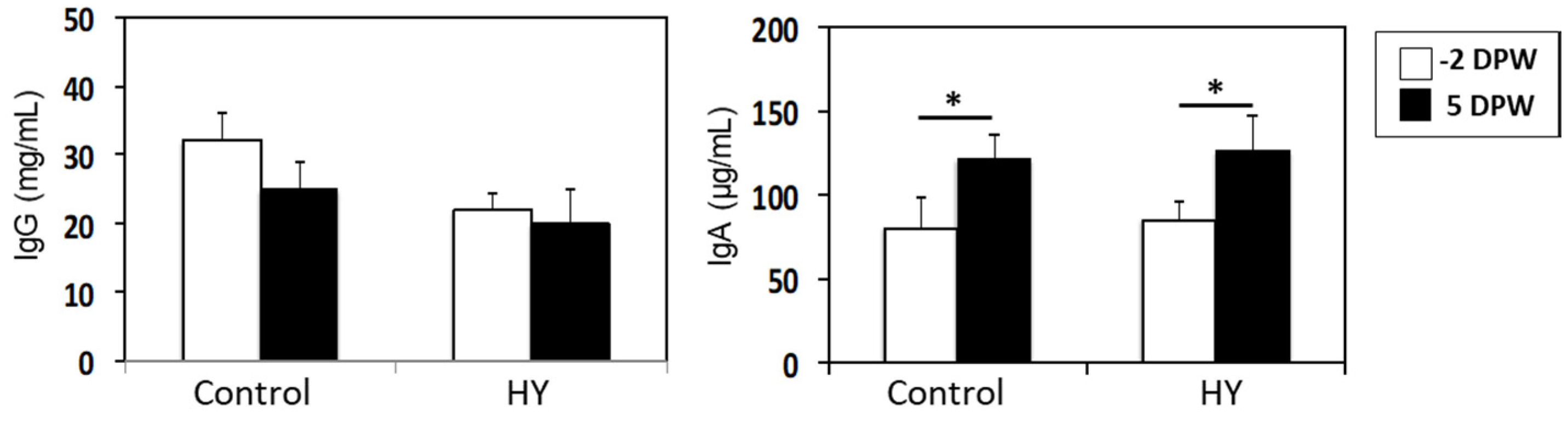
3.5. Immunoglobulins
3.6. Composition of PBMC
3.7. Health Status
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bach, A.; Ahedo, J. Record keeping and economics of dairy heifers. Vet. Clin. North. Am. Food Anim. Pract. 2008, 24, 117–138. [Google Scholar] [CrossRef] [PubMed]
- Chester-Jones, H.; Heins, B.; Ziegler, D.; Schimek, D.; Schuling, S.; Ziegler, B.; De Ondarza, M.; Sniffen, C.; Broadwater, N. Relationships between early-life growth, intake, and birth season with first-lactation performance of Holstein dairy cows. J. Dairy Sci. 2017, 100, 3697–3704. [Google Scholar] [CrossRef] [PubMed]
- Hulbert, L.E.; Moisá, S.J. Stress, immunity, and the management of calves. J. Dairy Sci. 2016, 99, 3199–3216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickey, M.C.; Drennan, M.; Earley, B. The effect of abrupt weaning of suckler calves on the plasma concentrations of cortisol, catecholamines, leukocytes, acute-phase proteins and in vitro interferon-gamma production. J. Anim. Sci. 2003, 81, 2847–2855. [Google Scholar] [CrossRef] [PubMed]
- Godbout, J.P.; Glaser, R. Stress-induced immune dysregulation: Implications for wound healing, infectious disease and cancer. J. Neuroimmune Pharmacol. 2006, 1, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Foote, M.; Nonnecke, B.; Beitz, D.; Waters, W. Antigen-specific B-cell responses by neonatal calves after early vaccination. J. Dairy Sci. 2007, 90, 5208–5217. [Google Scholar] [CrossRef]
- Elenkov, I.J. Glucocorticoids and the Th1/Th2 balance. Ann. Acad. Sci. 2004, 1024, 138–146. [Google Scholar] [CrossRef]
- Li, L.A.; Yang, J.J.; Li, Y.; Lv, L.; Xie, J.J.; Du, G.M.; Jin, T.M.; Qin, S.Y.; Jiao, X.L. Effect of weaning age on cortisol release in piglets. Genet. Mol. Res. 2016, 15, 10. [Google Scholar] [CrossRef]
- Foote, A.P.; Jones, S.A.; Kuehn, L.A. Association of preweaning and weaning serum cortisol and metabolites with ADG and incidence of respiratory disease in beef cattle. J. Anim. Sci. 2017, 95, 5012–5019. [Google Scholar] [CrossRef]
- Carroll, J.A.; Arthington, J.D.; Chase, C.C., Jr. Early weaning alters the acute-phase reaction to an endotoxin challenge in beef calves. J. Anim. Sci. 2009, 87, 4167–4172. [Google Scholar] [CrossRef] [Green Version]
- Lynch, E.M.; Earley, B.; McGee, M.; Doyle, S. Effect of abrupt weaning at housing on leukocyte distribution, functional activity of neutrophils, and acute phase protein response of beef calves. BMC Vet. Res. 2010, 6, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.I.; Yoon, K.J. An overview of calf diarrhea–infectious etiology, diagnosis, and intervention. J. Vet. Sci. 2014, 15, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thrune, M.; Bach, A.; Ruiz-Moreno, M.; Stern, M.; Linn, J. Effects of Saccharomyces cerevisiae on ruminal pH and microbial fermentation in dairy cows: Yeast supplementation on rumen fermentation. Livest. Sci. 2009, 124, 261–265. [Google Scholar] [CrossRef]
