
Effects of Selected Prebiotics or Synbiotics Administered in ovo on Lymphocyte Subsets in Bursa of the Fabricius, Thymus, and Spleen in Non-Immunized and Immunized Chicken Broilers

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. In Ovo Treatment
2.2. Post Hatch Treatment of Animals
2.3. Immunization
2.4. Measurements
2.5. Statistical Analysis
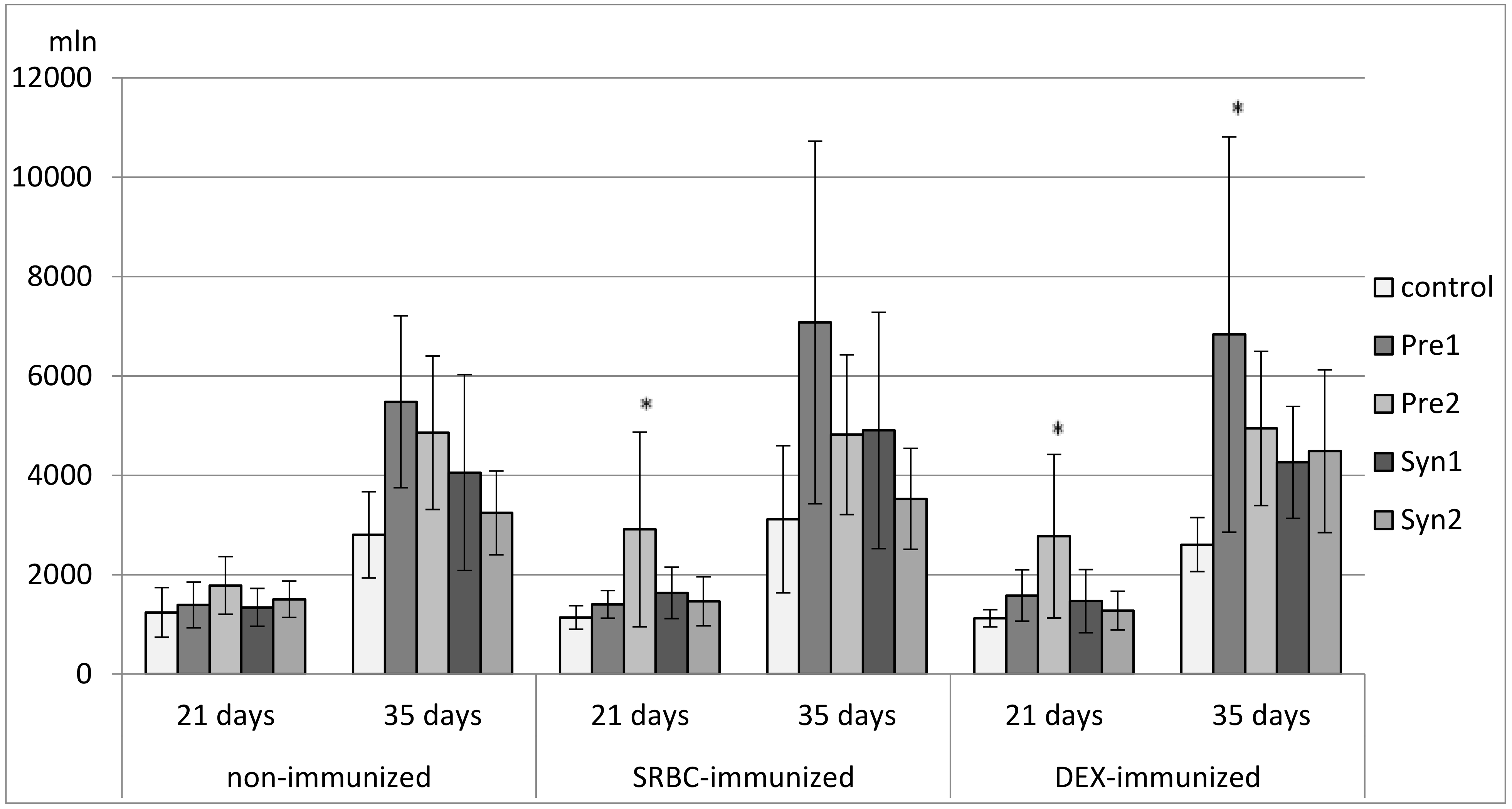
3. Results
3.1. The Effects of Prebiotics and Synbiotics on the Subsets of Bursal Lymphocytes
3.2. The Effects of Prebiotics and Synbiotics on the Subsets of Thymocytes
3.3. The Effects of Prebiotics and Synbiotics on the Subsets of Splenocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mookiah, S.; Sieo, C.C.; Ramasamy, K.; Abdullah, N.; Ho, Y.W. Effects of dietary prebiotics, probiotic and synbiotics on performance, caecal bacterial populations and caecal fermentation concentrations of broiler chickens. J. Sci. Food Agric. 2014, 94, 341–348. [Google Scholar] [CrossRef]
- Patterson, J.A.; Burkholder, K.M. Application of prebiotics and probiotics in poultry production. Poult. Sci. 2003, 82, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Ishii, Y.; Arakawa, K.; Uemura, K.; Saitoh, B.; Nishimura, J.; Kitazawa, H.; Yamazaki, Y.; Tateno, Y.; Itoh, T.; et al. Structural and functional differences in two cyclic bacteriocins with the same sequences produced by lactobacilli. Appl. Environ. Microbiol. 2004, 70, 2906–2911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santini, C.; Baffoni, L.; Gaggia, F.; Granata, M.; Gasbarri, R.; Di Gioia, D.; Biavati, B. Characterization of probiotic strains: An application as feed additives in poultry against Campylobacter jejuni. Int. J. Food Microbiol. 2010, 141 (Suppl. 1), S98–S108. [Google Scholar] [CrossRef] [PubMed]
- Brisbin, J.T.; Gong, J.; Sharif, S. Interactions between commensal bacteria and the gut-associated immune system of the chicken. Anim. Health Res. Rev. 2008, 9, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, K.-J.; Sethupathi, P.; Driks, A.; Lanning, D.K.; Knight, K.L. Role of Commensal Bacteria in Development of Gut-Associated Lymphoid Tissues and Preimmune Antibody Repertoire. J. Immunol. 2004, 172, 1118–1124. [Google Scholar] [CrossRef] [Green Version]
- Baurhoo, B.; Ferket, P.R.; Zhao, X. Effects of diets containing different concentrations of mannanoligosaccharide or antibiotics on growth performance, intestinal development, cecal and litter microbial populations, and carcass parameters of broilers. Poult. Sci. 2009, 88, 2262–2272. [Google Scholar] [CrossRef]
