
Insights on Immune Function in Free-Ranging Green Sea Turtles (Chelonia mydas) with and without Fibropapillomatosis

 ,
,  , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Sites
2.1.1. Big Bend Region, Florida
2.1.2. Jennings Cove, Indian River Lagoon, Florida
2.1.3. Jupiter Inlet, Lake Worth Lagoon, Florida
2.2. Animal Capture and Sample Collection and Processing
2.3. Molecular Diagnostics for ChHV5 and ChHV6 DNA
2.4. ELISA Detection of Antibodies to ChHV5 and ChHV6 Peptides
2.5. Isolation of Peripheral Blood Mononuclear Cells
2.6. Lymphocyte Proliferation
2.7. Natural Killer Cell Activity
2.8. Statistical Analyses
3. Results
3.1. Physical Examination and Morphometrics
3.2. Fibropapilloma Tumor Score and Severity
3.3. Molecular Detection of ChHV5 and ChHV6 DNA and Antibodies
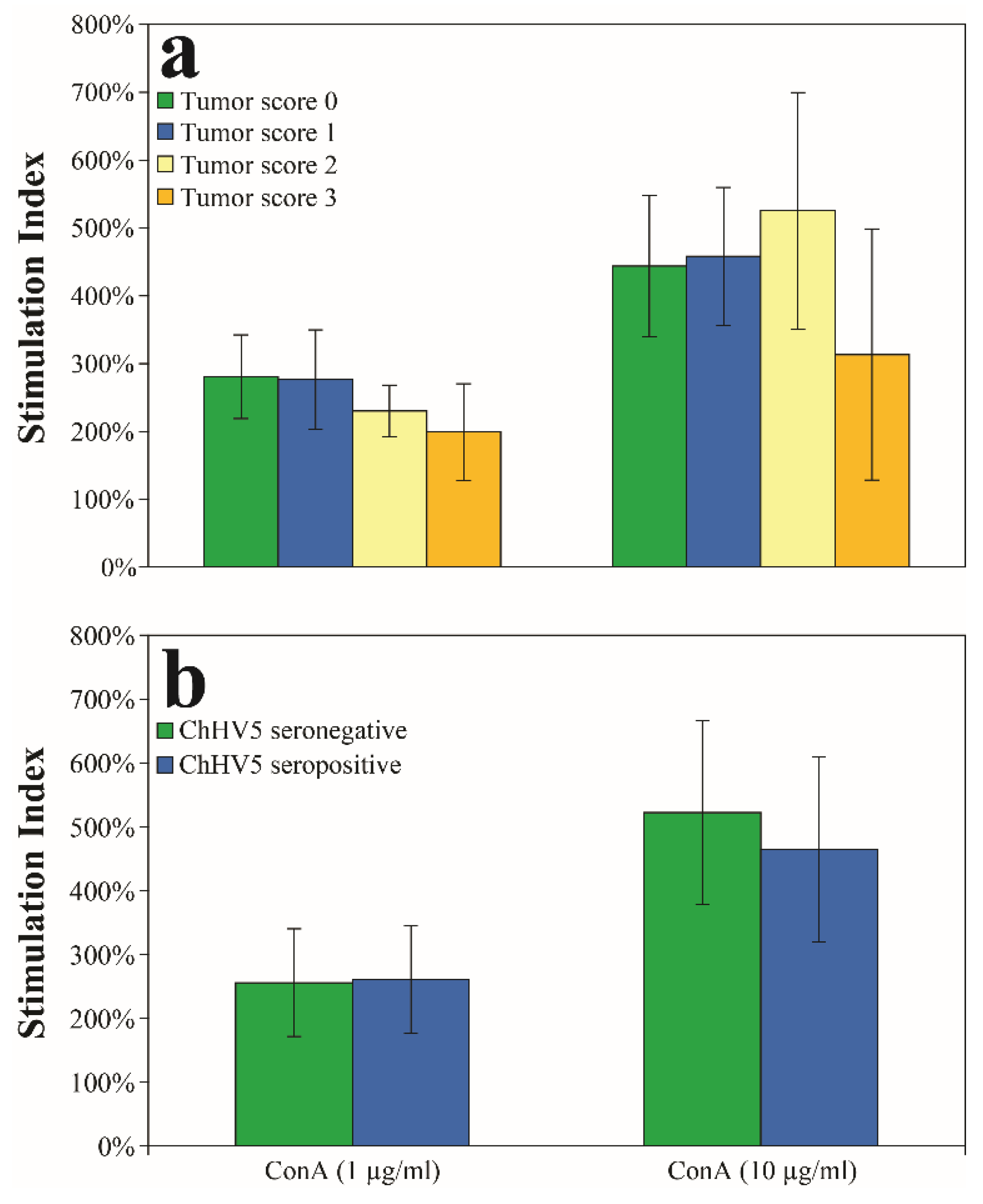
3.4. Relationships of Fibropapillomatosis and ChHV5 Seroprevalence to Lymphocyte Proliferation Assays
3.5. Relationship of Fibropapillomatosis to Natural Killer Cell Activity
4. Discussion
4.1. Fibropapillomatosis in Green Turtles from Florida
4.2. Diagnostic Assays for ChHV5 and ChHV6
4.3. Lymphocyte Proliferation
4.4. Natural Killer Cell Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; et al. Historical overfishing and the recent collapse of coastal ecosystems. Science 2001, 293, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, A.A.; Lutz, P.L. Marine turtles as sentinels of ecosystem health: Is fibropapillomatosis an indicator? EcoHealth 2004, 1, 275–283. [Google Scholar] [CrossRef]
- Chaloupka, M.; Bjorndal, K.A.; Balazs, G.H.; Bolten, A.B.; Ehrhart, L.M.; Limpus, C.J.; Suganuma, H.; Troëng, S.; Yamaguchi, M. Encouraging outlook for recovery of a once severely exploited marine megaherbivore. Glob. Ecol. Biogeogr. 2008, 17, 297–304. [Google Scholar] [CrossRef]
- Page-Karjian, A.; Chabot, R.; Stacy, N.I.; Morgan, A.S.; Valverde, R.A.; Stewart, S.; Coppenrath, C.M.; Manire, C.A.; Herbst, L.H.; Gregory, C.R.; et al. Comprehensive health assessment of green turtles Chelonia mydas nesting in southeastern Florida, USA. Endang. Spec. Res. 2020, 42, 21–35. [Google Scholar] [CrossRef]
- Thayer, G.W.; Bjorndal, K.A.; Ogden, J.C.; Williams, S.L.; Zieman, J.C. Role of larger herbivores in seagrass communities. Estuaries 1984, 7, 351–376. [Google Scholar] [CrossRef]
- Bjorndal, K.A.; Bolten, A.B.; Chaloupka, M.Y. Green turtle somatic growth model: Evidence for density dependence. Ecol. Appl. 2000, 10, 269–282. [Google Scholar]
- Witherington, B.E.; Bresette, M.J.; Herren, R. Chelonia mydas–Green turtle. In Biology and Conservation of Florida Turtles; Meylan, P.A., Ed. Chelonian Res. Monogr. 2006, 3, 90–104. [Google Scholar]
- Valdivia, A.; Wolf, S.; Suckling, K. Marine mammals and sea turtles listed under the U.S. Endangered Species Act are recovering. PLoS ONE 2019, 14, e0210164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrhart, L.M.; Redfoot, W.E.; Bagley, D.A. Marine turtles of the central region of the Indian River Lagoon System, Florida. Fla. Sci. 2007, 70, 415–434. [Google Scholar]
- Seminoff, J.A.; Allen, C.D.; Balazs, G.H.; Dutton, P.H.; Eguchi, T.; Haas, H.L.; Hargrove, S.A.; Jensen, M.; Klemm, D.L.; Lauritsen, A.M.; et al. Status Review of the Green Turtle (Chelonia mydas) under the Endangered Species Act. NOAA Tech Memo NOAA-NMFS-SWFSC-539; NOAA Tech Memo: La Jolla, CA, USA, 2015; p. 571.
- Brost, B.; Witherington, B.; Meylan, A.; Leone, E.; Ehrhart, L.; Bagley, D. Sea turtle hatchling production from Florida (USA) beaches, 2002–2012, with recommendations for analyzing hatching success. Endang. Spec. Res. 2015, 27, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Florida Fish and Wildlife Conservation Commission (FWC). Index Nesting Beach Survey Totals (1989–2019). 2020. Available online: https://myfwc.com/research/wildlife/sea-turtles/nesting/beach-survey-totals/ (accessed on 30 October 2020).