- Davis, M.; Maxwell, C.; Erf, G.; Brown, D.; Wistuba, T. Dietary supplementation with phosphorylated mannans improves growth response and modulates immune function of weanling pigs. J. Anim. Sci. 2004, 82, 1882–1891. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Guo, Y.; Yuan, J.; Zhang, B. Effect of Dietary β-1, 3/1, 6-glucan Supplementation on Growth Performance, Immune Response and Plasma Prostaglandin E2, Growth Hormone and Ghrelin in Weanling Piglets. Asian-Australas J. Anim Sci. 2008, 21, 707–714. [Google Scholar] [CrossRef]
- Jensen, G.S.; Patterson, K.M.; Yoon, I. Yeast culture has anti-inflammatory effects and specifically activates NK cells. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 487–500. [Google Scholar] [CrossRef]
- Pedroso, M. Application of beta-1,3-glucan to prevent shipping fever in imported heifers. Arch. Med. Res. 1994, 25, 181–184. [Google Scholar]
- Lowry, V.K.; Farnell, M.B.; Ferro, P.J.; Swaggerty, C.L.; Bahl, A.; Kogut, M.H. Purified β-glucan as an antibiotic feed additive up-regulates the innate immune response in immature chickens against Salmonella enterica serovar Enteritidis. Int. J. Food Microbiol. 2005, 98, 309–318. [Google Scholar] [CrossRef]
- Rodriguez, A.; Cuesta, A.; Ortuño, J.; Esteban, M.A.; Meseguer, J. Immunostimulant properties of a cell wall-modified whole Saccharomyces cerevisiae strain administered by diet to seabream (Sparus aurata L.). Vet. Immunol. Immunopathol. 2003, 96, 183–192. [Google Scholar] [CrossRef]
- Khan, M.; Lee, H.; Lee, W.; Kim, H.; Kim, S.; Ki, K.; Ha, J.; Lee, H.; Choi, Y. Pre-and postweaning performance of Holstein female calves fed milk through step-down and conventional methods. J. Dairy Sci. 2007, 90, 876–885. [Google Scholar] [CrossRef]
- AOAC. Official methods of analysis of AOAC International. 15th ed. Arlington (VA): AOAC International. Assoc. Anal. Chem 1990, 1, 771. [Google Scholar]
- Stoldt, W. Vorschlag zur vereinheitlichung der fettbestimmung in lebensmitteln. Fette Seifen 1952, 54, 206–207. [Google Scholar] [CrossRef]
- Larson, L.; Owen, F.; Albright, J.; Appleman, R.; Lamb, R.; Muller, L. Guidelines toward more uniformity in measuring and reporting calf experimental data. J. Dairy Sci. 1977, 60, 989–991. [Google Scholar] [CrossRef]
- Bate, S.T.; Clark, R.A. The Design and Statistical Analysis of Animal Experiments; Cambridge University Press: Cambridge, London, UK, 2014; p. 324. [Google Scholar]
- Institute, S. Statistical Analysis Software (SAS) User’s Guide, Version 9.2; SAS Publishing: Cary, NC, USA, 2001; p. 176. [Google Scholar]