- Baurhoo, B.; Phillip, L.; Ruiz-Feria, C.A. Effects of Purified Lignin and Mannan Oligosaccharides on Intestinal Integrity and Microbial Populations in the Ceca and Litter of Broiler Chickens. Poult. Sci. 2007, 86, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.-B.; Seo, Y.M.; Kim, C.H.; Paik, I.K. Effect of dietary prebiotic supplementation on the performance, intestinal microflora, and immune response of broilers. Poult. Sci. 2011, 90, 75–82. [Google Scholar] [CrossRef]
- Janardhana, V.; Broadway, M.M.; Bruce, M.P.; Lowenthal, J.W.; Geier, M.S.; Hughes, R.J.; Bean, A.G.D. Prebiotics Modulate Immune Responses in the Gut-Associated Lymphoid Tissue of Chickens. J. Nutr. 2009, 139, 1404–1409. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Abdel-Raheem, S.; Böhm, J. Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens. Poult. Sci. 2009, 88, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Alzueta, C.; Rodríguez, M.; Ortiz, L.; Rebolé, A.; Trevino, J. Effects of inulin on growth performance, nutrient digestibility and metabolisable energy in broiler chickens. Br. Poult. Sci. 2010, 51, 393–398. [Google Scholar] [CrossRef]
- Erdogan, Z.; Aslantas, O.; Çelik, S. Original Article: Effects of dietary supplementation of synbiotics and phytobiotics on performance, caecal coliform population and some oxidant/antioxidant parameters of broilers. J. Anim. Physiol. Anim. Nutr. 2010, 94, e40–e48. [Google Scholar] [CrossRef]
- Houshmand, M.; Azhar, K.; Zulkifli, I.; Bejo, M.; Kamyab, A. Effects of prebiotic, protein level, and stocking density on performance, immunity, and stress indicators of broilers. Poult. Sci. 2012, 91, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Bednarczyk, M.; Urbanowski, M.; Gulewicz, P.; Kasperczyk, K.; Maiorano, G.; Szwaczkowski, T. Field and in vitro study on prebiotic effect of raffinose family oligosaccharides in chickens. Bull. Vet. Inst. Pulawy 2011, 55, 465–469. [Google Scholar]
- Siwek, M.; Slawinska, A.; Stadnicka, K.; Bogucka, J.; Dunislawska, A.; Bednarczyk, M. Prebiotics and synbiotics—In ovo delivery for improved lifespan condition in chicken. BMC Vet. Res. 2018, 14, 402. [Google Scholar] [CrossRef] [PubMed]
- Pilarski, R.; Bednarczyk, M.; Lisowski, M.; Rutkowski, A.; Bernacki, Z.; Wardeńska, M.; Gulewicz, K. Assessment of the effect of α-galactosides injected during embryogenesis on selected chicken traits. Folia Biol. 2005, 53, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villaluenga, C.M.; Wardeńska, M.; Pilarski, R.; Bednarczyk, M.; Gulewicz, K. Utilization of the chicken embryo model for assessment of biological activity of different oligosaccharides. Folia Biol. 2004, 52, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Madej, J.; Bednarczyk, M. Effect of in ovo- delivered prebiotics and synbiotics on the morphology and specific immune cell composition in the gut-associated lymphoid tissue. Poult. Sci. 2016, 95, 19–29. [Google Scholar] [CrossRef]
