
Effect of a Synbiotic Supplement on Fear Response and Memory Assessment of Broiler Chickens Subjected to Heat Stress


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Synbiotic
2.3. Animals and Housing
2.4. Dietary Treatments
2.5. Behavioral Tests
2.5.1. Object Memory Test
2.5.2. Novel Object Test
2.5.3. Touch Test
2.5.4. Isolation Test
2.5.5. Tonic Immobility (TI)
2.6. Blood sample Collection
2.6.1. Heterophil/Lymphocyte (H/L) Ratio
2.6.2. Plasma Tryptophan and Corticosterone Analyses
2.7. Statistical Analysis
3. Results
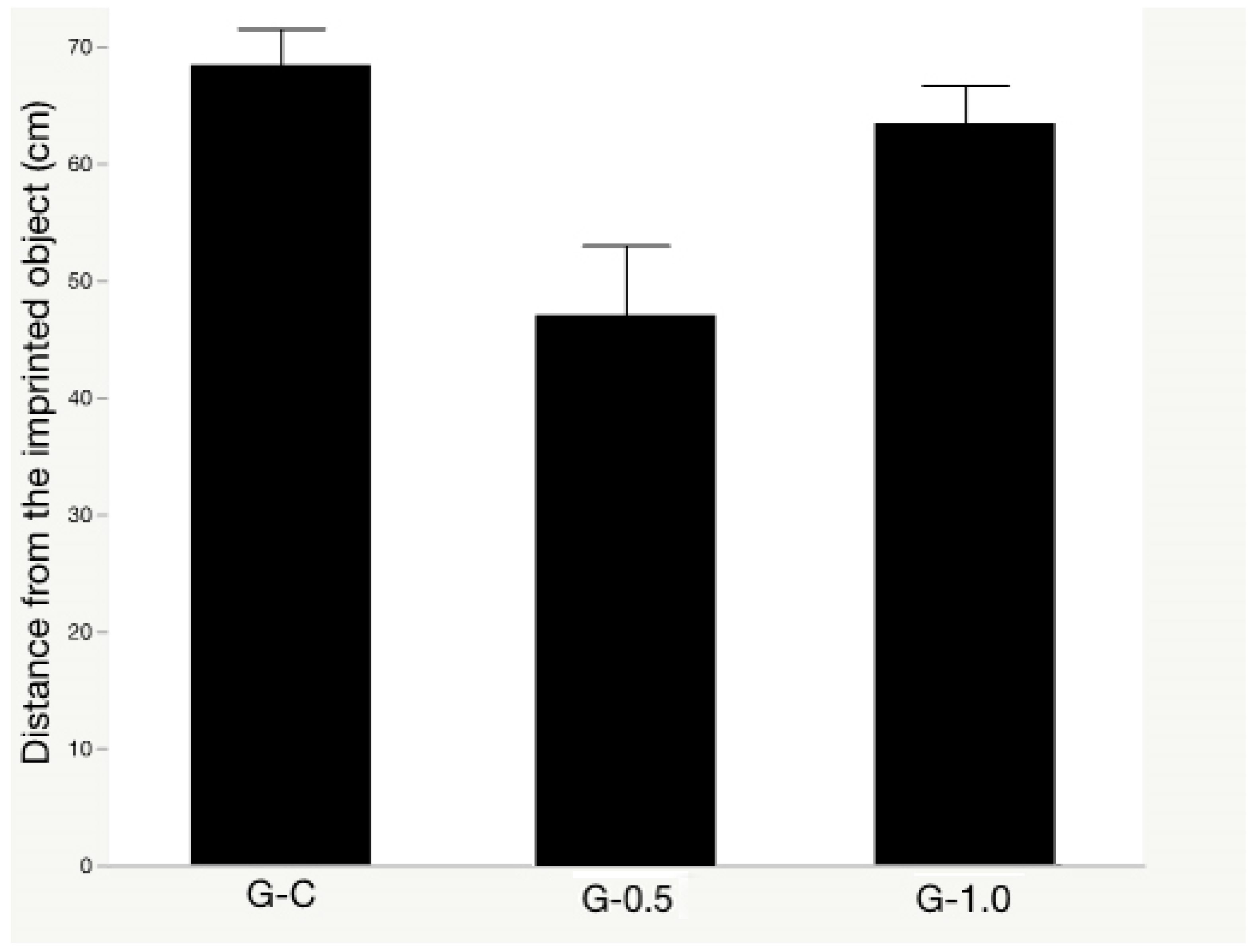
3.1. Object Memory Test
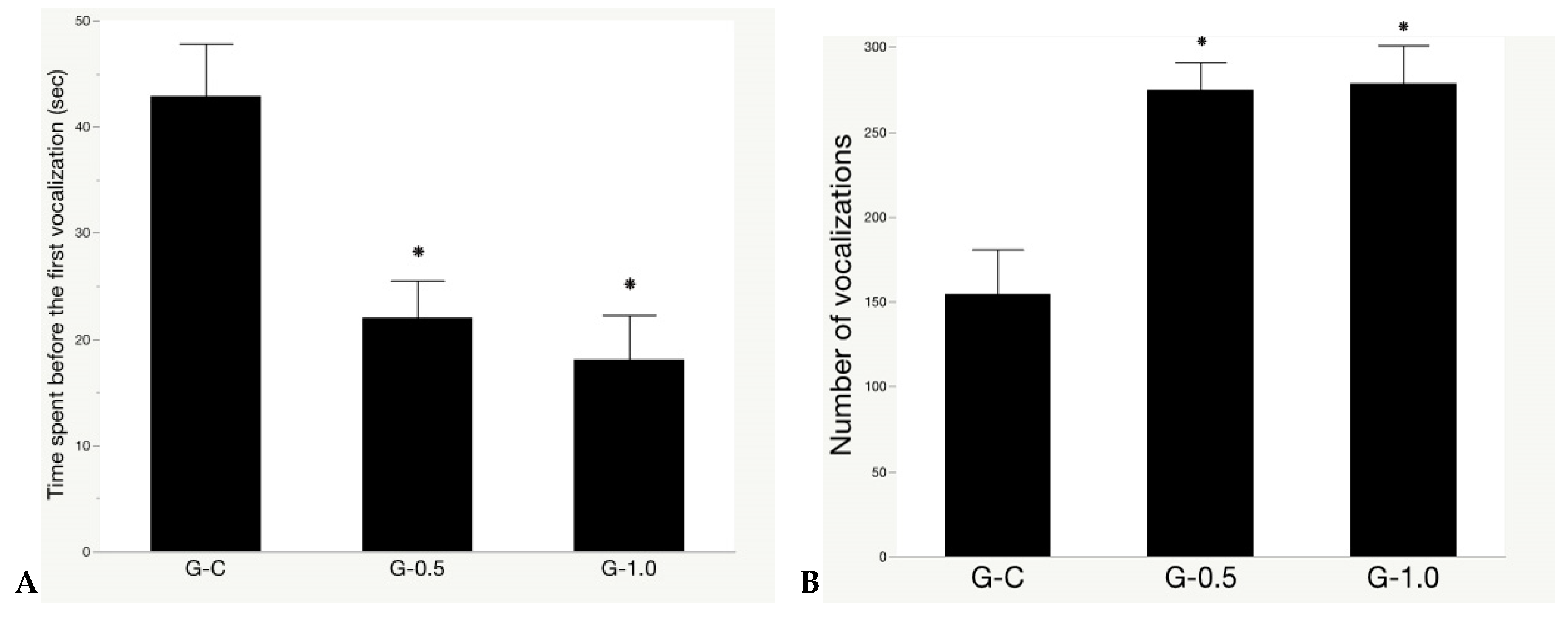
3.2. Isolation Test
3.3. Novel Object Test, Touch Test, and Tonic Immobility (TI)Test
3.4. Plasma Tryptophan and Corticosterone Analyses
3.5. Heterophil/Lymphocyte (H/L) Ratio
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Russo, S.; Sillmann, J.; Sterl, A. Humid heat waves at different warming levels. Sci. Rep. 2017, 7, 7477. [Google Scholar] [CrossRef]
- Hartcher, K.M.; Lum, H.K. Genetic selection of broilers and welfare consequences: A review. World Poult. Sci. J. 2020, 76, 154–167. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, V.S. Heat stress biomarker amino acids and neuropeptide afford thermotolerance in chicks. J. Poult. Sci. 2019, 56, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Chronic heat stress impairs the quality of breast-muscle meat in broilers by affecting redox status and energy-substance metabolism. J. Agric. Food Chem. 2017, 65, 11251–11258. [Google Scholar] [CrossRef]
