Effects of RAC1 on Proliferation of Hen Ovarian Prehierarchical Follicle Granulosa Cells

 ,
, 
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chickens
2.2. Primary Culture of Chicken Ovarian GC
2.3. Quantitative Real-Time Polymerase and Chain Reaction (RT-qPCR)
2.4. Immunohistochemistry Assay
2.5. Construction of Recombinant Expression Vector and Cell Transfection
2.6. Western Blotting
2.7. Transfection of siRNA
2.8. Cell Proliferation Analysis
2.9. Statistical Analysis
3. Results
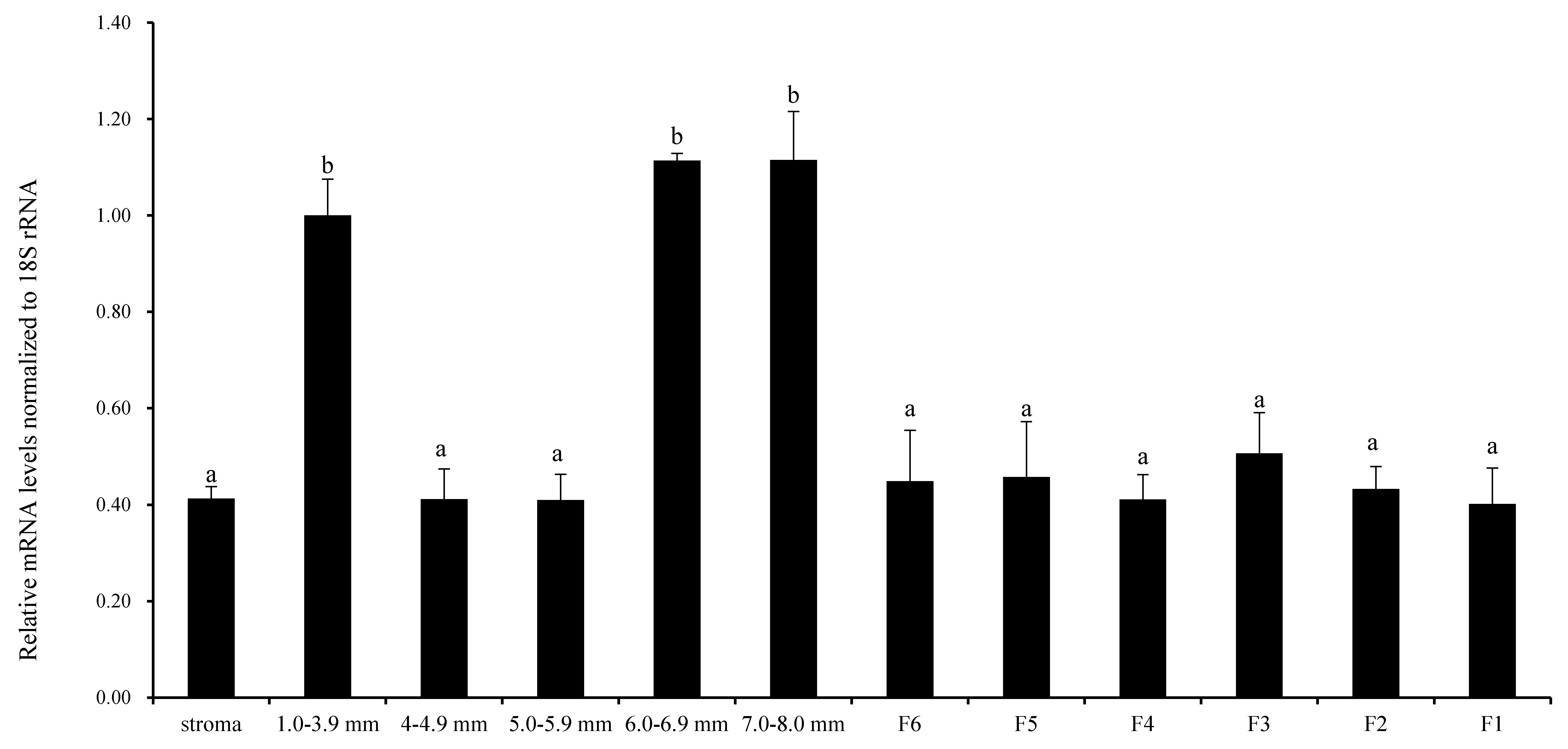
3.1. Expression of RAC1 in Chicken Ovarian Follicles
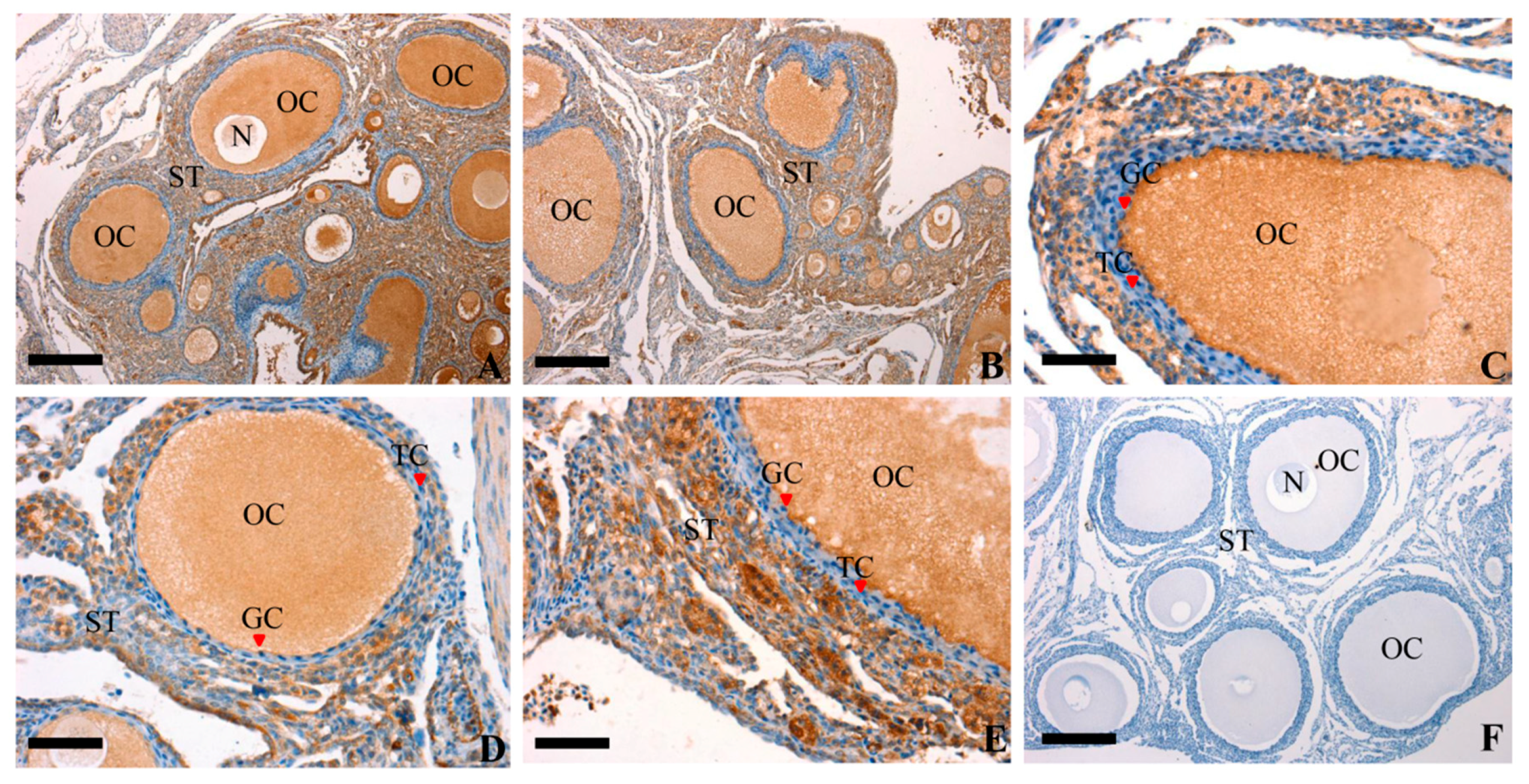
3.2. Localization of RAC1 in the Chicken Ovarian Follicles
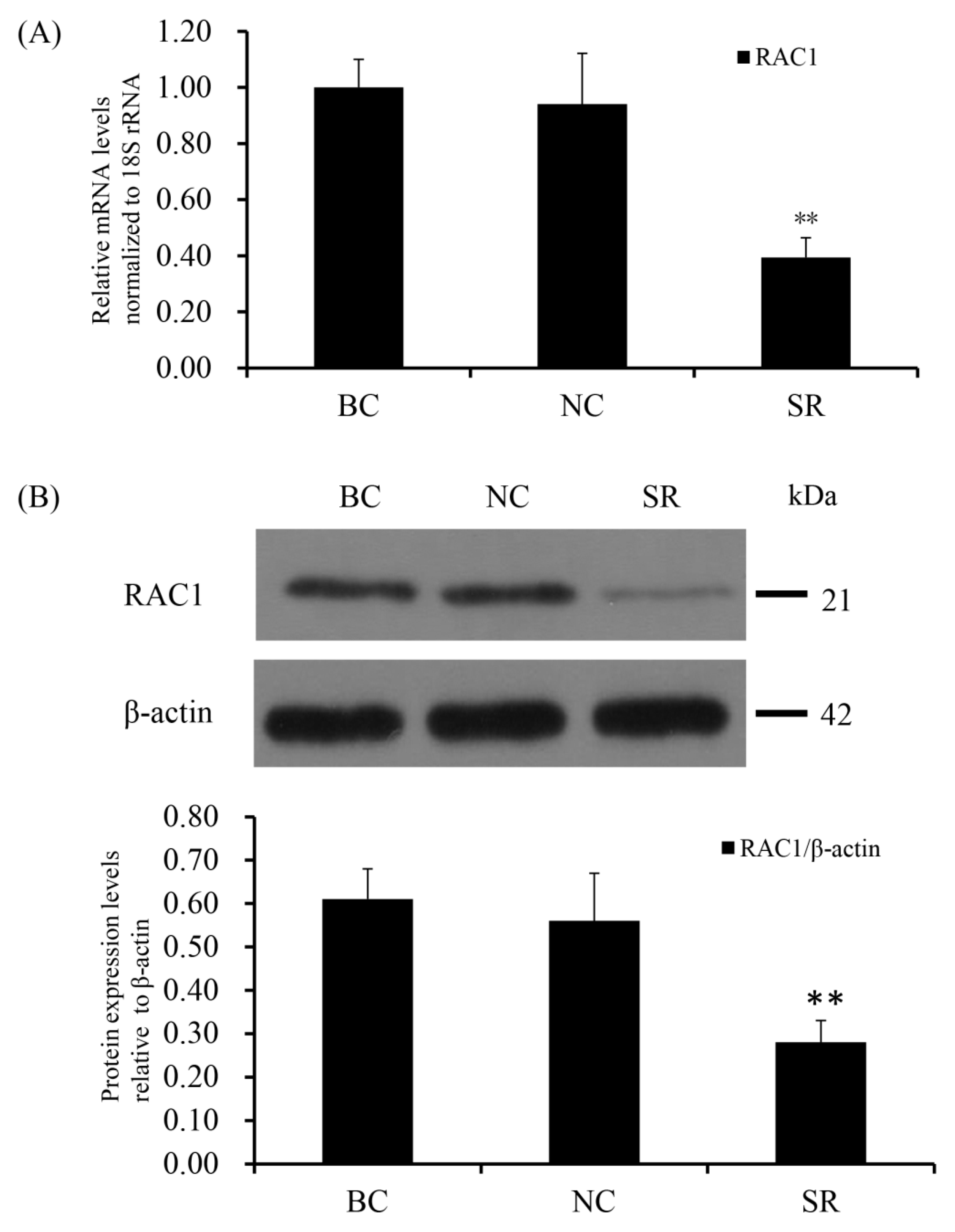
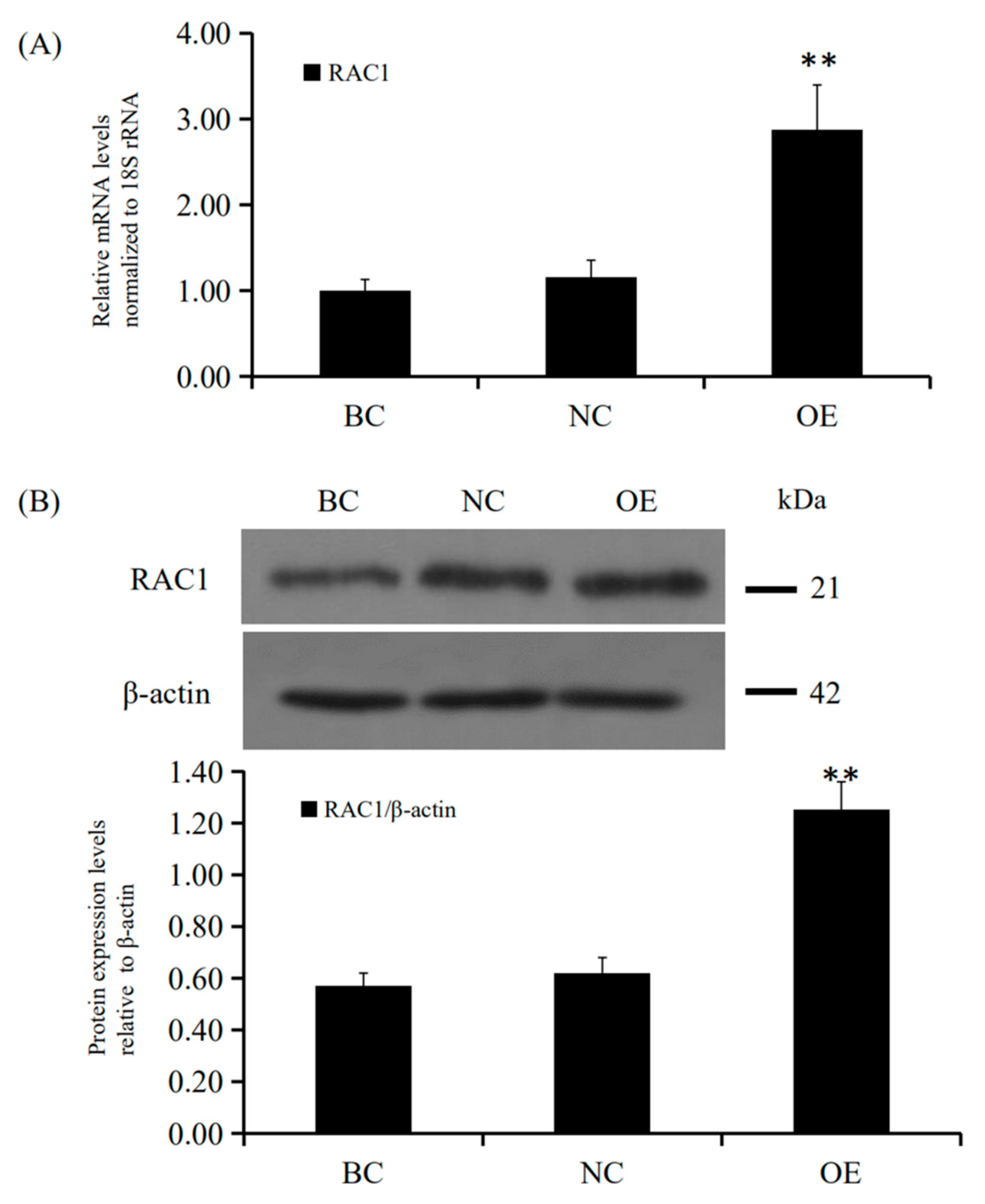
3.3. RAC1 Promoted the GCs Proliferation
3.4. Conformity of the Positive Effect of RAC1 on Granulosa Cell Proliferation
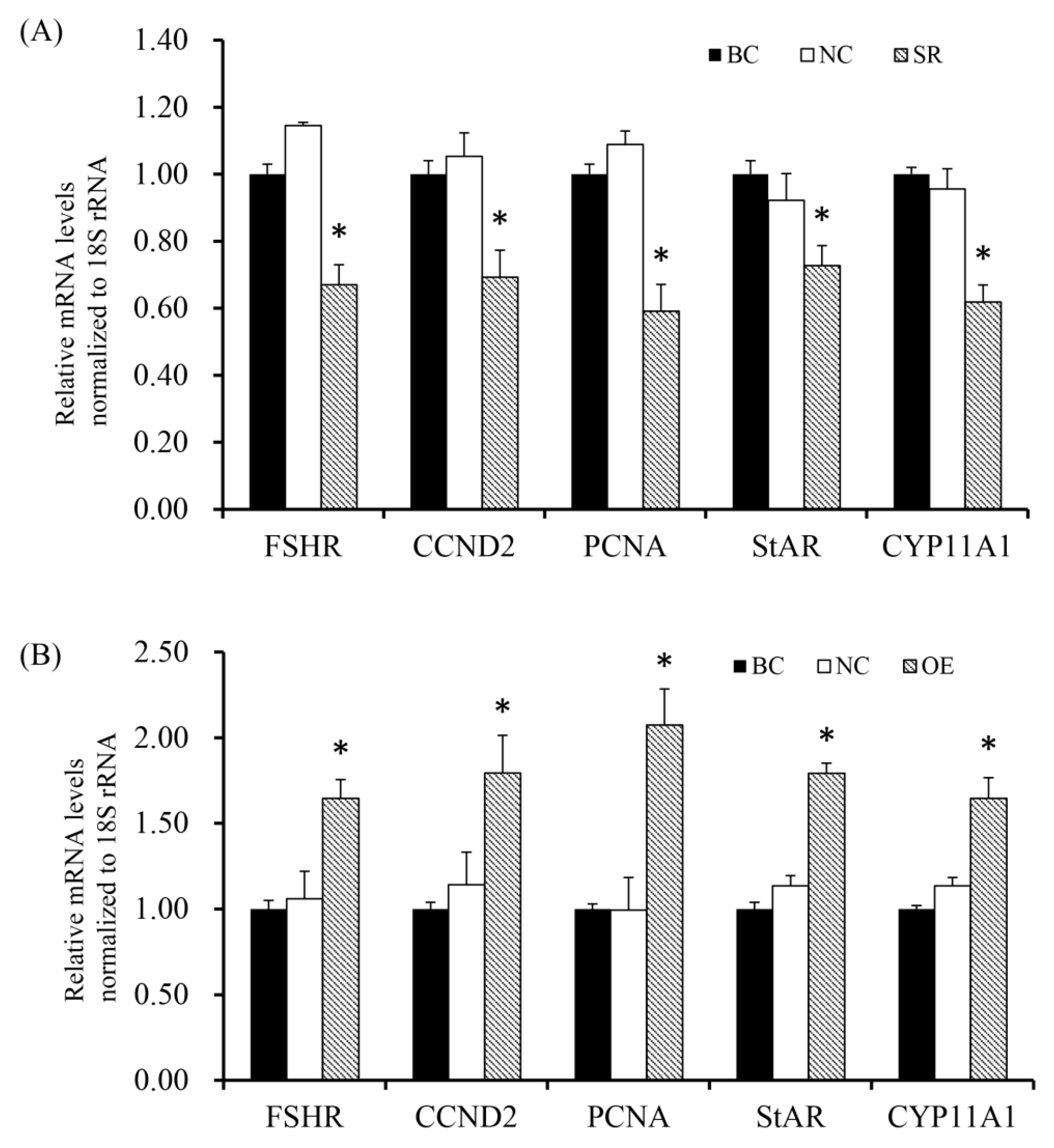
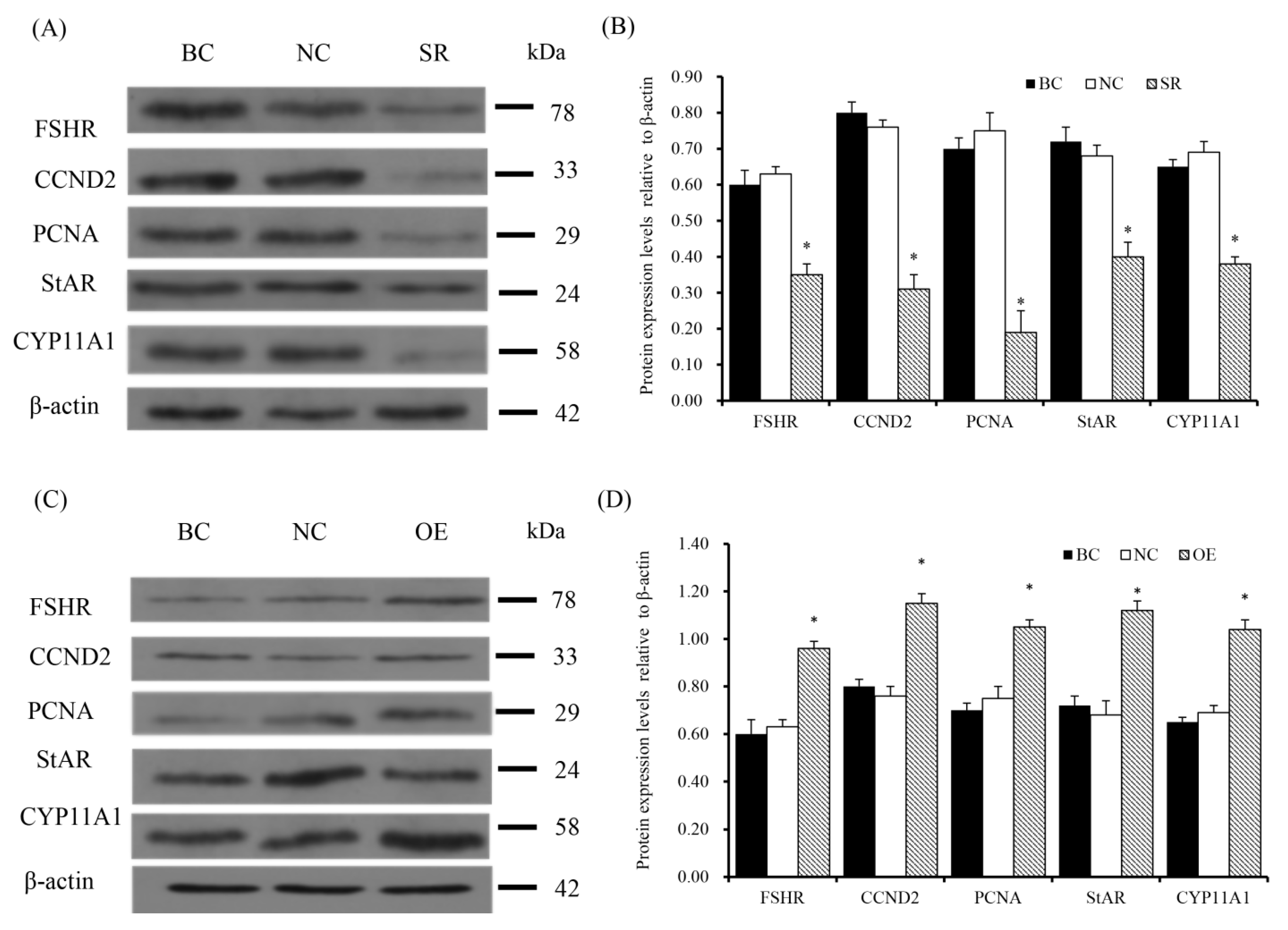
3.5. RAC1 Stimulated the mRNA and Protein Expression of FSHR, CCND2, PCNA, StAR and CYP11A1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhu, H.Y.; Qin, N.; Xu, X.X.; Sun, X.; Chen, X.X.; Zhao, J.H.; Xu, R.F.; Mishra, B. Synergistic inhibition of csal1 and csal3 in granulosa cell proliferation and steroidogenesis of hen ovarian prehierarchical development. Biol. Reprod. 2019, 101, 986–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, N.; Fan, X.C.; Xu, X.X.; Tyasi, T.L.; Li, S.J.; Zhang, Y.Y.; Wei, M.L.; Xu, R.F. Cooperative effects of FOXL2 with the members of TGF-β superfamily on FSH receptor mRNA expression and granulosa cell proliferation from hen prehierarchical follicles. PLoS ONE 2015, 10, e0141062. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.F.; Qin, N.; Xu, X.X.; Sun, X.; Chen, X.X.; Zhao, J.H. Inhibitory effect of SLIT2 on granulosa cell proliferation mediated by the CDC42-PAKs-ERK1/2 MAPK pathway in the prehierarchical follicles of the chicken ovary. Sci. Rep. 2018, 8, 9168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Vilchez, S.; Whitmore, L.; Asmussen, H.; Zareno, J.; Horwitz, R.; Newell-Litwa, K. RhoGTPase regulators orchestrate distinct stages of synaptic development. PLoS ONE 2017, 12, e0170464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo, A.; González-Billault, C. Crosstalk between Rac1-mediated actin regulation and ROS production. Free Radic. Biol. Med. 2018, 116, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.D.; Hao, T.; Gan, Y.H. RhoG/Rac1 signaling pathway involved in migration and invasion of salivary adenoid cystic carcinoma cells. Oral Dis. 2019, 26, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Castilho, R.M.; Squarize, C.H.; Leelahavanichkul, K.; Zheng, Y.; Bugge, T.; Gutkind, J.S. Rac1 is required for epithelial stem cell function during dermal and oral mucosal wound healing but not for tissue homeostasis in mice. PLoS ONE 2010, 5, e10503. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, N.; Nakao, M.; Matsui, S.; Satoh, T. A Crucial Role for the Small GTPase Rac1 Downstream of the Protein Kinase Akt2 in Insulin Signaling that Regulates Glucose Uptake in Mouse Adipocytes. Int. J. Mol. Sci. 2019, 20, 5443. [Google Scholar] [CrossRef] [Green Version]