- Foley, A.M.; Schroeder, B.A.; Redlow, A.E.; Fick-Child, K.J.; Teas, W.G. Fibropapillomatosis in stranded green turtles (Chelonia mydas) from the eastern United States (1980–1998): Trends and associations with environmental factors. J. Wildl. Dis. 2005, 41, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirama, S.; Ehrhart, L.M. Description, prevalence and severity of green turtle fibropapillomatosis in three developmental habitats on the east coast of Florida. Fla. Sci. 2007, 70, 435–448. [Google Scholar]
- Hirama, S.; Ehrhart, L.M.; Rea, L.D.; Kiltie, R.A. Relating fibropapilloma tumor severity to blood parameters in green turtles Chelonia mydas. Dis. Aquat. Org. 2014, 111, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Gorham, J.C.; Bresette, M.J.; Guertin, J.R.; Shamblin, B.M.; Nairn, C.J. Green turtles (Chelonia mydas) in an urban estuary system: Lake Worth Lagoon, Florida. Fla. Sci. 2016, 79, 14–27. [Google Scholar]
- Perrault, J.R.; Stacy, N.I.; Lehner, A.F.; Mott, C.R.; Hirsch, S.; Gorham, J.C.; Buchweitz, J.P.; Bresette, M.J.; Walsh, C.J. Potential effects of brevetoxins and toxic elements on various health variables in Kemp’s ridley (Lepidochelys kempii) and green (Chelonia mydas) sea turtles after a red tide bloom event. Sci. Total Environ. 2017, 605–606, 967–979. [Google Scholar] [CrossRef]
- Chabot, R.M.; Welsh, R.C.; Mott, C.R.; Guertin, J.R.; Shamblin, B.M.; Witherington, B.E. A sea turtle population assessment for Florida’s Big Bend, northeastern Gulf of Mexico. Gulf Caribb. Res. 2021, in press. [Google Scholar]
- Herbst, L.H. Fibropapillomatosis of marine turtles. Ann. Rev. Fish. Dis. 1994, 4, 389–425. [Google Scholar] [CrossRef]
- Herbst, L.H.; Klein, P.A. Green turtle fibropapillomatosis: Challenges to assessing the role of environmental cofactors. Environ. Health Perspect. 1995, 103, 27–30. [Google Scholar]
- Herbst, L.H.; Jacobson, E.R.; Moretti, R.; Brown, T.; Sundberg, J.P.; Klein, P.A. Experimental transmission of green turtle fibropapillomatosis using cell-free tumor extracts. Dis. Aquat. Org. 1995, 22, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Landsberg, J.H.; Balazs, G.H.; Steidinger, K.A.; Baden, D.G.; Work, T.M.; Russell, D.J. The potential role of natural tumor promoters in marine turtle fibropapillomatosis. J. Aquat. Anim. Health 1999, 11, 199–210. [Google Scholar] [CrossRef]
- Arthur, K.; Limpus, C.; Balazs, G.; Capper, A.; Udy, J.; Shaw, G.; Keuper-Bennett, U.; Bennett, P. The exposure of green turtles (Chelonia mydas) to tumour promoting compounds produced by the cyanobacterium Lyngbya majuscula and their potential role in the aetiology of fibropapillomatosis. Harmful Algae 2008, 7, 114–125. [Google Scholar] [CrossRef]
- dos Santos, R.G.; Martins, A.S.; Torezani, E.; Baptistotte, C.; Farias, J.d.N.; Horta, P.A.; Work, T.M.; Balazs, G.H. Relationship between fibropapillomatosis and environmental quality: A case study with Chelonia mydas off Brazil. Dis. Aquat. Org. 2010, 89, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, C.C.; Klein, R.D.; Barcarolli, I.F.; Bianchini, A. Metal contamination as a possible etiology of fibropapillomatosis in juvenile female green sea turtles Chelonia mydas from the southern Atlantic Ocean. Aquat. Toxicol. 2016, 170, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Perrault, J.R.; Perkins, C.R.; Ajemian, M.J.; Bresette, M.J.; Mott, C.R.; Page-Karjian, A. Harmful algal and cyanobacterial toxins in foraging green turtles (Chelonia mydas) in Florida’s Big Bend. Toxicon X 2020, 5, 100020. [Google Scholar] [CrossRef]
- Duffy, D.J.; Schnitzler, C.; Karpinski, L.; Thomas, R.; Whilde, J.; Eastman, C.; Yang, C.; Krstic, A.; Rollinson, D.; Zirkelbach, B.; et al. Sea turtle fibropapilloma tumors share genomic drivers and therapeutic vulnerabilities with human cancers. Commun. Biol. 2018, 1, 63. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.M.; Coates, C.W. Fibro-epithelial growths of the skin in large marine turtles, Chelonia mydas (Linnaeus). Zoologica 1938, 23, 93–98. [Google Scholar]
