Simple Summary
Flystrike is a major ectoparasitic disease of sheep and it creates both an economic impact and welfare issue for the sheep industry. Several factors control the responses of sheep to flystrike, and among these, immune response is regarded as an important factor. Toll-like receptors (TLRs) plays a crucial role in the innate immune system by recognizing pathogen-associated molecular patterns derived from various microbes. Of these receptors, TLR9 recognises unmethylated cytosine-phosphate-guanine (CpG) motifs that are known to be prevalent in bacterial genomes and are also reported in Dipteran insects, including Lucilia cuprina, one of the main species associated with flystrike in sheep. In this study, we used a polymerase chain reaction-single strand conformation polymorphism (PCR-SSCP) technique to investigate TLR9 variation in sheep with and without flystrike, and found that variation in a gene region containing the coding sequence of the putative CpG-DNA binding domain was associated with the likelihood of flystrike occurrence. This suggests that variation in ovine TLR9 may affect a sheep’s response to flystrike.
Abstract
Toll-like receptors (TLRs) are a family of proteins that play a role in innate immune responses by recognising pathogen-associated molecular patterns derived from various microbes. Of these receptors, TLR9 recognises bacterial and viral DNA containing unmethylated cytosine-phosphate-guanine (CpG) motifs, and variation in TLR9 has been associated with resistance to various infectious diseases. Flystrike is a problem affecting the sheep industry globally and the immune response of the sheep has been suggested as one factor that influences the response to the disease. In this study, variation in ovine TLR9 from 178 sheep with flystrike and 134 sheep without flystrike was investigated using a polymerase chain reaction-single strand conformation polymorphism (PCR-SSCP) approach. These sheep were collected from both commercial and stud farms throughout New Zealand and they were of 13 different breeds, cross-breds and composites. Four alleles of TLR9 were detected, including three previously identified alleles (*01, *02 and *03) and a new allele (*04). In total six single nucleotide polymorphisms (SNPs) were found. Of the three common alleles in the sheep studied, the presence of *03 was found to be associated with a reduced likelihood of flystrike being present (OR = 0.499, p = 0.024). This suggests that variation in ovine TLR9 may affect a sheep’s response to flystrike, and thus the gene may have value as a genetic marker for improving resistance to the disease.
1. Introduction
Flystrike, or infestation of the fleece and skin with the larvae of various fly species, is a problem affecting the sheep industry globally, and the topic has been comprehensively reviewed recently [1]. It causes economic losses due to its negative impact on animal welfare and production performance, and there is increased cost associated with treating and preventing the disease. The occurrence of flystrike is environmentally dependent and its incidence and prevalence are variable, but there are several factors that make individual sheep more predisposed to flystrike, such as variation in the fleece, skin and immune response [2].
It has been determined that the immune system of a sheep can recognise the foreign components of a fly larva and produce an immune response to it [3], hence immune responses may play a role in defining flystrike resistance and susceptibility. However, using the Trangie research flocks in Australia, which were bred for resistance or susceptibility to fleece rot and flystrike, O’Meara et al. [4] found that antibody titres did not differ between the resistant and the susceptible animals. This led them to suggest that the resistance of sheep to fleece rot and flystrike was likely to be founded in the innate immune response, and not the adaptive response.
The toll-like receptors (TLRs) are a family of proteins that play a role in the innate immune response. Each TLR recognises particular patterns of pathogen-associated molecules, which can subsequently provoke innate immune responses [5]. The TLRs are widely expressed in immunologically active cells, including dendritic cells, macrophages, lymphocytes, and endothelial cells [6]. They are also classified into two subfamilies based on their localization on or in the cell: specifically the cell-surface TLRs and the intracellular TLRs [7].
The receptor TLR9 is an intracellular TLR and recognizes DNA containing unmethylated cytosine-phosphate-guanine (CpG) motifs [8]. Unmethylated CpG motifs are more prevalent in bacterial genomes than in vertebrate genomes, and their recognition by TLR9 triggers the activation of the mitogen-activated protein kinase pathway, nuclear factor kappa B, and multiple transcription and translation factors [9]. However, a recent study on 41 insect species from six orders revealed that DNA methylation was found in all orders investigated with the exception of the Diptera (flies), including Lucilia cuprina, one of the main species associated with flystrike in sheep [10].