- Halet, G.; Carroll, J. Rac activity is polarized and regulates meiotic spindle stability and anchoring in mammalian oocytes. Dev. Cell 2007, 12, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Grewal, S.; Carver, J.G.; Ridley, A.J.; Mardon, H.J. Implantation of the human embryo requires Rac1-dependent endometrial stromal cell migration. Proc. Natl. Acad. Sci. USA 2008, 105, 16189–16194. [Google Scholar] [CrossRef] [Green Version]
- Nicola, C.; Lala, P.K.; Chakraborty, C. Prostaglandin E2-mediated migration of human trophoblast requires RAC1 and CDC42. Biol. Reprod. 2008, 78, 976–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Liu, J.; Qi, Y.; Brakebusch, C.; Chrostek-Grashoff, A.; Edgar, D.; Yurchenco, P.D.; Corbett, S.A.; Lowry, S.F.; Graham, A.M.; et al. Rac1 is essential for basement membrane-dependent epiblast survival. Mol. Cell. Biol. 2010, 30, 3569–3581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.H.; Du, X.H.; Huang, K.; Zhang, T.; Teng, Z.; Niu, W.B.; Wang, C.; Xia, G.L. Rac1 modulates the formation of primordial follicles by facilitating STAT3-directed Jagged1, GDF9 and BMP15 transcription in mice. Sci. Rep. 2016, 6, 23972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, A.L.; Bridgham, J.T.; Woods, D.C. Cellular mechanisms and modulation of activin A- and transforming growth factor beta-mediated differentiation in cultured hen granulosa cells. Biol. Reprod. 2004, 71, 1844–1851. [Google Scholar] [CrossRef] [PubMed]
- Knapczyk-Stwora, K.; Grzesiak, M.; Witek, P.; Duda, M.; Koziorowski, M.; Slomczynska, M. Neonatal exposure to agonists and antagonists of sex steroid receptors induces changes in the expression of oocyte-derived growth factors and their receptors in ovarian follicles in gilts. Theriogenology 2019, 134, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dan, G.; Shangguan, T.; Hao, H.; Tang, R.; Peng, K.; Zhao, J.; Sun, H.; Zou, Z. miR-198 