- Wagner, D.; Quinonez, J.; Bush, L. The effect of corn-or wheat-based diets and yeast culture on performance, ruminal pH, and volatile fatty acids in dairy calves. Agri Pract. 1990, 11, 7–12. [Google Scholar]
- Quigley, J.; Wallis, L.; Dowlen, H.; Heitmann, R. Sodium bicarbonate and yeast culture effects on ruminal fermentation, growth, and intake in dairy calves. J. Dairy Sci. 1992, 75, 3531–3538. [Google Scholar] [CrossRef]
- Kim, M.; Seo, J.; Yun, C.; Kang, S.; Ko, J.; Ha, J. Effects of hydrolyzed yeast supplementation in calf starter on immune responses to vaccine challenge in neonatal calves. Animal 2011, 5, 953–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callaway, E.; Martin, S. Effects of a Saccharomyces cerevisiae culture on ruminal bacteria that utilize lactate and digest cellulose. J. Dairy Sci. 1997, 80, 2035–2044. [Google Scholar] [CrossRef]
- Kumar, U.; Sareen, V.K.; Singh, S. Effect of yeast culture supplement on ruminal microbial populations and metabolism in buffalo calves fed a high roughage diet. J. Sci. Food Agric. 1997, 73, 231–236. [Google Scholar] [CrossRef]
- Dann, H.; Drackley, J.K.; McCoy, G.; Hutjens, M.; Garrett, J. Effects of yeast culture (Saccharomyces cerevisiae) on prepartum intake and postpartum intake and milk production of Jersey cows. J. Dairy Sci. 2000, 83, 123–127. [Google Scholar] [CrossRef]
- Seymour, W.; Nocek, J.; Siciliano-Jones, J. Effects of a colostrum substitute and of dietary brewer’s yeast on the health and performance of dairy calves. J. Dairy Sci. 1995, 78, 412–420. [Google Scholar] [CrossRef]
- Lesmeister, K.; Heinrichs, A.J.; Gabler, M. Effects of supplemental yeast (Saccharomyces cerevisiae) culture on rumen development, growth characteristics, and blood parameters in neonatal dairy calves. J. Dairy Sci. 2004, 87, 1832–1839. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, A.; Kholif, A.; Lugo-Coyote, R.; Elghandour, M.; Cipriano, M.; Rodríguez, G.; Odongo, N.; Salem, A. The effect of garlic oil, xylanase enzyme and yeast on biomethane and carbon dioxide production from 60-d old Holstein dairy calves fed a high concentrate diet. J. Clean. Prod. 2017, 142, 2384–2392. [Google Scholar] [CrossRef]
- Harrison, G.; Hemken, R.; Dawson, K.; Harmon, R.; Barker, K. Influence of addition of yeast culture supplement to diets of lactating cows on ruminal fermentation and microbial populations 1. J. Dairy Sci. 1988, 71, 2967–2975. [Google Scholar] [CrossRef]
- Roger, V.; Fonty, G.; Komisarczuk-Bony, S.; Gouet, P. Effects of physicochemical factors on the adhesion to cellulose avicel of the ruminal bacteria Ruminococcus flavefaciens and Fibrobacter succinogenes subsp. succinogenes. Appl. Environ. Microbiol. 1990, 56, 3081–3087. [Google Scholar] [CrossRef] [Green Version]