- Shin, J.E.; Kim, J.H.; Goo, D.; Han, G.P.; Pitargue, F.M.; Kang, H.K.; Kil, D.Y. Effect of dietary supplementation of betaine on productive performance, egg quality and jejunal tight junction-related gene expression in laying hens raised under hot environmental conditions. Livest. Sci. 2018, 214, 79–82. [Google Scholar] [CrossRef]
- Goo, D.; Kim, H.J.H.; Park, G.H.; Reyes, J.B.D.; Kil, D.Y. Effect of heat stress and stocking density on growth performance, breast meat quality, and intestinal barrier function in broiler chickens. Animals 2019, 9, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic losses from heat stress by US livestock industries. J. Dairy Sci. 2003, 86, 52–77. [Google Scholar] [CrossRef] [Green Version]
- Somero, G.N. The cellular stress response and temperature: Function, regulation, and evolution. J. Exp. Zool. A Ecol. Integr. Physiol. 2020, 333, 1–73. [Google Scholar] [CrossRef] [PubMed]
- Cianconi, P.; Betrò, S.; Janiri, L. The impact of climate change on mental health: A systematic descriptive review. Front. Psychiatry 2020, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Palinkas, L.A.; Wong, M. Global climate change and mental health. Curr. Opin. Psychol. 2020, 32, 12–16. [Google Scholar] [CrossRef]
- Xu, Y.; Lai, X.; Li, Z.; Zhang, X.; Luo, Q. Effect of chronic heat stress on some physiological and immunological parameters in different breed of broilers. Poult. Sci. 2018, 97, 4073–4082. [Google Scholar] [CrossRef] [PubMed]