represses the proliferation of HaCaT cells by targeting Cyclin D2. Int. J. Mol. Sci. 2015, 16, 17018–17028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moons, D.S.; Jirawatnotai, S.; Tsutsui, T.; Franks, R.; Parlow, A.F.; Hales, D.B.; Gibori, G.; Fazleabas, A.T.; Kiyokawa, H. Intact follicular maturation and defective luteal function in mice deficient for cyclin-dependent kinase-4. Endocrinology 2002, 143, 647–654. [Google Scholar] [CrossRef]
- Xu, K.; Wang, J.; Liu, H.; Zhao, J.; Lu, W. Melatonin Promotes the Proliferation of Chicken Sertoli Cells by Activating the ERK/Inhibin Alpha Subunit Signaling Pathway. Molecules 2020, 25, 1230. [Google Scholar] [CrossRef] [Green Version]
- Lyu, Z.; Qin, N.; Tyasi, T.L.; Zhu, H.; Liu, D.; Yuan, S.; Xu, R. The Hippo/MST pathway member SAV1 plays a suppressive role in development of the prehierarchical follicles in hen ovary. PLoS ONE 2016, 11, e0160896. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.L.; Lee, J. Granulosa cell responsiveness to follicle stimulating hormone during early growth of hen ovarian follicles. Poult. Sci. 2015, 95, 108–114. [Google Scholar] [CrossRef]
- Xu, R.F.; Qin, N.; Xu, X.X.; Sun, X.; Chen, X.X.; Zhao, J.H. Implication of SLIT3-ROBO1/ROBO2 in granulosa cell proliferation, differentiation and follicle selection in the prehierarchical follicles of hen ovary. Cell Biol. Int. 2018, 42, 1643–1657. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Li, W.; Meng, X.; Zhang, L.; Shen, M.; Liu, H. Corticosterone Injection Impairs Follicular Development, Ovulation and Steroidogenesis Capacity in Mice Ovary. Animals 2019, 9, 1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Wang, Y.; Yuan, Z.; Chen, Y.; Guo, M.; Kang, L.; Sun, Y.; Jang, Y. The Role of PTHLH in Ovarian Follicle Selection, Its Transcriptional Regulation and Genetic Effects on Egg Laying Traits in Hens. Front. Genet. 