- Chaucheyras-Durand, F.; Walker, N.; Bach, A. Effects of active dry yeasts on the rumen microbial ecosystem: Past, present and future. Anim. Feed Sci. Technol. 2008, 145, 5–26. [Google Scholar] [CrossRef]
- Weary, D.M.; Jasper, J.; Hötzel, M.J. Understanding weaning distress. Appl. Anim. Behav. Sci. 2008, 110, 24–41. [Google Scholar] [CrossRef]
- Klis, F.M.; Mol, P.; Hellingwerf, K.; Brul, S. Dynamics of cell wall structure in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2002, 26, 239–256. [Google Scholar] [CrossRef]
- Rop, O.; Mlcek, J.; Jurikova, T. Beta-glucans in higher fungi and their health effects. Nutr. Rev. 2009, 67, 624–631. [Google Scholar] [CrossRef]
- Galvão, K.N.; Santos, J.E.; Coscioni, A.; Villaseñor, M.; Sischo, W.M.; Berge, A.C.B. Effect of feeding live yeast products to calves with failure of passive transfer on performance and patterns of antibiotic resistance in fecal Escherichia coli. Reprod. Nutr. Dev. 2005, 45, 427–440. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, V.; Susca, F.; Lima, F.; Branco, A.; Yoon, I.; Santos, J. Effect of feeding yeast culture on performance, health, and immunocompetence of dairy calves. J. Dairy Sci. 2008, 91, 1497–1509. [Google Scholar] [CrossRef] [Green Version]
- Seljelid, R.; Rasmussen, L.T.; Larm, O.; Hoffman, J. The Protective Effect of β1–3D-Glucan-Derivatized Plastic Beads against Escherichia coli Infection in Mice. Scand. J. Immunol. 1987, 25, 55–60. [Google Scholar] [CrossRef]
- Djeraba, A.; Quere, P. In vivo macrophage activation in chickens with Acemannan, a complex carbohydrate extracted from Aloe vera. Int. J. Immunopharmacol. 2000, 22, 365–372. [Google Scholar] [CrossRef]
- Shurson, G.C. Yeast and yeast derivatives in feed additives and ingredients: Sources, characteristics, animal responses, and quantification methods. Anim. Feed Sci. Technol. 2018, 235, 60–76. [Google Scholar] [CrossRef]
- Pérez-Sotelo, L.S.; Talavera-Rojas, M.; Monroy-Salazar, H.G.; Lagunas-Bernabé, S.; Cuarón-Ibargüengoytia, J.A.; Jimenez, R.; Vázquez-Chagoyán, J.C. In vitro evaluation of the binding capacity of Saccharomyces cerevisiae Sc47 to adhere to the wall of Salmonella spp. Rev. Lat. Microbiol. 2005, 47, 70–75. [Google Scholar]
- Beerda, B.; Schilder, M.B.; Janssen, N.S.; Mol, J.A. The use of saliva cortisol, urinary cortisol, and catecholamine measurements for a noninvasive assessment of stress responses in dogs. Horm. Behav. 1996, 30, 272–279. [Google Scholar] [CrossRef]
- Queyras, A.; Carosi, M. Non-invasive techniques for analysing hormonal indicators of stress. Ann. Ist. Super Sanita 2004, 40, 211–221. [Google Scholar] [PubMed]