- He, X.F.; Lu, Z.; Ma, B.B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, J.; Gao, F. Chronic heat stress alters hypothalamus integrity, the serum indexes and attenuates expressions of hypothalamic appetite genes in broilers. J. Therm. Biol. 2019, 81, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Iñiguez, S.D.; Flores-Ramirez, F.J.; Riggs, L.M.; Alipio, J.B.; Garcia-Carachure, I.; Hernandez, M.A.; Sanchez, D.O.; Lobo, M.K.; Serrano, P.A.; Braren, S.H.; et al. Vicarious social defeat stress induces depression-related outcomes in female mice. Biol. Psychiatry 2018, 83, 9–17. [Google Scholar] [CrossRef]
- Weina, H.; Yuhu, N.; Christian, H.; Birong, L.; Feiyu, S.; Le, W. Liraglutide attenuates the depressive- and anxiety-like behaviour in the corticosterone induced depression model via improving hippocampal neural plasticity. Brain Res. 2018, 1694, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Campbell, D.L.M.; Dickson, E.J.; Lee, C. Application of open field, tonic immobility, and attention bias tests to hens with different ranging patterns. Peer J. 2019, 7, e8122. [Google Scholar] [CrossRef] [PubMed]
- Bock, B.B.; Jong, I.D. Assessment of Animal Welfare in Broiler Farm; Welfare Quality® Reports no. 18 Cardiff: Cardiff Univesity Press, May 2010. Available online: http://www.welfarequality.net/media/1126/wqr18.pdf (accessed on 4 October 2020).