2019, 10, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, N.; Fan, X.C.; Zhang, Y.Y.; Xu, X.X.; Tyasi, T.L.; Jing, Y.; Mu, F.; Wei, M.L.; Xu, R.F. New insights into implication of the SLIT/ROBO pathway in the prehierarchical follicle development of hen ovary. Poult. Sci. 2015, 94, 2235–2246. [Google Scholar] [CrossRef]
- Johnson, A.L. Ovarian follicle selection and granulosa cell differentiation. Poult. Sci. 2015, 94, 781–785. [Google Scholar] [CrossRef]
- Robinson, F.E.; Fasenko, G.M.; Renema, R.A. Female reproduction: Control of ovarian function. Optimizing chick production in broiler breeders. Optim. Chick Prod. Broiler Breed. 2003, 1, 3–10. [Google Scholar]
- Johnson, P.A.; Dickens, M.J.; Kent, T.R.; Giles, J.R. Expression and function of growth differentiation factor-9 in an oviparous species, Gallus domesticus. Biol. Reprod. 2005, 72, 1095–1100. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Von, O.S.; Robenek, H.; Diedrich, K.; Nofer, J.R. Follicular Fluid High-Density Lipoprotein-Associated Sphingosine 1-Phosphate (S1P) Promotes Human Granulosa Lutein Cell Migration via S1P Receptor Type 3 and Small G-Protein RAC1. Biol. Reprod. 2011, 84, 604–612. [Google Scholar] [CrossRef] [Green Version]
- Fortune, J.E.; Rivera, G.M.; Yang, M.Y. Follicular development: The role of the follicular microenvironment in selection of the dominant follicle. Anim. Reprod. Sci. 2004, 82, 109–126. [Google Scholar] [CrossRef]
- Zhu, G.; Fang, C.; Li, J.; Mo, C.; Wang, Y.; Li, J. Transcriptomic Diversification of Granulosa Cells during Follicular Development in Chicken. Sci. Rep. 2019, 9, 5462. [Google Scholar] [CrossRef]