- Warren, E.; Finck, B.; Arkins, S.; Kelley, K.; Scamurra, R.; Murtaugh, M.P.; Johnson, R.W. Coincidental changes in behavior and plasma cortisol in unrestrained pigs after intracerebroventricular injection of tumor necrosis factor-α. Endocrinology 1997, 138, 2365–2371. [Google Scholar] [CrossRef]
- Heegaard, P.M.; Godson, D.L.; Toussaint, M.J.; Tjørnehøj, K.; Larsen, L.E.; Viuff, B.; Rønsholt, L. The acute phase response of haptoglobin and serum amyloid A (SAA) in cattle undergoing experimental infection with bovine respiratory syncytial virus. Vet. Immunol. Immunopathol. 2000, 77, 151–159. [Google Scholar] [CrossRef]
- Murata, H. Stress and acute phase protein response: An inconspicuous but essential linkage. Vet. J. 2007, 173, 473. [Google Scholar] [CrossRef] [PubMed]
- Roza, A.M.; Tuitt, D.; Shizgal, H.M. Transferrin—A poor measure of nutritional status. J. Parenter Enter. Nutr. 1984, 8, 523–528. [Google Scholar] [CrossRef]
- Moser, M.; Pfister, H.; Bruckmaier, R.; Rehage, J.; Blum, J. Blood Serum Transferrin Concentration in Cattle in Various Physiological States, in Veal Calves Fed Different Amounts of Iron, and in Cattle Affected by Infectious and Non-Infectious Diseases. Zentrabl. Vet. A 1994, 41, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.Y.; Cisalpino, D.; Varadarajan, S.; Hellman, J.; Warren, H.S.; Cascalho, M.; Inohara, N.; Núñez, G. Gut microbiota-induced immunoglobulin G controls systemic infection by symbiotic bacteria and pathogens. Immunity 2016, 44, 647–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Church, J.; Hudson, R. Comparison of the stress of abrupt and interval weaning of farmed wapiti calves (Cervus elaphus). Small Rumin. Res. 1999, 32, 119–124. [Google Scholar] [CrossRef]
- Kang, S.Y.; Ko, Y.H.; Moon, Y.S.; Sohn, S.H.; Jang, I.S. Effects of the combined stress induced by stocking density and feed restriction on hematological and cytokine parameters as stress indicators in laying hens. Asian Australas J. Anim. Sci 2011, 24, 414–420. [Google Scholar] [CrossRef]
- Anderson, B.; Watson, D.; Colditz, I. The effect of dexamethasone on some immunological parameters in cattle. Vet. Res. Commun. 1999, 23, 399–413. [Google Scholar] [CrossRef]
- Shen, Y.; Piao, X.; Kim, S.; Wang, L.; Liu, P.; Yoon, I.; Zhen, Y. Effects of yeast culture supplementation on growth performance, intestinal health, and immune response of nursery pigs. J. Anim. Sci. 2009, 87, 2614–2624. [Google Scholar] [CrossRef]
- Nonnecke, B.; Foote, M.; Smith, J.; Pesch, B.; Van Amburgh, M. Composition and functional capacity of blood mononuclear leukocyte populations from neonatal calves on standard and intensified milk replacer diets. J. Dairy Sci. 2003, 86, 3592–3604. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (% of DM) | CON | HY |
|---|---|---|
| Ground corn | 19.98 | 19.98 |
| Ground wheat | 14.30 | 14.30 |