- Dennis, R.L.; Lay, D.L.; Cheng, H.W. Effects of early serotonin programming on behavior and central monoamine concentrations in an avian model. Behav. Brain Res. 2013, 253, 290–296. [Google Scholar] [CrossRef]
- Zulkifli, I.; Al-Aqil, A.; Omar, A.R.; Sazili, A.Q.; Rajion, M.A. Crating and heat stress influence blood parameters and heat shock protein 70 expression in broiler chickens showing short or long tonic immobility reactions. Poult. Sci. 2009, 88, 471–476. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar] [PubMed]
- Le Sciellour, M.; Zemb, O.; Hochu, I.; Riquet, J.; Gilbert, H.; Giorgi, M.; Billon, Y.; Gourdine, J.L.; Renaudeau, D. Effect of chronic and acute heat challenges on fecal microbiota composition, production, and thermoregulation traits in growing pigs. J. Anim. Sci. 2019, 97, 3845–3858. [Google Scholar] [CrossRef]
- Xia, Z.; Huang, L.; Yin, P.; Liu, F.; Liu, Y.; Zhang, Z.; Lin, J.; Zou, W.; Li, C.L. Arginine alleviates heat stress-induced intestinal epithelial barrier damage by promoting expression of tight junction proteins via the AMPK pathway. Mol. Biol. Rep. 2019, 46, 6435–6451. [Google Scholar] [CrossRef] [Green Version]
- Kpodo, K.R.; Duttlinger, A.W.; Radcliffe, J.S.; Johnson, J.S. Time course determination of the effects of rapid and gradual cooling after acute hyperthermia on body temperature and intestinal integrity in pigs. J. Therm. Biol. 2020, 87, 102481. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Yan, J.; Li, M.; Ying, S.; Shi, Z. Gut microbiota dysbiosis increases the risk of visceral gout in goslings through translocation of gut-derived lipopolysaccharide. Poult. Sci. 2019, 98, 5361–5373. [Google Scholar] [CrossRef]
- Bahrami, A.; Barreto, G.E.; Lombardi, G.; Pirro, M.; Sahebkar, A. Emerging roles for high-density lipoproteins in neurodegenerative disorders. BioFactors 2019, 45, 725–739. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C. The endotoxin hypothesis of neurodegeneration. J. Neuroinflammation 2019, 16, 180. [Google Scholar] [CrossRef] [Green Version]
- Chunchai, T.; Thunapong, W.; Yasom, S.; Wanchai, K.; Eaimworawuthikul, S.; Metzler, G.; Lungkaphin, A.; Pongchaidecha, A.; Sirilun, S.; Chaiyasut, C.; et al. Decreased microglial activation through gut-brain axis by prebiotics, probiotics, or synbiotics effectively restored cognitive function in obese-insulin resistant rats. J. Neuroinflammion 2018, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Jiang, Y.; Xu, K.; Cui, M.; Ye, W.; Zhao, G.; Jin, L.; Chen, X. The progress of gut microbiome research related to brain disorders. J. Neuroinflammation 2020, 17, 25. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.S.; Greene, M.W.; Babu, J.R.; Frugé, A.D. Psychobiotics as treatment for anxiety, depression, and related symptoms: A systematic review. Nutr. Neurosci. 2019, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Vaghef-Mehrabany, E.; Maleki, V.; Behrooz, M.; Ranjbar, F.; Ebrahimi-Mameghani, M. Can psychobiotics "mood" ify gut? An update systematic review of randomized controlled trials in healthy and clinical subjects, on anti-depressant effects of probiotics, prebiotics, and synbiotics. Clin. Nutr. 2020, 39, 1395–1410. [Google Scholar] [CrossRef]
- Śliżewska, K.; Markowiak-Kopeć, P.; Żbikowski, A.; Szeleszczuk, P. The effect of synbiotic preparations on the intestinal microbiota and her metabolism in broiler chickens. Sci. Rep. 2020, 10, 4281. [Google Scholar] [CrossRef] [Green Version]