- Work, T.M.; Balazs, G.H. Relating tumor score to hematology in green turtles with fibropapillomatosis in Hawaii. J. Wildl. Dis. 1999, 35, 804–807. [Google Scholar] [CrossRef] [Green Version]
- Limpus, C.J.; Couper, P.J.; Couper, K.L.D. Crab Island revisited: Reassessment of the world’s largest flatback turtle rookery after twelve years. Mem. Qld. Mus. 1993, 33, 227–289. [Google Scholar]
- Barragan, A.R.; Sarti, M.L. A possible case of fibropapilloma in Kemp’s ridley turtle (Lepidochelys kempii). Mar. Turtle Newsl. 1994, 67, 27. [Google Scholar]
- Quackenbush, S.L.; Work, T.M.; Balazs, G.H.; Casey, R.N.; Rovnak, A.; Chaves, A.; duToit, L.; Baines, C.R.; Parrish, C.R.; Bowser, P.R.; et al. Three closely related herpesviruses are associated with fibropapillomatosis in marine turtles. Virology 1998, 246, 392–399. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, A.A.; Spraker, T.R.; Chaves, A.; Toit, L.; Eure, W.; Balazs, G.H. Pathology of fibropapillomatosis in olive ridley turtles Lepidochely olivacea nesting in Costa Rica. J. Aquat. Health 1999, 11, 283–289. [Google Scholar] [CrossRef]
- D’Amato, A.F.; Moraes-Neto, M. First documentation of fibropapillomas verified by histopathology in Eretmochelys imbricata. Mar. Turt. Newsl. 2000, 89, 12–13. [Google Scholar]
- Page-Karjian, A.; Norton, T.M.; Harms, C.; Mader, D.; Herbst, L.H.; Stedman, N.; Gottdenker, N.L. Case descriptions of fibropapillomatosis in rehabilitating loggerhead sea turtles Caretta caretta in the southeastern USA. Dis. Aquat. Org. 2015, 115, 185–191. [Google Scholar] [CrossRef]
- Jones, K.; Ariel, E.; Burgess, G.; Read, M. A review of fibropapillomatosis in green turtles (Chelonia mydas). Vet. J. 2016, 212, 48–57. [Google Scholar] [CrossRef]
- Díaz-Delgado, J.; Gomes-Borges, J.C.; Silveira, A.M.; Einhardt-Vergara, J.; Groch, K.R.; Cirqueira, C.S.; Sansone, M.; Gattamorta, M.A.; Matushima, E.R.; Catao-Dias, J.L. Primary multicentric pulmonary low-grade fibromyxoid sarcoma and chelonid alphaherpesvirus 5 detection in a leatherback sea turtle (Dermochelys coriacea). Comp. Pathol. 2019, 168, 1–7. [Google Scholar] [CrossRef]
- Herbst, L.H.; Jacobson, E.R.; Klein, P.A.; Balazs, G.H.; Moretti, R.; Brown, T.; Sundberg, J.P. Comparative pathology and pathogenesis of spontaneous and experimentally induced fibropapillomas of green turtles (Chelonia mydas). Vet. Pathol. 1999, 36, 551–564. [Google Scholar] [CrossRef]
- Monezi, T.A.; Mehnert, D.U.; de Moura, E.M.M.; Muller, N.M.G.; Garrafa, P.; Matushima, E.R.; Werneck, M.R.; Borella, M.I. Chelonid herpesvirus 5 in secretions and tumor tissues from green turtles (Chelonia mydas) from Southeastern Brazil: A ten-year study. Vet. Microbiol. 2016, 186, 150–156. [Google Scholar] [CrossRef]
- Page-Karjian, A.; Norton, T.M.; Ritchie, B.; Brown, C.C.; Mancia, C.; Jackwood, M.; Gottdenker, N.L. Quantifying chelonid herpesvirus 5 in symptomatic and asymptomatic rehabilitating green sea turtles. Endanger. Species Res. 2015, 28, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Page-Karjian, A.; Norton, T.M.; Ritchie, B.; Whitfield, J.; Gottdenker, N.L. Potential non-cutaneous sites of chelonid herpesvirus 5 persistence and shedding in green sea turtles (Chelonia mydas). J. Aquat. Anim. Health 2017, 29, 136–142. [Google Scholar] [CrossRef]
- Yetsko, K.; Farrell, J.A.; Blackburn, N.B.; Whitmore, L.; Stammnitz, M.R.; Whilde, J.; Eastman, C.B.; Ramia, D.R.; Thomas, R.; Krstic, A.; et al. Molecular characterization of a marine turtle tumor epizootic, profiling external, internal and post-surgical regrowth tumors. Commun. Biol. 2021, 4, 152. [Google Scholar] [CrossRef]
- Milton, S.L.; Lutz, P.L. Physiological and genetic responses to environmental stress. In The Biology of Sea Turtles Volume II; Lutz, P.L., Musick, J.A., Wyneken, J., Eds.; CRC Press: Boca Raton, FL, USA, 2003; pp. 163–197. [Google Scholar]