Variation in human TLR9 gene (TRL9) has been reported to be associated with variation in susceptibility to a diverse range of diseases, such as Crohn’s disease [11], lupus erythematosus [12], malaria [13] and talaromycosis [14]. In cattle, variation in TLR9 has been associated with susceptibility to bovine tuberculosis [15], and in sheep it has been reported to be associated with seropositivity for small ruminant lentivirus in the Boutsko, Friesarta and Comisana breeds in Greece [16], and in 91 flocks of sheep reared in Italy [17]. Collectively these studies suggest a role for TLR9 in the immune response to infectious and other diseases.
In this study, we collected blood samples from sheep with flystrike, and phenotypically normal sheep from the same flocks for a variety of sheep breeds farmed in New Zealand (NZ). We used polymerase chain reaction-single strand conformation polymorphism (PCR-SSCP; see [18] for a review of its applicability) to analyse variation in TLR9 and looked for associations between variation in TLR9 and the occurrence of flystrike.
2. Materials and Methods
2.1. Animals
One hundred and seventy-eight sheep that had flystrike or were recovering from flystrike were sampled from three strike seasons (2015–2017), and 134 sheep that had no symptoms of flystrike were also sampled from the same farms at the same time. These sheep were from both commercial and stud breeding operations throughout NZ and they were of different breeds, including: Merino, Corriedale, Perendale, Romney, Lincoln, Coopworth, Poll Dorset, Texel, Finn, Dorset Down, Suffolk, South Suffolk, Shropshire, and various crossbred sheep and composites. The sheep were from farms where the farmers had used a variety of management practices to control flystrike.
2.2. Identification of Flystrike
Sheep with flystrike, and those that had had flystrike, were identified either during shearing, or in the yard. These sheep were diagnosed as having active flystrike when larvae could be seen on the skin, and their wool was discoloured and bad smelling. Sheep that had had flystrike were identified through having areas of pink skin with little or no wool, obvious scar tissue from larval damage and/or flaky, dry skin.
2.3. Blood Collection
Blood samples from individual sheep were taken via ear nicking and collection onto FTATM cards (Whatman, Middlesex, UK). This approach to blood collection complies with the Animal Identification Section 7.5 of the Animal Welfare (Sheep and Beef Cattle) Code of Welfare 2010, which is a welfare issued code under the New Zealand Animal Welfare Act 1999 (New Zealand Government). In contemporary NZ farming, both blood and ear tissue samples are regularly collected for the purpose of animal breeding, and this can include the use of commercially available single-gene diagnostic tests for both disease and performance traits, and the use of genome wide screening approaches including SNP chip typing and whole genome sequencing. Accordingly, ethics approval was not directly required for this research.
Genomic DNA for the PCR amplifications was purified from the dried blood spots using a two-step procedure described by Zhou et al. [19].
2.4. PCR-SSCP Analysis and Genotyping of Ovine TLR9
A 414-bp fragment of ovine TLR9 exon 2 that encodes for the Z-loop [20] and a putative CpG-DNA binding domain [21], was amplified using the PCR primers described in Zhou et al. [22]. These primers were 5′-ttcgtggacctgtcggac-3′ and 5′-ctggctgttgtagctgag-3′, and they were synthesised by Integrated DNA Technologies (Coralville, IA, USA).
PCR amplification was performed in a 15-μL reaction. This contained the purified genomic DNA on a 1.2 mm punch of the FTATM card, 0.25 μM of each primer, 150 μM dNTP’s (Bioline, London, UK), 2.5 mM of Mg2+, 0.5 U of Taq DNA polymerase (Qiagen, Hilden, Germany) and 1× the reaction buffer supplied with the enzyme. The thermal profile for amplification consisted of 2 min denaturation at 94 °C, followed by 34 cycles of 30 s at 94 °C, 30 s at 61 °C and 30 s at 72 °C, with a final extension of 5 min at 72 °C. Amplification was carried out in S1000 thermal cyclers (Bio-Rad, Hercules, CA, USA).
A 0.7-μL aliquot of each amplicon was mixed with 7 μL of loading dye (98% formamide, 10 mM EDTA, 0.025% bromophenol blue, 0.025% xylene-cyanol). After denaturation of the amplicon at 95 °C for 5 min, the samples were rapidly cooled on wet ice and then loaded on 16 cm × 18 cm, 14% acrylamide: bisacrylamide (37.5:1) (Bio-Rad) gels. Electrophoresis was performed using Protean II xi cells (Bio-Rad), at 300 V for 19 h at 23 °C in 0.5×TBE buffer. Polymerase chain reaction amplicons of three previously known alleles (*01, *02 and *03) of ovine TLR9 [22] were run as standards to identify the TLR9 alleles present in the individual samples. The gels were silver-stained according to the method of Byun et al. [23].