- Madej, J.; Stefaniak, T.; Bednarczyk, M. Effect of in ovo-delivered prebiotics and synbiotics on lymphoid-organs’ morphology in chickens. Poult. Sci. 2015, 94, 1209–1219. [Google Scholar] [CrossRef]
- Stefaniak, T.; Madej, J.; Graczyk, S.; Siwek, M.; Łukaszewicz, E.; Kowalczyk, A.; Sieńczyk, M.; Bednarczyk, M. Selected prebiotics and synbiotics administered in ovo can modify innate immunity in chicken broilers. BMC Vet. Res. 2019, 15, 105. [Google Scholar] [CrossRef]
- Stefaniak, T.; Madej, J.; Graczyk, S.; Siwek, M.; Łukaszewicz, E.; Kowalczyk, A.; Sieńczyk, M.; Maiorano, G.; Bednarczyk, M. Impact of Prebiotics and Synbiotics Administered in ovo on the Immune Response against Experimental Antigens in Chicken Broilers. Animals 2020, 10, 643. [Google Scholar] [CrossRef] [Green Version]
- Sławińska, A.; Siwek, M.; Żylińska, J.; Bardowski, J.; Brzezińska, J.; Gulewicz, K.A.; Nowak, M.; Urbanowski, M.; Płowiec, A.; Bednarczyk, M. Influence of Synbiotics Delivered in ovo on Immune Organs Development and Structure. Folia Biol. 2014, 62, 277–285. [Google Scholar] [CrossRef]
- Alkie, T.N.; Yitbarek, A.; Hodgins, D.C.; Kulkarni, R.; Taha-Abdelaziz, K.; Sharif, S. Development of innate immunity in chicken embryos and newly hatched chicks: A disease control perspective. Avian Pathol. 2019, 48, 288–310. [Google Scholar] [CrossRef]
- Berndt, A.; Pieper, J.; Methner, U. Circulating γδ T Cells in Response to Salmonellaenterica Serovar Enteritidis Exposure in Chickens. Infect. Immun. 2006, 74, 3967–3978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamura, M.; Lillehoj, H.S.; Raybourne, R.; Babu, U.; Heckert, R. Cell-mediated immune responses to a killed Salmonella enteritidis vaccine: Lymphocyte proliferation, T-cell changes and interleukin-6 (IL-6), IL-1, IL-2, and IFN-γ production. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Kjærup, R.M.; Dalgaard, T.S.; Norup, L.R.; Hamzić, E.; Sørensen, P.; Juul-Madsen, H.R. Characterization of Cellular and Humoral Immune Responses After IBV Infection in Chicken Lines Differing in MBL Serum Concentration. Viral Immunol. 2014, 27, 529–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratcliffe, M.J.H. B cells, the bursa of Fabricius and the generation of antibody repertoires. In Avian Immunology; Davison, F., Kaspers, B., Schat, K.A., Eds.; Academic Press: London, UK, 2008; pp. 67–89. [Google Scholar]
- Sato, K.; Takahashi, K.; Tohno, M.; Miura, Y.; Kamada, T.; Ikegami, S.; Kitazawa, H. Immunomodulation in gut-associated lymphoid tissue of neonatal chicks by immunobiotic diets. Poult. Sci. 2009, 88, 2532–2538. [Google Scholar] [CrossRef]