- Page-Karjian, A.; Serrano, M.E.; Cartzendafner, J.; Morgan, A.; Ritchie, B.W.; Gregory, C.R.; McNeill, J.B.; Perrault, J.R.; Christiansen, E.F.; Harms, C.A. Molecular assessment of chelonid alphaherpesvirus 5 infection in tumor-free green (Chelonia mydas) and loggerhead (Caretta caretta) sea turtles in North Carolina, USA, 2015–2019. Animals 2020, 10, 1964. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.A.; Balazs, G.H. Blood biochemistry values of green turtles, Chelonia mydas, with and without fibropapillomatosis. Comp. Haematol. Int. 2000, 10, 132–137. [Google Scholar] [CrossRef]
- Cray, C.; Varella, R.; Bossart, G.D.; Lutz, P. Altered in vitro immune responses in green turtles (Chelonia mydas) with fibropapillomatosis. J. Zoo Wildl. Med. 2001, 32, 436–440. [Google Scholar] [PubMed]
- Work, T.M.; Rameyer, R.A.; Balazs, G.H.; Cray, C.; Chang, S.P. Immune status of free-ranging green turtles with fibropapillomatosis from Hawaii. J. Wildl. Dis. 2001, 37, 574–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page-Karjian, A.; Norton, T.M.; Krimer, P.; Groner, M.; Nelson, S.E., Jr.; Gottdenker, N.L. Factors influencing survivorship of rehabilitating green sea turtles (Chelonia mydas) with fibropapillomatosis. J. Zoo Wildl. Med. 2014, 45, 507–519. [Google Scholar] [CrossRef]
- Deus Santos, M.R.d.; Martins, A.S.; Baptistotte, C.; Work, T.M. Health condition of juvenile Chelonia mydas related to fibropapillomatosis in southeast Brazil. Dis. Aquat. Org. 2015, 115, 193–201. [Google Scholar] [CrossRef] [Green Version]
- da Silva, C.C.; Bianchini, A. Blood cholesterol as a biomarker of fibropapillomatosis in green turtles. Mar. Turt. Newsl. 2019, 158, 16–21. [Google Scholar]
- Li, T.-H.; Chang, C.-C. The impact of fibropapillomatosis on clinical characteristics, blood gas, plasma biochemistry, and hematological profiles in juvenile green turtles (Chelonia mydas). Bull. Mar. Sci. 2020, 96, 723–734. [Google Scholar] [CrossRef]
- Davison, A.J. Herpesvirus systematics. Vet. Microbiol. 2010, 143, 52–69. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, E.R.; Gaskin, J.M.; Roelke, M.; Greiner, E.C.; Allen, J. Conjunctivitis, tracheitis, and pneumonia associated with herpesvirus infection in green sea turtles. J. Am. Vet. Med. Assoc. 1986, 189, 1020–1023. [Google Scholar]
- Coberley, S.S.; Herbst, L.H.; Brown, D.R.; Ehrhart, L.M.; Bagley, D.A.; Schaf, S.A.; Moretti, R.H.; Jacobson, E.R.; Klein, P.A. Detection of antibodies to a disease-associated herpesvirus of the green turtle, Chelonia mydas. J. Clin. Microbiol. 2001, 39, 3572–3577. [Google Scholar] [CrossRef] [Green Version]
- Coberley, S.S.; Herbst, L.H.; Ehrhart, L.M.; Bagley, D.A.; Hirama, S.; Jacobson, E.R.; Klein, P.A. Survey of Florida green turtles for exposure to a disease-associated herpesvirus. Dis. Aquat. Org. 2001, 47, 159–167. [Google Scholar] [CrossRef]
- Iverson, R.L.; Bittaker, H.F. Seagrass distribution and abundance in eastern Gulf of Mexico coastal waters. Estuar. Coast. Shelf Sci. 1986, 22, 577–602. [Google Scholar] [CrossRef]
- Hale, J.A.; Frazer, T.K.; Tomasko, D.A.; Hall, M.O. Changes in the distribution of seagrass species along Florida’s Central Gulf Coast: Iverson and Bittaker revisited. Estuaries 2004, 27, 36–43. [Google Scholar] [CrossRef]
- Livingston, R.J. Ontogenetic trophic relationships and stress in a coastal seagrass system in Florida. In Estuarine Perspectives; Kennedy, V., Ed.; Academic Press: New York, NY, USA, 1980; pp. 423–435. [Google Scholar]
- Seavy, J.R.; Pine III, W.E.; Frederick, P.; Sturmer, L.; Berrigan, M. Decadal changes in oyster reefs in the Big Bend of Florida’s Gulf Coast. Ecosphere 2011, 2, 1–14. [Google Scholar] [CrossRef]