2.5. Sequencing of New Alleles and Sequence Analysis
Alleles that exhibited PCR-SSCP patterns different to those of the previous identified alleles *01, *02 and *03 were regarded as new alleles and were sequenced at the Lincoln University DNA Sequencing Facility. For those alleles that were only found in heterozygous samples, they were sequenced using an approach described previously [24]. Briefly, single bands of interest from the heterozygous were recovered directly from the SSCP gels as a gel slice. These were macerated and the DNA was eluted into 50 µL TE buffer by incubating at 70 °C for 20 min. One µL of the eluted solution was used as a template for PCR amplification to produce PCR amplicons with SSCP patterns that represented a hemizygous portion of the patterns derived from the original heterogeneous amplicons. These ‘homozygous’ PCR amplicons were then directly sequenced. Sequence alignment and comparisons were carried out using DNAMAN (version 5.2.10, Lynnon BioSoft, Vaudreuil, QC, Canada).
2.6. Statistical Analyses
All statistical analyses were performed using R Statistics version 3.6.0. First, univariate Pearson Chi-square tests were undertaken to explore the relationships between the known variables (age, gender, year, wool type, breed, farm, and region) and the occurrence of flystrike to ascertain whether any confounding factors needed to be adjusted for. Next, for each of the four TRL9 alleles, a Pearson Chi-square test along with a binary logistic regression was performed to explore the relationship between the presence of each allele and the occurrence of flystrike, with a correction for the variables applied, if they were associated with the occurrence of the disease.
3. Results
3.1. Alleles of TLR9 and Their Frequencies
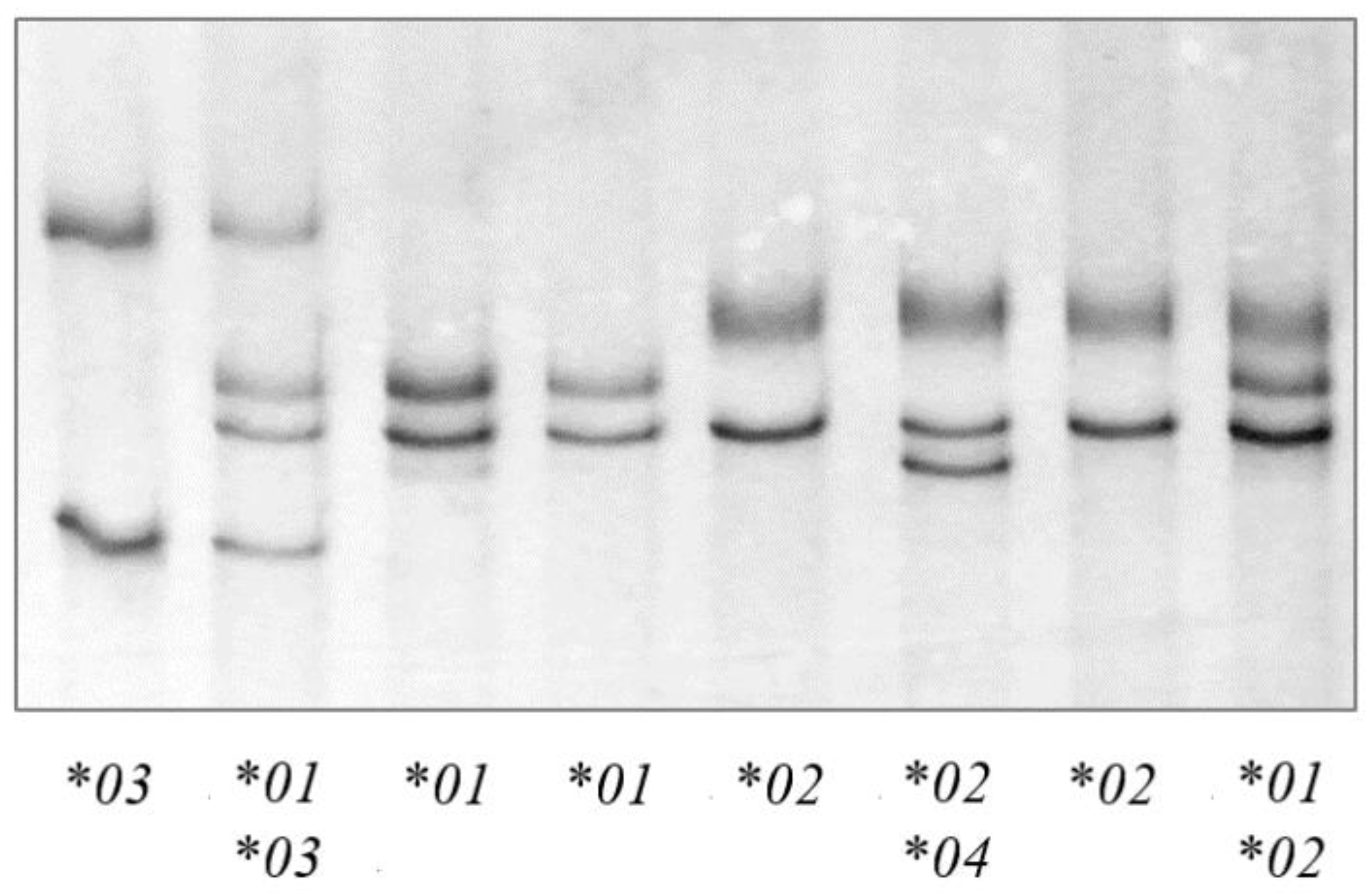
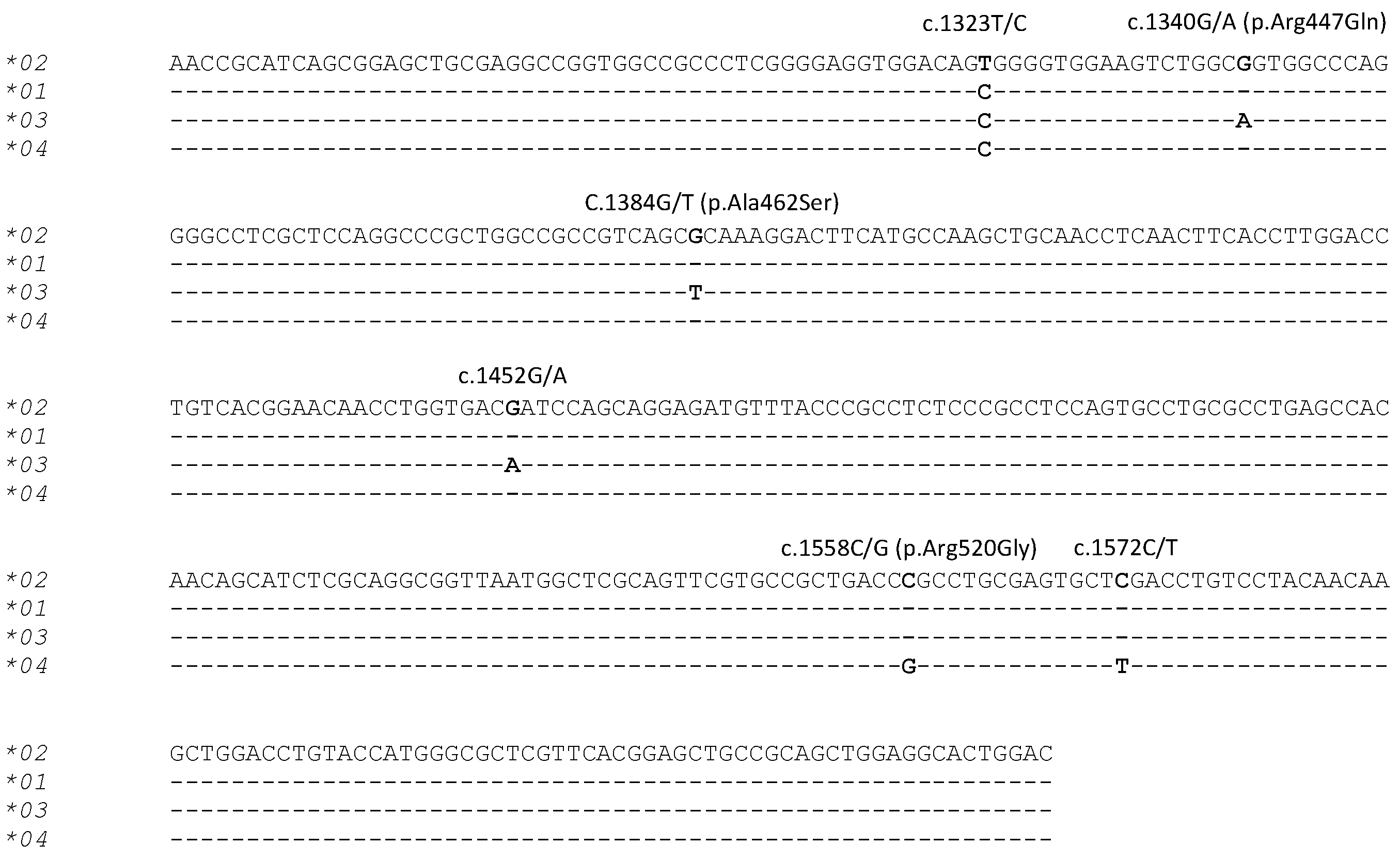
PCR-SSCP analysis of the 312 sheep investigated revealed four unique banding patterns (*01 to *04) (Figure 1). The first three banding patterns matched those of the three previously identified alleles *01 to *03 [22], whereas the fourth patterns represented a new allele *04 (Figure 2). The identification of this new allele led to the addition of two further single nucleotide polymorphisms (SNPs) to the gene region investigated, including one synonymous SNP (c.1572C/T) and one non-synonymous SNP (c.1558C/G), which would led to the amino acid change p.Arg520Gly.