- Richards, P.J.; Lafontaine, G.M.F.; Connerton, P.L.; Liang, L.; Asiani, K.; Fish, N.M.; Connerton, I.F. Galacto-Oligosaccharides modulate the juvenile gut microbiome and innate immunity to improve broiler chicken performance. mSystems 2020, 5, e00827-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricke, S.C.; Lee, S.L.; Kim, S.A.; Park, S.H.; Shi, Z. Prebiotics and the poultry gastrointestinal tract microbiome. Poult. Sci. 2020, 99, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, R.; Mortada, M.; Cosby, D.E.; Singh, M.; Applegate, T.J.; Syed, B.; Pender, C.M.; Curry, S.; Murugesan, G.R.; Selvaraj, R.K. Synbiotic supplementation to decrease Salmonella colonization in the intestine and carcass contamination in broiler birds. PLoS ONE 2019, 14, e0223577. [Google Scholar] [CrossRef] [PubMed]
- Kridtayopas, C.; Rakangtong, C.; Bunchasak, C.; Loongyai, W. Effect of prebiotic and synbiotic supplementation in diet on growth performance, small intestinal morphology, stress, and bacterial population under high stocking density condition of broiler chickens. Poult. Sci. 2019, 98, 4595–4605. [Google Scholar] [CrossRef] [PubMed]
- Bogucka, J.; Ribeiro, D.M.; Bogusławska-Tryk, M.; Dankowiakowska, A.; da Costa, R.P.R.; Bednarczyk, M. Microstructure of the small intestine in broiler chickens fed a diet with probiotic or synbiotic supplementation. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1785–1791. [Google Scholar] [CrossRef] [PubMed]
- Rostagno, M.H. Effects of heat stress on the gut health of poultry. J. Anim. Sci. 2020, 98, skaa090. [Google Scholar] [CrossRef] [PubMed]
- Sandikci, M.; Eren, U.; Onol, A.G.; Kum, S. The effect of heat stress and the use of Saccharomyces cerevisiae or (and) bacitracin zinc against heat stress on the intestinal mucosa in quails. Revue. Méd. Vét. 2004, 155, 552–556. [Google Scholar]
- Sohail, M.U.; Hume, M.E.; Byrd, J.A.; Nisbet, D.J.; Shabbir, M.Z.; Ijaz, A.; Rehman, H. Molecular analysis of the caecal and tracheal microbiome of heat-stressed broilers supplemented with prebiotic and probiotic. Avian Pathol. 2015, 44, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Derovs, A.; Laivacuma, S.; Krumina, A. Targeting microbiota: What do we know about it at present? Medicina 2019, 55, 459. [Google Scholar] [CrossRef] [Green Version]
- Aviagen. Ross Broiler Management Handbook; Aviagen Inc.: Huntsville, AL, USA, 2018; Available online: www.aviagen.com (accessed on 4 June 2020).
- Mohammed, A.A.; Jacobs, J.A.; Murugesan, G.R.; Cheng, H.W. Effect of dietary synbiotic supplement on behavioral patterns and growth performance of broiler chickens reared under heat stress. Poult. Sci. 2018, 97, 1101–1108. [Google Scholar] [CrossRef]
- McCabe, B.J.; Horn, G. Learning and memory: Regional changes in N-methyl-d-aspartate receptors in the chick brain after imprinting. Proc. Natl. Acad. Sci. USA 1988, 85, 2979–2983. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.W.; Dillworth, G.; Singleton, P.; Chen, Y.; Muir, W.M. Effects of group selection for productivity and longevity on blood concentrations of serotonin, catecholamines, and corticosterone of laying hens. Poult. Sci. 2001, 80, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Campbell, T.W. Avian Hematology and Cytology, 2nd ed.; Iowa State University Press: Ames, IA, USA, 1995; p. 105. ISBN 0-8138-2970-4. [Google Scholar]
- Steel, R.; Torrie, J.; Dickey, D. Principles and Procedures of Statistics: A Biometrical Approach; McGraw Hill Book Co.: New York, NY, USA, 1997. [Google Scholar]
- Mack, L.A.; Felver-Grant, J.N.; Dennis, R.L.; Cheng, H.W. Genetic variation alters production and behavioral responses following heat stress in 2 strains of laying hens. Poult. Sci. 2013, 92, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Jiao, H.; Song, Z.; Zhao, J.; Wang, X.; Lin, H. Heat stress impairs mitochondria functions and induces oxidative injury in broiler chickens. J. Anim. Sci. 2015, 93, 2144–2153. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.C.; Yan, F.F.; Hu, J.H.; Amen, O.A.; Cheng, H.W. Supplementation of Bacillus subtilis-based probiotic reduces heat stress-related behaviors and inflammatory response in broiler chickens. J. Anim. Sci. 2018, 96, 1654–1666. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Mohammed, A.A.; Jacobs, J.A.; Cramer, T.A.; Cheng, H.W. Effect of synbiotics on thyroid hormones, intestinal histomorphology, and heat shock protein 70 expression in broiler chickens reared under cyclic heat stress. Poult. Sci. 2020, 99, 142–150. [Google Scholar] [CrossRef]