- Swain, H.M.; Breininger, D.R.; Busby, D.S.; Clark, K.B.; Cook, S.B.; Day, R.A.; Defreese, D.E.; Gilmore, R.G.; Hart, A.W.; Hinkle, C.R.; et al. Indian River Lagoon biodiversity conference–Introduction. Bull. Mar. Sci. 1995, 57, 1–7. [Google Scholar]
- Indian River Lagoon National Estuary Program (IRLNEP). IRL Comprehensive Conservation and Management Plan Update; IRL National Estuary Program: Palm Bay, FL, USA, 2008; p. 120. Available online: https://www.epa.gov/sites/production/files/2015-09/documents/ccmp_update_2008_final.pdf (accessed on 6 November 2020).
- Sigua, G.C.; Tweedale, W.A. Watershed scale assessment of nitrogen and phosphorus loadings in the Indian River Lagoon basin, Florida. J. Environ. Manag. 2003, 67, 363–372. [Google Scholar] [CrossRef]
- Barile, P.J. Evidence of anthropogenic nitrogen enrichment of the littoral waters of east central Florida. J. Coast. Res. 2004, 20, 1237–1245. [Google Scholar] [CrossRef]
- Barile, P.J. Widespread sewage pollution of the Indian River Lagoon system, Florida (USA) resolved by spatial analyses of macroalgal biogeochemistry. Mar. Pollut. Bull. 2018, 128, 557–574. [Google Scholar] [CrossRef]
- Ehrhart, L.; Redfoot, W.; Mansfield, K.; Gorham, J.; Weege, S.; Provancha, J. Prevalence and Trends in Fibropapillomatosis in Green Turtles on Florida’s Atlantic Coast. In Proceedings of the 2015 International Summit on Fibropapillomatosis: Global Status, Trends, and Population Impacts, Honolulu, HI, USA, 11–14 June 2015; Hargrove, S., Work, T., Brunson, S., Foley, A.M., Balazs, G., Eds.; NOAA Technical Memorandum, NOAA-TM-NMFS-PIFSC-54. U.S. Department of Commerce: Honolulu, HI, USA, 2016; pp. 15–21. [Google Scholar]
- Crigger, D.K.; Graves, G.A.; Fike, D.L. Lake Worth Lagoon conceptual ecological model. Wetlands 2005, 25, 943–954. [Google Scholar]
- Bolter, K. An Ecological Study of Photoautotrophs in Lake Worth. Master’s Thesis, Florida Atlantic University, Boca Raton, FL, USA, 2010. [Google Scholar]
- Eaton, C.; McMichael, E.; Witherington, B.; Foley, A.; Hardy, R.; Meylan, A. In-Water Sea Turtle Monitoring and Research in Florida: Review and Recommendations; NOAA Technical Memorandum NMFS-OPR-38; U.S. Department of Commerce: St. Petersburg, FL, USA, 2008; p. 233.
- Deem, S.L.; Harris, H.S. Health assessments. In Sea Turtle Health and Rehabilitation; Manire, C.A., Norton, T.M., Stacy, B.A., Harms, C.A., Innis, C.J., Eds.; J. Ross Publishing: Plantation, FL, USA, 2017; pp. 945–957. [Google Scholar]
- Page-Karjian, A.; Perrault, J.R. Sea turtle health assessments: Maximizing turtle encounters to better understand health. In Sea Turtle Research and Conservation: Lessons from Working in the Field; Nahill, B., Ed.; Academic Press: San Diego, CA, USA, 2021; pp. 31–44. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Herbst, L.H.; Lemaire, S.; Ene, A.R.; Heslin, D.J.; Ehrhart, L.M.; Bagley, D.A.; Klein, P.A.; Lenz, J. Use of baculovirus-expressed glycoprotein H in an enzyme-linked immunosorbent assay developed to assess exposure to chelonid fibropapillomatosis-associated herpesvirus and its relationship to the prevalence of fibropapillomatosis in sea turtles. Clin. Vaccine Immunol. 2008, 15, 843–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousselet, E.; Levin, M.; Gebhard, E.; Higgins, B.M.; De Guise, S.; Godard-Codding, C.A. Evaluation of immune functions in captive immature loggerhead sea turtles (Caretta caretta). Vet. Immunol. Immunopathol. 2013, 156, 43–53. [Google Scholar] [CrossRef]
- Rousselet, E.; Levin, M.; Gebhard, E.; Higgins, B.M.; De Guise, S.; Godard-Codding, C. Polychlorinated biphenyls modulate both phagocytosis and NK cell activity in vitro in juvenile loggerhead sea turtles (Caretta caretta). J. Toxicol. Environ. Health A 2017, 80, 556–561. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FWC. Marine Turtle Conservation Handbook. 2016. Available online: https://myfwc.com/media/3133/fwc-mtconservationhandbook.pdf (accessed on 10 December 2020).