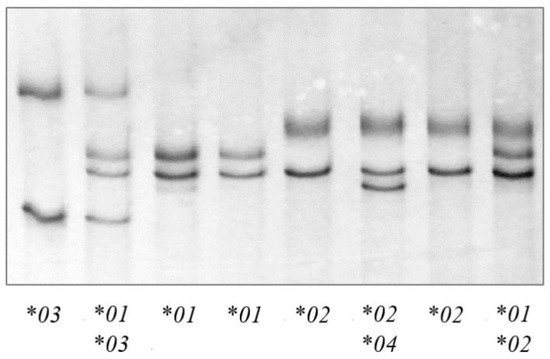
Figure 1.
Polymerase chain reaction—single strand conformation polymorphism (PCR-SSCP) analysis of the ovine TLR9 exon 2 region. Four unique banding patterns representing four alleles (*01, *02, *03 and *04) were observed in either homozygous or heterozygous forms.
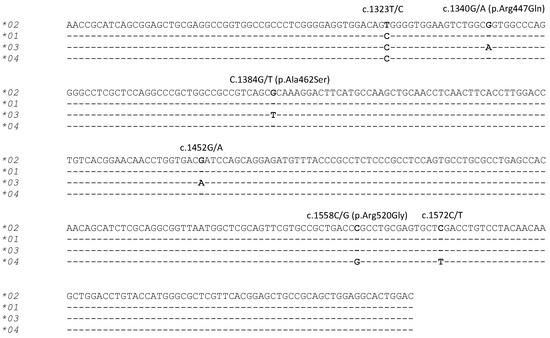
Figure 2.
Alignment of the four ovine TLR9 allele nucleotide sequences. Six SNPs identified in the three previously identified alleles (*01, *02 and *03) [22] and the newly identified allele (*04) are marked above the sequences with the SNPs marked in bold. The annotation of the SNPs and amino acid substitutions follows the HGVS nomenclature (http://www.hgvs.org/mutnomen/). The sequences exclude the PCR primer binding regions and dashes represent nucleotide sequences identical to the reference sequence (Allele *02).
The number and percentage of sheep carrying the TLR9 alleles in the flystrike and non-flystrike groups are shown in Table 1. Among the three common alleles, the largest difference between these two groups was observed for the percentage of sheep carrying allele *03. In the flystrike group, 17.4% of sheep carried *03, compared to 26.1% of sheep who carried *03 in the non-flystrike group.

Table 1.
Number and percentage of animals carrying individual TLR9 alleles, for sheep with and without.
3.2. Associations between the Variables and Flystrike Occurrence
The categories of the variables (age, gender, year, wool type, breed, farm, and region) and their frequencies are summarized in Supplementary Table S1. The univariate Pearson Chi-square tests between the variables (age, gender, year, wool type, breed, farm, and region) and the occurrence of flystrike are shown in Supplementary Table S2. No significant association was detected between these variables and flystrike occurrence, suggesting there was no need to correct for these variables.
Pearson Chi-square tests along with binary logistic regression using the occurrence of flystrike as dependent variable and the allele as independent variable, revealed a relationship between the presence of TLR9 allele *03 and flystrike (Table 2), with the presence of allele *03 being associated with a lower likelihood of flystrike occurrence (OR = 0.499, p = 0.024).
Table 2.
The association of the presence of each TRL9 allele with the occurrence of flystrike.
4. Discussion
This study was undertaken to investigate whether variation in ovine TLR9 was related to variation in flystrike susceptibility. A new allele sequence of TLR9 was identified, which may be a consequence of more sheep from different sheep breeds being investigated in this study, when compared to the work of Zhou et al. [22].