- Hirakawa, R.; Nurjanah, S.; Furukawa, K.; Murai, A.; Kikusato, M.; Nochi, T.; Toyomizu, M. Heat stress causes immune abnormalities via massive damage to effect proliferation and differentiation of lymphocytes in broiler chickens. Front. Vet. Sci. 2020, 7, 46. [Google Scholar] [CrossRef]
- Kim, H.G.; Kim, T.M.; Park, G.; Lee, T.H.; Oh, M.S. Repeated heat exposure impairs nigrostriatal dopaminergic neurons in mice. Biol. Pharm. Bull. 2013, 36, 1556–1561. [Google Scholar] [CrossRef] [Green Version]
- Gareau, M.G. Microbiota-gut-brain axis and cognitive function. Adv. Exp. Med. Biol 2014, 817, 3–24. [Google Scholar]
- Westfall, S.; Lomis, N.; Prakash, S. A novel synbiotic delays Alzheimer’s disease onset via combinatorial gut-brain-axis signaling in Drosophila melanogaster. PLoS ONE 2019, 14, e0214985. [Google Scholar] [CrossRef]
- Li, W.; Guo, J.; Shen, Y.; Shen, Y.; Huang, L.; Leng, B.; Fan, D.; Shui, L.; Chen, C. Probiotics, prebiotics, and synbiotics for the treatment of dementia: Protocol for a systematic review. Medicine 2020, 99, e18608. [Google Scholar] [CrossRef]
- Louzada, E.R.; Ribeiro, S.M.L. Synbiotic supplementation, systemic inflammation, and symptoms of brain disorders in elders: A secondary study from a randomized clinical trial. Nutr. Neurosci. 2020, 23, 93–100. [Google Scholar] [CrossRef]
- Freire, R.; Cheng, H.W. Experience dependent changes in hippocampus of domestic chicks: A model of spatial memory. Eur. J. Neurosci. 2004, 20, 1065–1068. [Google Scholar] [CrossRef] [PubMed]
- Romo-Araiza, A.; Gutiérrez-Salmeán, G.; Galván, E.J.; Hernández-Frausto, M.; Herrera-López, G.; Romo-Parra, H.; García-Contreras, V.; Fernández-Presas, A.M.; Jasso-Chávez, R.; Borlongan, C.V.; et al. Probiotics and prebiotics as a therapeutic strategy to improve memory in a model of middle-aged rats. Front. Aging Neurosci. 2018, 10, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Muir, W. The effects of genetic selection for survivability and productivity on chicken physiological homeostasis. World Poult. Sci. J. 2005, 61, 383–397. [Google Scholar] [CrossRef]
- Krause, E.T.; Phi-van, L.; Dudde, L.; Schrader, A.; Kjaer, J.B. Behavioural consequences of divergent selection on general locomotor activity in chickens. Behav. Process. 2019, 169, 103980. [Google Scholar] [CrossRef] [PubMed]
- Gualtieri, F.; Armstrong, E.A.; Longmoor, G.K.; D’Eath, R.B.; Sandilands, V.; Boswell, T.; Smulders, T.V. Unpredictable chronic mild stress suppresses the incorporation of new neurons at the caudal pole of the chicken hippocampal formation. Sci. Rep. 2019, 9, 7129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasdal, G.; Moe, R.O.; de Jong, I.C.; Granquist, E.G. The relationship between measures of fear of humans and lameness in broiler chicken flocks. Animal 2018, 12, 334–339. [Google Scholar] [CrossRef]
- Ghareeb, K.; Awad, W.A.; Nitsch, S.; Abdel-raheem, S.; Bohm, J. Effects of transportation on stress and fear responses of growing broilers supplemented with prebiotic or probiotic. Int. J. Poult. Sci. 2008, 7, 678–685. [Google Scholar] [CrossRef] [Green Version]
- Cowan, C.S.; Callaghan, B.L.; Richardson, R. The effects of a probiotic formulation (Lactobacillus rhamnosus and L. helveticus) on developmental trajectories of emotional learning in stressed infant rats. Transl. Psychiatry 2016, 6, e823. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.H.; Tsai, T.C.; Huang, W.Y.; Wu, H.M.; Hsu, K.S. Probiotic treatment restores normal developmental trajectories of fear memory retention in maternally separated infant rats. Neuropharmacology 2019, 153, 53–62. [Google Scholar] [CrossRef]