- Williams, E.H.; Bunkley-Williams, L.; Peters, E.C.; Pinto-Rodriguez, B.; Matos-Morales, R.; Mignucci-Giannoni, A.A.; Hall, K.V.; Rueda-Almonacid, J.V.; Sybesma, J.; de Calventi, I.B.; et al. An epizootic of cutaneous fibropapillomas in green turtles Chelonia mydas of the Caribbean: Part of a panzootic? J. Aquat. Anim. Health 1994, 6, 70–78. [Google Scholar] [CrossRef]
- Ene, A.; Su, M.; Lemaire, S.; Rose, C.; Schaff, S.; Moretti, R.; Lenz, J.; Herbst, L.H. Distribution of chelonid fibropapillomatosis-associated herpesvirus variants in Florida: Molecular genetic evidence for infection of turtles following recruitment to neritic developmental habitats. J. Wildl. Dis. 2005, 41, 489–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holloway-Adkins, K.G. A comparative study of the feeding ecology of Chelonia mydas (green turtle) and the incidental ingestion of Prorocentrum spp. Master’s Thesis, University of Central Florida, Orlando, FL, USA, 2001. [Google Scholar]
- Bresette, M.J.; Herren, R.M.; Singewald, D.A. Comparison of Fibropapilloma Rates of Green Turtles (Chelonia mydas) from Two Different Sites in St. Lucie County, Florida. In Proceedings of the Twenty-First Annual Symposium on Sea Turtle Biology and Conservation, Philadelphia, PA, USA, 24–28 February 2001; NOAA Technical Memorandum, NOAA-NMFS-SEFSC-528. Coyne, M.S., Clark, R.D., Eds.; U.S. Department of Commerce: Miami, FL, USA, 2004; pp. 125–126. [Google Scholar]
- Herren, R.M.; Bagley, D.A.; Bresette, M.J.; Holloway-Adkins, K.G.; Clark, D.; Witherington, B.E. Sea turtle abundance and demographic measurements in a marine protected area in the Florida Keys, USA. Herpetol. Conserv. Biol. 2018, 13, 224–239. [Google Scholar]
- Mattson, R.A. Seagrass ecosystem characteristics and research and management needs in the Florida Big Bend. In Seagrasses: Monitoring, Ecology, Physiology, and Management; Bortone, S.A., Ed.; CRC Press: Boca Raton, FL, USA, 2000; pp. 259–277. [Google Scholar]
- Patrício, A.R.; Velez-Zuazo, X.; Diez, C.E.; Van Dam, R.; Sabat, A.M. Survival probability of immature green turtles in two foraging grounds at Culebra, Puerto Rico. Mar. Ecol. Prog. Ser. 2011, 440, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Greenblatt, R.J.; Work, T.M.; Balazs, G.H.; Sutton, C.A.; Casey, R.N.; Casey, J.W. The Ozobranchus leech is a candidate mechanical vector for the fibropapilloma-associated turtle herpesvirus found latently infecting skin tumors on Hawaiian green turtles (Chelonia mydas). Virology 2004, 321, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Rittenburg, L.T.; Kelley, J.R.; Mansfield, K.L.; Savage, A.E. Marine leech parasitism of sea turtles varies across host species, seasons, and the tumor disease fibropapillomatosis. Dis. Aquat. Org. 2021, 143, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Alfaro-Núñez, A.; Bertelsen, M.F.; Bojesen, A.M.; Rasmussen, I.; Zepeda-Mendoza, L.; Olsen, M.T.; Gilbert, M.T.P. Global distribution of Chelonid fibropapilloma-associated herpesvirus among clinically healthy sea turtles. BMC Evol. Biol. 2014, 14, 206. [Google Scholar] [CrossRef]
- Laegreid, W.W.; Skowronek, A.; Stone-Marschat, M.; Burrage, T. Characterization of virulence variants of African horsesickness virus. Virology 1993, 195, 836–839. [Google Scholar] [CrossRef]
- Herbst, L.; Ene, A.; Su, M.; Sesalle, R.; Lenz, J. Tumor outbreaks in marine turtles are not due to recent herpesvirus mutations. Curr. Biol. 2004, 14, R697–R699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yunis, R.; Jarosinski, K.W.; Schat, K.A. Association between rate of viral genome replication and virulence of Marek’s disease herpesvirus strains. Virology 2004, 328, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, K.; Burgess, G.; Budd, A.M.; Huerlimann, R.; Mashkour, N.; Ariel, E. Molecular evidence for horizontal transmission of chelonid alphaherpesvirus 5 at green turtle (Chelonia mydas) foraging grounds in Queensland, Australia. PLoS ONE 2020, 15, e0227268. [Google Scholar] [CrossRef] [PubMed]