| Molasses | 5.00 | 5.00 |
| Wheat hulls | 15.00 | 15.00 |
| Gluten feed | 10.00 | 10.00 |
| Soybean hulls | 7.00 | 7.00 |
| SBM | 15.43 | 15.23 |
| HY | − | 0.20 |
| Corn germ meal | 5.00 | 5.00 |
| Copra meal | 5.00 | 5.00 |
| Salt | 0.50 | 0.50 |
| Ca3(PO4)2 | 0.38 | 0.38 |
| CaCo3 | 1.50 | 1.50 |
| Bio–Zn | 0.01 | 0.01 |
| Virginiamycin | 0.10 | 0.10 |
| Premix (vitamins & trace minerals) 1 | 0.30 | 0.30 |
| NaHCo3 | 0.50 | 0.50 |
| Chemical composition (% of feed) | CON | HY |
| CP | 17.50 | 17.52 |
| CF | 2.68 | 2.66 |
| FAT | 2.55 | 2.57 |
| Ash | 6.44 | 6.44 |
| Calf Starter | |||
|---|---|---|---|
| Variables | CON | HY | p-Value |
| BW gain (kg) | |||
| Pre-weaning 1 | 11.58 ± 2.1 | 9.68 ± 1.3 | 0.11 |
| Post-weaning 2 | 4.11 ± 0.09 a | 4.74 ± 0.05 b | 0.003 |
| Calf starter intake (kg/week) | |||
| Pre-weaning | 2.59 ± 0.05 a | 2.96 ± 0.030 b | 0.001 |
| Post-weaning | 3.43 ± 0.05 | 3.31 ± 0.04 | 0.16 |
| Milk intake (L/week) | |||
| 2–3 weeks | 68.03 ± 1.4 | 71.80 ± 1.6 | 0.055 |
| 4–5 weeks | 48.30 ± 0.3 | 47.00 ± 0.8 | 0.61 |
| Total forage intake (g) | |||
| Pre-weaning | 510.41 ± 12.1 | 508.68 ± 10.3 | 0.66 |
| Post-weaning | 483.11 ± 13.5 | 447.35 ± 11.0 | 0.56 |
| Total DMI 3 (kg) | 47.08 ± 2.1 | 45.03 ± 1.1 | 0.47 |
| Feed efficiency 4 | 0.43 ± 0.02 | 0.44 ± 0.01 | 0.56 |
| Variables | Calf Starter | p-Value | |
|---|---|---|---|
| CON | HY | ||
| Diarrheal incidence 1 (%) | |||
| Pre-weaning | 11.11 ± 4.1 | 7.04 ± 3.1 | 0.22 |
| Post-weaning | 6.92 ± 2.2 | 3.18 ± 1.1 | 0.35 |
| Diarrhea severity 2 | |||
| Pre-weaning | 2.42 ± 0.7 | 2.33 ± 0.7 | 0.53 |
| Post-weaning | 2.51 ± 0.5 a | 1.09 ± 0.2 b | 0.04 |
| Health score 3 | |||
| Pre-weaning | 2.91 ± 1.3 | 2.61 ± 0.5 | 0.17 |
| Post-weaning | 2.76 ± 0.4 a | 1.11 ± 0.2 b | 0.03 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.T.; Lee, H.G.; Kim, D.H.; Son, J.K.; Kim, B.-W.; Joo, S.S.; Park, D.S.; Park, Y.J.; Lee, S.Y.; Kim, M.H. Hydrolyzed Yeast Supplementation in Calf Starter Promotes Innate Immune Responses in Holstein Calves under Weaning Stress Condition. Animals 2020, 10, 1468. https://doi.org/10.3390/ani10091468
Kim ET, Lee HG, Kim DH, Son JK, Kim B-W, Joo SS, Park DS, Park YJ, Lee SY, Kim MH. Hydrolyzed Yeast Supplementation in Calf Starter Promotes Innate Immune Responses in Holstein Calves under Weaning Stress Condition. Animals. 2020; 10(9):1468. https://doi.org/10.3390/ani10091468
Chicago/Turabian StyleKim, Eun Tae, Hyo Gun Lee, Dong Hyeon Kim, Jun Kyu Son, Byeong-Woo Kim, Sang Seok Joo, Da Som Park, Yei Ju Park, Se Young Lee, and Myung Hoo Kim. 2020. "Hydrolyzed Yeast Supplementation in Calf Starter Promotes Innate Immune Responses in Holstein Calves under Weaning Stress Condition" Animals 10, no. 9: 1468. https://doi.org/10.3390/ani10091468