- Desbonnet, L.; Garrett, L.; Clarke, G.; Kiely, B.; Cryan, J.F.; Dinan, T.G. Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience 2010, 170, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Lyte, M. Microbial endocrinology: Host-microbiota neuroendocrine interactions influencing brain and behavior. Gut Microbes 2014, 5, 381–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall, R.; Cryan, J.F.; Paul Ross, R.; Fitzgerald, G.F.; Dinan, T.G.; Stanton, C. Bacterial neuroactive compounds produced by psychobiotics. Adv. Exp. Med. Biol. 2014, 17, 221–239. [Google Scholar]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef]
- Bocchio, M.; McHugh, S.B.; Bannerman, D.M.; Sharp, T.; Capogna, M. Serotonin, amygdala and fear: Assembling the puzzle. Front. Neural Circuits 2016, 10, 24. [Google Scholar] [CrossRef]
- Chauhan, N.R.; Kapoor, M.; Singh, L.P.; Gupta, R.K.; Meena, R.C.; Tulsawani, R.; Nanda, S.; Singh, S.B. Heat stress-induced neuroinflammation and aberration in monoamine levels in hypothalamus are associated with temperature dysregulation. Neuroscience 2017, 358, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Opoola, E.; Onimisi, P.A.; Ogundipe, S.O.; Bawa, G.S. Effect of dietary tryptophan levels on growth performance of broiler chickens reared in the hot season under tropical environment. Trop. Subtrop. Agroecosyst. 2017, 20, 429–437. [Google Scholar]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2014, 277, 32–48. [Google Scholar] [CrossRef]
- Hu, J.; Chen, H.; Cheng, H.W. Effect of direct-fed microbials, Bacillus subtilis, on production performance, serotonin concentrations and behavioral parameters in a selected dominant strain of white leghorn hens. Int. J. Poult. Sci. 2018, 17, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.D.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 2013, 18, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Höglund, E.; Overli, O.; Winberg, S. Tryptophan metabolic pathways and brain serotonergic activity: A Comparative review. Front. Endocrinol. 2019, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Tomonaga, S.; Okuyama, H.; Tachibana, T.; Makino, R. Effects of high ambient temperature on plasma metabolomic profiles in chicks. Anim. Sci. J. 2018, 89, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, A.M.; Tanabe, A.; Iwahori, Y. A systematic review of the effect of L-tryptophan supplementation on mood and emotional functioning. J. Diet. Suppl. 2020, 2020, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, C.E.; Martin, J.A.; Manriquez, F.V.; Dinan, T.G.; Cryan, J.F.; Clarke, G. Focus on the essentials: Tryptophan metabolism and the microbiome-gut-brain axis. Curr. Opin. Pharmacol. 2019, 48, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wang, L.; Li, G.; Cao, C.; Fang, R.; Liu, P.; Luo, S.; Zhang, X. Correlation between hypothalamic-pituitary-adrenal axis gene polymorphisms and posttraumatic stress disorder symptoms. Horm. Behav. 2020, 117, 104604. [Google Scholar] [CrossRef]
- Ghareeb, K.; Awad, W.A.; Sid-Ahmed, O.E.; Böhm, J. Insights on the host stress, fear and growth responses to the deoxynivalenol feed contaminant in broiler chickens. PLoS ONE 2014, 9, e87727. [Google Scholar] [CrossRef]
- Van de Kar, L.D.; Blair, M.L. Forebrain pathways mediating stress-induced hormone secretion. Front. Neuroendocrinol. 1999, 20, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Tong, Q.; Zheng, W.; Tu, J.; Li, B. Effects of nest boxes in natural mating colony cages on fear, stress, and feather damage for layer breeders. J. Anim. Sci. 2019, 97, 4464–4474. [Google Scholar] [CrossRef]