- Work, T.M.; Balazs, G.H.; Rameye, R.A.; Morris, R.A. Retrospective pathology survey of green turtles Chelonia mydas with fibropapillomatosis in the Hawaiian Islands, 1993–2003. Dis. Aquat. Org. 2004, 62, 163–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.; Sánchez-Sarmiento, A.M.; dos Santos, R.G.; Zamana, R.R.; Prioste, F.E.S.; Gattamorta, M.A.; Ochoa, P.F.C.; Grisi-Filho, J.H.H.; Matushima, E.R. Monitoring green sea turtles in Brazilian feeding areas: Relating body condition index to fibropapillomatosis prevalence. J. Mar. Biol. Assoc. UK 2019, 99, 1879–1887. [Google Scholar] [CrossRef]
- Lawrance, M.F.; Mansfield, K.L.; Sutton, E.; Savage, A.E. Molecular evolution of fibropapilloma-associated herpesviruses infecting juvenile green and loggerhead sea turtles. Virology 2018, 521, 190–197. [Google Scholar] [CrossRef]
- James, A. Prevalence of Chelonid Alphaherpesvirus 5 and Associated Disease in Sea Turtles in Grenada, West Indies. Master’s Thesis, St. George’s University, True Blue, Grenada, 2020. [Google Scholar]
- Page-Karjian, A.; Herbst, L.H. Viruses. In Sea Turtle Health and Rehabilitation; Manire, C.A., Norton, T.M., Stacy, B.A., Harms, C.A., Innis, C.J., Eds.; J. Ross Publishing: Plantation, FL, USA, 2017; pp. 751–777. [Google Scholar]
- Rouse, B.T.; Sehrawat, S. Immunity and immunopathology to viruses: What decides the outcome? Nat. Rev. Immunol. 2010, 10, 514–526. [Google Scholar] [CrossRef]
- White, D.W.; Beard, R.S.; Barton, E.S. Immune modulation during latent herpesvirus infection. Immunol. Rev. 2012, 245, 189–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Work, T.M.; Dagenais, J.; Willimann, A.; Balazs, G.; Mansfield, K.; Ackermann, M. Differences in antibody response against chelonid alpha herpesvirus 5 (ChHV5) suggest differences in virus biology in ChHV5-seropositive green turtles from Hawaii and ChHV5-seropositive green turtles from Florida. J. Virol. 2020, 94, e01658-19. [Google Scholar] [PubMed]
- Curry, S.S.; Brown, D.R.; Gaskin, J.M.; Jacobson, E.R.; Ehrhart, L.M.; Blahak, S.; Herbst, L.H.; Klein, P.A. Persistent infectivity of a disease-associated herpesvirus in green turtles after exposure to seawater. J. Wildl. Dis. 2000, 36, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Perrault, J.R.; Page-Karjian, A. Disease survey of loggerhead sea turtles (Caretta caretta) in Florida’s Big Bend. Unpublished data.
- Aguirre, A.A.; Balazs, G.H.; Spraker, T.R.; Gross, T.S. Adrenal and hematological responses to stress in juvenile green turtles (Chelonia mydas) with and without fibropapillomas. Physiol. Zool. 1995, 68, 831–854. [Google Scholar] [CrossRef]
- Work, T.M.; Balazs, G.H.; Wolcott, M.; Morris, R. Bacteraemia in free-ranging Hawaiian green turtles Chelonia mydas with fibropapillomatosis. Dis. Aquat. Org. 2003, 53, 41–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.; Hazarbassanov, N.G.T.d.Q.; Sánchez-Sarmiento, A.M.; Prioste, F.E.S.; Matushima, E.R. Immune response of green sea turtles with and without fibropapillomatosis: Evaluating oxidative burst and phagocytosis via flow cytometry. Chelonian Conserv. Biol. 2016, 15, 273–278. [Google Scholar] [CrossRef]
- Yetsko, K.; Farrell, J.; Stammnitz, M.R.; Whitmore, L.; Whilde, J.; Eastman, C.B.; Ramia, D.R.; Thomas, R.; Krstic, A.; Linse, P.; et al. Mutational, transcriptional and viral shedding dynamics of the marine turtle fibropapillomatosis tumor epizootic. bioRxiv 2021, in press. [Google Scholar]
- Borysenko, M.; Lewis, S. The effect of malnutrition on immunocompetence and whole body resistance to infection in Chelydra serpentina. Dev. Comp. Immunol. 1979, 3, 89–100. [Google Scholar] [CrossRef]
- Acevedo-Whitehouse, K.; Duffus, A.L.J. Effects of environmental change on wildlife health. Philos. Trans. R. Soc. B 2009, 364, 3429–3438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerwenka, A.; Lanier, L.L. Natural killer cells, viruses and cancer. Nat. Rev. Immunol. 2001, 1, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Keller, J.M.; Peden-Adams, M.M.; Aguirre, A.A. Immunotoxicology and implications for reptilian health. In Toxicology of Reptiles; Gardner, S.C., Oberdörster, E., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 199–240. [Google Scholar]
- Ghorai, S.M.; Priyam, M. Reptilia: Cellular immunity in reptiles: Perspective on elements of evolution. In Advances in Comparative Immunology; Cooper, E.L., Ed.; Springer: Cham, Switzerland, 2018; pp. 773–792. [Google Scholar]