- Nabizadeh, A. The effect of inulin on broiler chicken intestinal microflora, gut morphology, and performance. J. Anim. Feed. Sci. 2012, 21, 725–734. [Google Scholar] [CrossRef]
- Wu, X.; Wen, Z.; Hua, J. Effects of dietary inclusion of Lactobacillus and inulin on growth performance, gut microbiota, nutrient utilization, and immune parameters in broilers. Poult. Sci. 2019, 98, 4656–4663. [Google Scholar] [CrossRef]
- Madej, J.; Skonieczna, J.; Siwek, M.; Kowalczyk, A.; Łukaszewicz, E.; Slawinska, A. Genotype-dependent development of cellular and humoral immunity in the spleen and cecal tonsils of chickens stimulated in ovo with bioactive compounds. Poult. Sci. 2020, 99, 4343–4350. [Google Scholar] [CrossRef]
- Alizadeh, M.; Shojadoost, B.; Astill, J.; Taha-Abdelaziz, K.; Karimi, S.H.; Bavananthasivam, J.; Kulkarni, R.R.; Sharif, S. Effects of in ovo Inoculation of Multi-Strain Lactobacilli on Cytokine Gene Expression and Antibody-Mediated Immune Responses in Chickens. Front. Vet. Sci. 2020, 7, 105. [Google Scholar] [CrossRef] [Green Version]
- Flaujac Lafontaine, G.M.; Richards, P.J.; Connerton, P.L.; O’Kane, P.M.; Ghaffar, N.M.; Cummings, N.J.; Fish, N.M.; Connerton, I.F. Prebiotic Driven Increases in IL-17A Do Not Prevent Campylobacter jejuni Colonization of Chickens. Front. Microbiol. 2020, 10, 3030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sławinska, A.; Siwek, M.Z.; Bednarczyk, M.F. Effects of synbiotics injected in ovo on regulation of immune-related gene expression in adult chickens. Am. J. Vet. Res. 2014, 75, 997–1003. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. The role of probiotics, prebiotics and synbiotics in animal nutrition. Gut Pathog. 2018, 10, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Shukla, R.; Goyal, A. Probiotics in valorization of innate immunity across various animal models. J. Funct. Foods 2015, 14, 549–561. [Google Scholar] [CrossRef]
- Ng, S.C.; Hart, A.L.; Kamm, M.A.; Stagg, A.J.; Knight, S.C. Mechanisms of action of probiotics: Recent advances. Inflamm. Bowel Dis. 2009, 15, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Broom, L.J.; Kogut, M.H. Gut immunity: Its development and reasons and opportunities for modulation in monogastric production animals. Anim. Health Res. Rev. 2018, 19, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Casteleyn, C.; Doom, M.; Lambrechts, E.; Broeck, W.V.D.; Simoens, P.; Cornillie, P. Locations of gut-associated lymphoid tissue in the 3-month-old chicken: A review. Avian Pathol. 2010, 39, 143–150. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day of Experiment | Groups Injected In Ovo with Bioactive Components | Control Group | Pre1 Group | Pre2 Group | Syn1 Group | Syn2 Group | Procedure |