The detection of this new allele brings the number of SNPs identified in this 378-bp fragment of exon 2 of TLR9 (excluding the primer binding regions) from four to six, giving an average SNP density of 15.9 SNPs per kb of genome. This SNP density is comparable to that reported for other highly polymorphic genes, such as the keratin-associated protein genes KRTAP28-1 [25] and KRTAP36-1 [26]. It is notably higher than the average density of 4.9 SNPs per kb across the sheep genome suggested by Kijas et al. [27], such that delineating this variation using tools like the Illumina OvineSNP50 DNA Analysis Kits (Illumina Inc., San Diego CA, USA) may be difficult. Even with the claimed 54,241 evenly spaced probes in this kit, and the assertion that this is sufficient SNP density for genome-wide association studies, it is still only detecting an average one SNP per 55 kb, compared to the one SNP per 63 bp in the four different coding region sequences described here. In this respect it is notable that half of the SNPs identified in TLR9 are non-synonymous, and that this may be important for the function of the receptor.
Of the three common alleles in the sheep studied, association with occurrence of flystrike was only detected for allele *03. This allele contained three unique nucleotide sequences at positions c.1340, c.1384 and c.1452, when compared to alleles *01 and *02. This suggests that the SNPs at these positions may have effect on the function of the receptor. Unlike many other TLRs, TLR9 possesses a loop region (known as the Z-loop) composed of approximately 38 amino acid residues [20]. Proteolytic processing at the Z-loop has been revealed to be necessary for creation of a functional TLR9 protein [28]. In this respect, the SNPs c.1340 and c.1384 are located within the Z-loop encoding region of the gene, and if translated, would lead to the amino acid changes p.Arg446Gln and p.Ala462Ser. These amino acid changes may affect the structure or proteolytic cleavage of TLR9, and consequently have functional effect on its binding activity and function in immune responses.
Additionally, the potential effect of the synonymous SNP at c.1452 cannot be ignored, as synonymous SNPs have been reported to influence mRNA stability and translation [29], and/or protein folding [30,31], or affect gene expression via codon preference differences, thereby potentially resulting in altered expression levels [17,30].
Alternatively, the SNPs detected here may not have any functional effect by themselves, but may be in linkage with variation in other regions of the gene, or nearby genes with functional or structural significance. In this respect, in a study of the Trangie sheep, Smith et al. [2] suggested that the genes for fibulin (FBLN1) and fatty-acid binding protein 4 (FABP4) are associated with to resistance to fleece rot in Australian Merino sheep. Subsequently, Burrows et al. [32] typed ovine FABP4 and confirmed an association between the gene and flystrike susceptibility.
In the context of the association revealed here, TLR9 (ENSOARG00000006162) is located on ovine chromosome 19, while FABP4 (ENSOARG00000009344) is on chromosome 9 and FBLN1 (ENSOARG00020001970) is on chromosome 3, suggesting that the association revealed with TLR9 is unlikely to be because of the latter two genes. The TLR9 gene is in proximity to the twinfilin actin binding protein 2 gene (TWF2: ENSOARG00000005975) on ovine chromosome 19, but this actin-binding protein is thought to be involved in motile and morphological processes in mammals, thus it having a role in susceptibility to flystrike seems unlikely.
5. Conclusions
Four alleles of TLR9 were detected in 312 sheep genomes, of which *04 was found for the first time, and *02 was the dominant allele in the sheep studied, followed by *03. The presence of TRL9 *03 was found to be associated with a lower likelihood of sheep having flystrike. If allele *03 is an effective molecular marker for selection of flystrike resistance in sheep, then it could potentially be used as a genetic marker for breeding, but to be confident of this approach the relationship between TLR9 variation and susceptibility needs to be tested more widely.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/ani11123549/s1, Table S1: Categorisation of the variables; Table S2: Pearson Chi-square analyses exploring associations between the variables and flystrike occurrence.
Author Contributions
Conceptualization, H.Z., Y.L. and J.G.H.H.; methodology, X.L., H.G., Q.F.; formal analysis, H.Z., W.L., H.G., J.W., S.L., investigation, X.L., Q.F., resources, J.G.H.H.; data curation, H.Z. and J.G.H.H.; writing—original draft preparation, X.L., H.Z., W.L. and J.G.H.H.; writing—review and editing, H.Z., J.H. and J.G.H.H.; project administration, S.L. and J.H.; funding acquisition, Y.L. and J.G.H.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Open Project of Gansu Key Laboratory of Herbivorous Animal Biotechnology (grant number GKLAB-201601), the International Science and Technology Cooperation Project of Gansu Province (grant number 18YF1WA082), the Gansu Province Higher Education Innovation Fund Project (grand number 2020A-52) and the Youth Tutor Fund Project of Gansu Agricultural University (grant number GAU-QDFC-2018-11).