- Pierce, J.G.; Parsons, T.F. Glycoprotein hormones: Structure and function. Annu. Rev. Biochem. 1981, 50, 465–495. [Google Scholar] [CrossRef]
- Cui, C.; Han, S.; Yin, H.; Luo, B.; Shen, X.; Yang, F.; Liu, Z.; Zhu, Q.; Li, D.; Wang, Y. FOXO3 Is Expressed in Ovarian Tissues and Acts as an Apoptosis Initiator in Granulosa Cells of Chickens. BioMed Res. Int. 2019, 9, 6902906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.H. Functions and Mechanisms of Rac1 in Mouse Follicular Assembly. Ph.D. Thesis, China Agricultural University, Beijing, China, 2016. [Google Scholar]
- Ubba, V.; Soni, U.K.; Chadchan, S.; Maurya, V.K.; Kumar, V.; Maurya, R.; Chaturvedi, H.; Singh, R.; Dwivedi, A.; Jha, R.K. RHOG-DOCK1-RAC1 Signaling Axis Is Perturbed in DHEA-Induced Polycystic Ovary in Rat Model. Reprod. Sci. 2017, 24, 738–752. [Google Scholar] [CrossRef] [PubMed]
- Pollack, S.E.; Furth, E.E.; Kallen, C.B.; Arakane, F.; Kiriakidou, M.; Kozarsky, K.F.; Strauss, J.F., III. Localization of the steroidogenic acute regulatory protein in human tissues. J. Clin. Endocrinol. Metab. 1997, 82, 4243–4251. [Google Scholar] [CrossRef] [PubMed]
- Bentsi-Barnes, I.K.; Kuo, F.T.; Barlow, G.M.; Pisarska, M.D. Human forkhead L2 represses key genes in granulosa cell differentiation including aromatase, P450scc, and cyclin D2. Fertil. Steril. 2010, 94, 353–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Xia, G.; Tsang, B.K. Regulation of cyclin D2 expression and degradation by follicle-stimulating hormone during rat granulosa cell proliferation in vitro. Biol. Reprod. 2013, 88, 57. [Google Scholar] [CrossRef] [PubMed]