|---|---|---|---|---|---|---|---|
| Numbers of Chickens/Groups | |||||||
| 0 | 49 | 49 | 49 | 49 | 49 | ||
| 7 first immunization | Non-immunized | −7 | −7 | −7 | −7 | −7 | chickens randomly selected and euthanized |
| Non-immunized | 14 | 14 | 14 | 14 | 14 | chickens randomly selected | |
| SRBC-immunized | 14 | 14 | 14 | 14 | 14 | chickens randomly selected and immunized | |
| DEX-immunized | 14 | 14 | 14 | 14 | 14 | chickens randomly selected and immunized | |
| 21 second immunization | Non- immunized | −7 | −7 | −7 | −7 | −7 | chickens randomly selected and euthanized |
| SRBC-immunized | −7 | −7 | −7 | −7 | −7 | chickens randomly selected and euthanized | |
| DEX-immunized | −7 | −7 | −7 | −7 | −7 | chickens randomly selected and euthanized | |
| Non- immunized | 7 | 7 | 7 | 7 | 7 | chickens randomly selected | |
| SRBC-immunized | 7 | 7 | 7 | 7 | 7 | chickens randomly selected and immunized | |
| DEX-immunized | 7 | 7 | 7 | 7 | 7 | chickens randomly selected and immunized | |
| 35 | Non- immunized | −7 | −7 | −7 | −7 | −7 | euthanized chickens |
| SRBC-immunized | −7 | −7 | −7 | −7 | −7 | euthanized chickens | |
| DEX-immunized | −7 | −7 | −7 | −7 | −7 | euthanized chickens | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczypka, M.; Suszko-Pawłowska, A.; Kuczkowski, M.; Gorczykowski, M.; Lis, M.; Kowalczyk, A.; Łukaszewicz, E.; Poradowski, D.; Zbyryt, I.; Bednarczyk, M.; et al. Effects of Selected Prebiotics or Synbiotics Administered in ovo on Lymphocyte Subsets in Bursa of the Fabricius, Thymus, and Spleen in Non-Immunized and Immunized Chicken Broilers. Animals 2021, 11, 476. https://doi.org/10.3390/ani11020476
Szczypka M, Suszko-Pawłowska A, Kuczkowski M, Gorczykowski M, Lis M, Kowalczyk A, Łukaszewicz E, Poradowski D, Zbyryt I, Bednarczyk M, et al. Effects of Selected Prebiotics or Synbiotics Administered in ovo on Lymphocyte Subsets in Bursa of the Fabricius, Thymus, and Spleen in Non-Immunized and Immunized Chicken Broilers. Animals. 2021; 11(2):476. https://doi.org/10.3390/ani11020476
Chicago/Turabian StyleSzczypka, Marianna, Agnieszka Suszko-Pawłowska, Maciej Kuczkowski, Michał Gorczykowski, Magdalena Lis, Artur Kowalczyk, Ewa Łukaszewicz, Dominik Poradowski, Iwona Zbyryt, Marek Bednarczyk, and et al. 2021. "Effects of Selected Prebiotics or Synbiotics Administered in ovo on Lymphocyte Subsets in Bursa of the Fabricius, Thymus, and Spleen in Non-Immunized and Immunized Chicken Broilers" Animals 11, no. 2: 476. https://doi.org/10.3390/ani11020476