Institutional Review Board Statement
Ethical review and approval were waived for this study research, due to that the collection of sheep blood drops by the nicking of their ears was covered by Section 7.5 Animal Identification, in: Code of Welfare: Sheep and Beef Cattle (2016); a code of welfare issued under the Animal Welfare Act 1999 (New Zealand Government).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Acknowledgments
The authors would like to thank Lucy Burrows for the collection of blood samples and phenotype data.
Conflicts of Interest
All authors declare no conflict of interest.
References
- Anstead, C.A.; Perry, T.; Richards, S.; Korhonen, P.K.; Young, N.D.; Bowles, V.M.; Batterham, P.; Robin, B.; Gasser, R.B. The Battle against Flystrike–Past Research and New Prospects through Genomics. Adv. Parasitol. 2017, 98, 227–281. [Google Scholar] [PubMed]
- Smith, W.J.; Li, Y.; Ingham, A.; Collis, E.; McWilliam, S.M.; Dixon, T.J.; Norris, B.J.; Mortimer, S.I.; Moore, R.J.; Reverter, A. A genomics-informed, SNP associated study reveals FBLN1 and FABP4 as contributing to resistance to fleecerot in Australian Merino sheep. BMC Vet. Res. 2010, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDiarmid, J.; Clarke, R.; McClure, S.; Bowen, F.; Burrell, D. Use of a monoclonal antibody to ovine IgE for fly strike studies in sheep. Int. J. Parasitol. 1995, 25, 1505–1507. [Google Scholar] [CrossRef]
- O’Meara, T.J.; Nesa, M.; Sandeman, R.M. Antibody responses to Lucilia cuprina in sheep selected for resistance or susceptibility to L. cuprina. Parasite Immunol. 1997, 19, 535–543. [Google Scholar] [CrossRef]
- Kumagai, Y.; Akira, S. Identification and functions of pattern-recognition receptors. J. Allergy Clin. Immunol. 2010, 125, 985–992. [Google Scholar] [CrossRef]
- Kang, J.Y.; Lee, J.O. Structural biology of the Toll-like receptor family. Annu. Rev. Biochem. 2011, 80, 917–941. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Takeda, K.; et al. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 408, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Krieg, A.M. CpG motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 2002, 20, 709–760. [Google Scholar] [CrossRef]
- Bewick, A.J.; Vogel, K.J.; Moore, A.J.; Schmitz, R.J. Evolution of DNA Methylation across Insects. Mol. Biol. Evol. 2017, 34, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Török, H.P.; Glas, J.; Tonenchi, L.; Bruennler, G.; Folwaczny, M.; Folwaczny, C. Crohn’s disease is associated with a toll-likereceptor-9 polymorphism. Gastroenterology 2004, 127, 365–366. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, C.; Zhou, Z.; Pei, F. TLR9 polymorphisms and systemic lupus erythematosus risk: An update meta-analysis study. Rheumatol. Int. 2016, 36, 585–595. [Google Scholar] [CrossRef]
- Omar, A.H.; Yasunami, M.; Yamazaki, A.; Shibata, H.; Ofori, M.F.; Akanmori, B.D.; Shuaibu, M.N.; Kikuchi, M.; Hirayama, K. Toll-like receptor 9 (TLR9) polymorphism associated with symptomatic malaria: A cohort study. Malar. J. 2012, 11, 168. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Li, L.; Xiao, S.; Chen, W.; Hu, F.; Li, F.; Guo, P.; Chen, X.; Cai, W.; Tang, X. The association of TLR2, TLR3, and TLR9 gene polymorphisms with susceptibility to Talaromycosis among Han Chinese AIDS patients in Guangdong. Front. Cell Infect. Microbiol. 2021, 11, 625461. [Google Scholar] [CrossRef] [PubMed]
- Alfano, F.; Peletto, S.; Lucibelli, M.G.; Borriello, G.; Urciuolo, G.; Maniaci, M.G.; Desiato, R.; Tarantino, M.; Barone, A.; Pasquali, P.; et al. Identification of single nucleotide polymorphisms in Toll-like receptor candidate genes associated with tuberculosis infection in water buffalo (Bubalus bubalis). BMC Genet. 2014, 15, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarafidou, T.; Stamatis, C.; Kalozoumi, G.; Spyrou, V.; Fthenakis, G.C.; Billinis, C.; Mamuris, Z. Toll like receptor 9 (TLR9) polymorphism G520R in sheep is associated with seropositivity for small ruminant lentivirus. PLoS ONE 2013, 8, e63901. [Google Scholar]