- Park, T.J.; Chun, J.Y.; Bae, J.S.; Park, B.L.; Cheong, H.S.; Lee, H.S.; Kim, Y.J.; Shin, H.D. CCND2 polymorphisms associated with clearance of HBV infection. J. Hum. Genet. 2010, 55, 416–420. [Google Scholar] [CrossRef]
- Leung, A.Y.; Leung, J.C.; Chan, L.Y.; Ma, E.S.; Kwan, T.T.; Lai, K.N.; Meng, A.; Liang, R. Proliferating cell nuclear antigen (PCNA) as a proliferative marker during embryonic and adult zebrafish hematopoiesis. Histochem. Cell Biol. 2005, 124, 105–111. [Google Scholar] [CrossRef]
- Xu, H.; Khan, A.; Zhao, S.; Wang, H.; Zou, H.; Pang, Y.; Zhu, H. Effects of Inhibin A on Apoptosis and Proliferation of Bovine Granulosa Cells. Animals 2020, 10, 367. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) | Accession No. | Size | Efficiency | Annealing Temperatures |
|---|---|---|---|---|---|
| RAC1 | F: GACCCAAACTTGATTCCTAG | NM_205017.1 | 232 bp | 101.6% | 58 °C |
| R: GACGGTGCTGTAGGTAAA | |||||
| FSHR | F: TCCTGTGCTAACCCTTTCCTCTA | NM_205079.1 | 207 bp | 102.3% | 60 °C |
| R: AACCAGTGAATAAATAGTCCCATC | |||||
| CCND2 | F: AACTTGCTCTACGACGACC | NM_204213.1 | 150 bp | 99.8% | 59.5 °C |
| R: TTCACAGACCTCCAACATC | |||||
| PCNA | F: TGAATGAGCCAGTCCAG | NM_204170.2 | 144 bp | 100.5% | 59 °C |
| R: AGTGTCCCATATCAGCAA | |||||
| StAR | F: AGCAGATGGGCGACTGGAAC | AF220436.1 | 147 bp | 98.9% | 59.5 °C |
| R: GGGAGCACCGAACACTCACAA | |||||
| CYP11A1 | F: TCCGCTTTGCCTTGGAGTCTGTG | NM_001001756.1 | 112 bp | 103.1% | 59.5 °C |
| R: ATGAGGGTGACGGCGTCGATGAA | |||||
| 18SrRNA | F: TAGTTGGTGGAGCGATTTGTCT | AF173612.1 | 169 bp | 102.6% | 60 °C |
| R: CGGACATCTAAGGGCATCACA |
| Protein Target | Primary Antibody | Diluted | Secondary Antibody | Diluted |
|---|---|---|---|---|