- Peixoto, M.R.L.V.; Karrow, N.A.; Newman, A.; Widowski, T.M. Effects of maternal stress on measures of anxiety and fearfulness in different strains of laying hens. Front. Vet. Sci. 2020, 7, 128. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, H.A.; Purswell, J.L.; Collier, S.D.; Branton, S.L. Effect of ambient temperature and light intensity on physiological reactions of heavy broiler chickens. Poult. Sci. 2010, 89, 2668–2677. [Google Scholar] [CrossRef]
- Scanes, C.G. Biology of stress in poultry with emphasis on glucocorticoids and the heterophil to lymphocyte ratio. Poult. Sci. 2016, 95, 2208–2215. [Google Scholar] [CrossRef] [PubMed]
- Cengiz, Ö.; Köksal, B.K.; Tatlı, O.; Sevim, Ö.; Ahsan, U.; Üner, A.G.; Ulutaş, P.A.; Beyaz, D.; Büyükyörük, S.; Yakan, A.; et al. Effect of dietary probiotic and high stocking density on the performance, carcass yield, gut microflora, and stress indicators of broilers. Poult. Sci. 2015, 94, 2395–2403. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.A.; Jiang, S.; Jacobs, J.A.; Cheng, H.W. Effect of a synbiotic supplement on cecal microbial ecology, antioxidant status, and immune response of broiler chickens reared under heat stress. Poult. Sci. 2019, 98, 4408–4415. [Google Scholar] [CrossRef] [PubMed]
- Emadi, M.; Kermanshahi, H. Effect of turmeric rhizome powder on the activity of some blood enzymes in broiler chickens. Int. J. Poult. Sci. 2007, 6, 48–51. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient % | Starter (1–14 day) | Grower (15–28 day) | Finisher (29–42 day) |
|---|---|---|---|
| Corn ground | 57.66 | 63.76 | 66.9 |
| Soybean meal 47.5% | 35.27 | 29.68 | 26.3 |
| Soybean oil degummed | 3 | 3 | 3.52 |
| Calcium carbonate | 1.41 | 1.38 | 1.49 |
| Phosphate monocalcium | 1.42 | 1.02 | 0.82 |
| L-Lysine | 0.11 | 0.1 | 0.02 |
| Salt plain | 0.48 | 0.46 | 0.48 |
| L-Threonine 98% | 0.06 | 0.04 | 0 |
| DL-Methionine | 0.24 | 0.21 | 0.12 |
| Poultry turkey starter | 0.35 | 0.35 | 0.35 |
| Calculated Analysis 3 | |||
| Crude protein % | 23.4 | 22.8 | 19.2 |
| Poultry ME kcal/kg | 3050 | 3151 | 3200 |
| Calcium % | 0.95 | 0.85 | 0.75 |
| Available phosphorus % | 0.50 | 0.44 | 0.36 |
| Methionine % | 0.66 | 0.59 | 0.53 |
| Methionine+Cystine % | 1.04 | 0.97 | 0.86 |
| Lysine % | 1.42 | 1.29 | 1.09 |
| Threonine % | 0.97 | 0.89 | 0.74 |
| Na % | 0.22 | 0.20 | 0.19 |
| Treatments 1 | G-C | G-0.5 | G-1.0 | SEM | p-Value |
|---|---|---|---|---|---|
| Novel object test 2 | |||||
| Number of birds within 30 cm from the object | 25.87 | 22.50 | 25.50 | 2.67 | 0.627 |
| Touch test 2 | |||||
| Number of touched birds | 25.12 b | 33.62 b | 78.87 a | 4.83 | 0.001 |
| Tonic immobility test 3 | |||||
| Latency of bird to right itself (min) | 5.50 | 4.07 | 3.55 | 0.74 | 0.185 |
| Treatments 1 | G-C | G-0.5 | G-1.0 | SEM | p-Value |
|---|---|---|---|---|---|
| H/L ratio | 0.79 a | 0.58 b | 0.43 c | 0.02 | 0.001 |
| Corticosterone (ng/mL) | 2.37 | 2.19 | 2.24 | 0.06 | 0.124 |
| Tryptophan (µmol/L) | 103.62 b | 104.87 ab | 107.50 a | 1.03 | 0.043 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammed, A.; Mahmoud, M.; Murugesan, R.; Cheng, H.-w. Effect of a Synbiotic Supplement on Fear Response and Memory Assessment of Broiler Chickens Subjected to Heat Stress. Animals 2021, 11, 427. https://doi.org/10.3390/ani11020427
Mohammed A, Mahmoud M, Murugesan R, Cheng H-w. Effect of a Synbiotic Supplement on Fear Response and Memory Assessment of Broiler Chickens Subjected to Heat Stress. Animals. 2021; 11(2):427. https://doi.org/10.3390/ani11020427
Chicago/Turabian StyleMohammed, Ahmed, Manal Mahmoud, Raj Murugesan, and Heng-wei Cheng. 2021. "Effect of a Synbiotic Supplement on Fear Response and Memory Assessment of Broiler Chickens Subjected to Heat Stress" Animals 11, no. 2: 427. https://doi.org/10.3390/ani11020427
APA StyleMohammed, A., Mahmoud, M., Murugesan, R., & Cheng, H.-w. (2021). Effect of a Synbiotic Supplement on Fear Response and Memory Assessment of Broiler Chickens Subjected to Heat Stress. Animals, 11(2), 427. https://doi.org/10.3390/ani11020427