- Arcangeli, C.; Lucarelli, D.; Torricelli, M.; Sebastiani, C.; Ciullo, M.; Pellegrini, C.; Felici, A.; Costarelli, S.; Giammarioli, M.; Feliziani, F.; et al. First survey of SNPs in TMEM154, TLR9, MYD88 and CCR5 genes in sheep reared in Italy and their association with resistance to SRLVs infection. Viruses 2021, 13, 1290. [Google Scholar] [CrossRef]
- Kakavas, K.V. Sensitivity and applications of the PCR Single-Strand Conformation Polymorphism method. Mol. Biol. Rep. 2021, 48, 3629–3635. [Google Scholar] [CrossRef]
- Zhou, H.; Hickford, J.G.H.; Fang, Q. A two-step procedure for extracting genomic DNA from dried blood spots on filter paper for polymerase chain reaction amplification. Anal. Biochem. 2006, 354, 159–161. [Google Scholar] [CrossRef]
- Ohto, U.; Shibata, T.; Tanji, H.; Ishida, H.; Krayukhina, E.; Uchiyama, S.; Miyake, K.; Shimizu, T. Structural basis of CpG and inhibitory DNA recognition by Toll-like receptor 9. Nature 2015, 520, 702–705. [Google Scholar] [CrossRef] [PubMed]
- Rutz, M.; Metzger, J.; Gellert, T.; Luppa, P.; Lipford, G.B.; Wagner, H.; Bauer, S. Toll-like receptor 9 binds single-stranded CpG-DNA in a sequence- and pH-dependent manner. Eur. J. Immunol. 2004, 34, 2541–2550. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Hickford, J.G.H. Polymorphism of Toll-like receptor 9 (TLR9) gene in sheep. Vet. Immunol. Immunopath. 2008, 121, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.O.; Fang, Q.; Zhou, H.; Hickford, J.G.H. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal. Biochem. 2009, 385, 174–175. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Yu, Z.; Dyer, J.; Plowman, J.E.; Hickford, J. Identification of the ovine keratin-associated protein KAP1-2 gene (KRTAP1-2). Exp. Dermatol. 2011, 20, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Wang, J.; Zhou, H.; Gong, H.; Tao, J.; Hickford, J.G.H. Identification of ovine KRTAP28-1 and its association with wool fibre diameter. Animals 2019, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Zhou, H.; Wang, J.; Li, S.; Luo, Y.; Hickford, J.G.H. Characterisation of an ovine keratin associated protein (KAP) gene, which would produce a protein rich in glycine and tyrosine, but lacking in cysteine. Genes 2019, 10, 848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kijas, J.W.; Townley, D.; Dalrymple, B.P.; Heaton, M.P.; Maddox, J.F.; McGrath, A.; Wilson, P.; Ingersoll, R.G.; McCulloch, R.; McWilliam, S. A genome wide survey of SNP variation reveals the genetic structure of sheep breeds. PLoS ONE 2009, 4, e4668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, B.; Brinkmann, M.M.; Spooner, E.; Lee, C.C.; Kim, Y.M.; Ploegh, H.L. Proteolytic cleavage in an endolysosomal compartment is required for activation of Toll-like receptor 9. Nat. Immunol. 2008, 9, 1407–1414. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Wainwright, M.S.; Comeron, J.M.; Saitou, N.; Sanders, A.R.; Gelernter, J.; Gejman, P.V. Synonymous mutations in the human dopamine receptor D2 (DRD2) affect mRNA stability and synthesis of the receptor. Hum. Mol. Genet. 2003, 12, 205–216. [Google Scholar] [CrossRef]
- Kimchi-Sarfaty, C.; Oh, J.M.; Kim, I.W.; Sauna, Z.E.; Calcagno, A.M.; Ambudkar, S.V.; Gottesman, M.M. A “silent” polymorphism in the MDR1 gene changes substrate specificity. Science 2007, 315, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Komar, A.A.; Lesnik, T.; Reiss, C. Synonymous codon substitutions affect ribosome traffic and protein folding during in vitro translation. FEBS Lett. 1999, 462, 387–391. [Google Scholar] [CrossRef] [Green Version]
- Burrows, L.E.R.; Zhou, H.; Frampton, C.M.A.; Forrest, R.H.J.; Hickford, J.G.H. Ovine FABP4 variation and its association with flystrike susceptibility. Front. Genet. 2021, 12, 675305. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).