| RAC1 | Rabbit anti-cRAC1 (Abcam, Cambridge, MA, USA) | 1:1000 | anti-rabbit IgG | 1:2000 |
| FSHR | Mouse anti-cFSHR (Boster, Biological Technology, Wuhan, China) | 1:1000 | anti-mouse IgG | 1:2000 |
| CCND2 | Mouse anti-cCCND2 (Invitrogen, Carlsbad, CA, USA) | 1:1000 | anti-mouse IgG | 1:2000 |
| PCNA | Rabbit anti-cPCNA (Sangon, Biotech Co., Ltd., Shanghai, China) | 1:1000 | anti-rabbit IgG | 1:2000 |
| StAR | Mouse anti-cStAR (Invitrogen, Carlsbad, CA, USA) | 1:1000 | anti-mouse IgG | 1:2000 |
| CYP11A1 | Mouse anti-cCYP11A1 (Invitrogen, Carlsbad, CA, USA) | 1:1000 | anti-mouse IgG | 1:2000 |
| β-actin | Mouse anti-cβ-actin (Boster, Biological Technology, Wuhan, China) | 1:1000 | anti-mouse IgG | 1:2000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyasi, T.L.; Sun, X.; Shan, X.; Liswaniso, S.; Chimbaka, I.M.; Qin, N.; Xu, R. Effects of RAC1 on Proliferation of Hen Ovarian Prehierarchical Follicle Granulosa Cells. Animals 2020, 10, 1589. https://doi.org/10.3390/ani10091589
Tyasi TL, Sun X, Shan X, Liswaniso S, Chimbaka IM, Qin N, Xu R. Effects of RAC1 on Proliferation of Hen Ovarian Prehierarchical Follicle Granulosa Cells. Animals. 2020; 10(9):1589. https://doi.org/10.3390/ani10091589
Chicago/Turabian StyleTyasi, Thobela Louis, Xue Sun, Xuesong Shan, Simushi Liswaniso, Ignatius Musenge Chimbaka, Ning Qin, and Rifu Xu. 2020. "Effects of RAC1 on Proliferation of Hen Ovarian Prehierarchical Follicle Granulosa Cells" Animals 10, no. 9: 1589. https://doi.org/10.3390/ani10091589
APA StyleTyasi, T. L., Sun, X., Shan, X., Liswaniso, S., Chimbaka, I. M., Qin, N., & Xu, R. (2020). Effects of RAC1 on Proliferation of Hen Ovarian Prehierarchical Follicle Granulosa Cells. Animals, 10(9), 1589. https://doi.org/10.3390/ani10091589