- Ishido, S.; Choi, J.-K.; Lee, B.-S.; Wang, C.; DeMaria, M.; Johnson, R.P.; Cohen, G.B.; Jung, J.U. Inhibition of natural killer cell-mediated cytotoxicity by Kaposi’s sarcoma-associated herpesvirus K5 protein. Immunity 2000, 13, 365–374. [Google Scholar] [CrossRef] [Green Version]
- De Pelsmaeker, S.; Romero, N.; Vitale, M.; Favoreel, H.W. Herpesvirus evasion of natural killer cells. J. Virol. 2018, 92, e02105-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, F.J.; De la Fuente, M. The effect of the seasonal cycle on the splenic leukocyte functions in the turtle Mauremys caspica. Physiol. Biochem. Zool. 2001, 74, 660–667. [Google Scholar] [CrossRef]
- Muñoz, F.J.; De la Fuente, M. The immune response of thymic cells from the turtle Mauremys caspica. J. Comp. Physiol. B 2001, 171, 195–200. [Google Scholar]
- Blackburn, N.B.; Leandro, A.C.; Nahvi, N.; Devlin, M.A.; Leandro, M.; Escobedo, I.M.; Peralta, J.M.; George, J.; Stacy, B.A.; deMaar, T.W.; et al. Transcriptomic profiling of fibropapillomatosis in green sea turtles (Chelonia mydas) from south Texas. Front. Immunol. 2021, 12, 630988. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Koriabine, M.; Hartmann-Fritsch, F.; de Jong, P.J.; Lewis, T.D.; Schetle, N.; Work, T.M.; Dagenais, J.; Balazs, G.H.; Leong, J.-A.C. The genome of chelonid herpesvirus 5 harbors atypical genes. PLoS ONE 2012, 7, e46623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, M.J.; Godfrey, D.I.; Trapani, J.A. A fresh look at tumor immunosurveillance and immunotherapy. Nat. Immunol. 2001, 2, 293–299. [Google Scholar] [CrossRef]
- Hamann, M.; Godfrey, M.; Seminoff, J.; Arthur, K.; Barata, P.; Bjorndal, K.; Bolten, A.; Broderick, A.; Campbell, L.; Carreras, C.; et al. Global research priorities for sea turtles: Informing management and conservation in the 21st century. Endanger. Spec. Res. 2010, 11, 245–269. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physical Examination | Big Bend | Jennings Cove | Jupiter Inlet | Total |
|---|---|---|---|---|
| N | 16 | 18 | 11 | 45 |
| Sampling dates | Aug 2020 | Sep 2018–Oct 2020 | Sep 2020 | Sep 2019–Oct 2020 |
| SCL (cm) | 34.4 ± 4.6 A (33.8) (27.9–43.4) | 48.2 ± 10.3 B (49.8) (32.5–70.6) | 35.4 ± 9.1 A (32.1) (27.2–53.0) | 40.2 ± 10.6 (36.9) (27.2–70.6) |
| Body mass (kg) | 5.6 ± 2.7 A (5.2) (2.8–12.3) | 16.8 ± 11.6 B (14.4) (4.5–46.8) | 6.7 ± 6.4 A (3.8) (2.8–19.5) | 10.2 ± 9.4 (6.6) (2.8–46.8) |
| Body condition index | 1.29 ± 0.11 (1.31) (1.10–1.50) | 1.32 ± 0.12 (1.31) (1.07–1.52) | 1.26 ± 0.09 (1.27) (1.10–1.39) | 1.29 ± 0.11 (1.31) (1.07–1.52) |
| Subjective body condition | Good: 44% (7/16) Robust: 56% (9/16) | Good: 72% (13/18) Robust: 28% (5/18) | Good: 91% (10/11) Robust: 9% (1/11) | Good: 67% (30/45) Robust: 33% (15/45) |
| Leech prevalence | 0% (0/16) A | 50% (9/18) B | 9% (1/11) AB | 22% (10/45) |
| Leech cocoon prevalence | 0% (0/16) A | 44% (8/18) B | 9% (1/11) AB | 20% (9/45) |
| Barnacle prevalence | 63% (10/16) | 61% (11/18) | 73% (8/11) | 64% (29/45) |
| Flipper damage | 6% (1/16) | 22% (4/18) | 18% (2/11) | 16% (7/45) |
| Carapace damage | 19% (3/16) | 11% (2/18) | 0% (0/11) | 11% (5/45) |
| FP and diagnostic assays | Big Bend | Jennings Cove | Jupiter Inlet | Total |
| FP prevalence | 69% (11/16) | 56% (10/18) | 36% (4/11) | 56% (25/45) |
| Balazs–Work tumor score | 1.0 ± 0.8 (1) (0–2) | 1.3 ± 1.2 (2) (0–3) | 0.5 ± 0.8 (0) (0–2) | 1.0 ± 1.0 (1) (0–3) |
| Tumor number | 10 ± 15 (3) (0–44) | 21 ± 25 (13) (0–79) | 4 ± 8 (0) (0–26) | 13 ± 20 (2) (0–79) |
| ChHV5 qPCR+ | 6% (1/16) | 0% (0/13) | 0% (0/11) | 3% (1/40) |
| ChHV5 seropositive | 13% (2/15) | 46% (6/16) | 0% (0/9) | 20% (8/40) |
| ChHV6 qPCR+ | 0% (0/16) | 0% (0/13) | 0% (0/11) | 0% (0/40) |
| ChHV6 seropositive | 0% (0/15) | 8% (1/13) | 0% (0/9) | 3% (1/37) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perrault, J.R.; Levin, M.; Mott, C.R.; Bovery, C.M.; Bresette, M.J.; Chabot, R.M.; Gregory, C.R.; Guertin, J.R.; Hirsch, S.E.; Ritchie, B.W.; et al. Insights on Immune Function in Free-Ranging Green Sea Turtles (Chelonia mydas) with and without Fibropapillomatosis. Animals 2021, 11, 861. https://doi.org/10.3390/ani11030861
Perrault JR, Levin M, Mott CR, Bovery CM, Bresette MJ, Chabot RM, Gregory CR, Guertin JR, Hirsch SE, Ritchie BW, et al. Insights on Immune Function in Free-Ranging Green Sea Turtles (Chelonia mydas) with and without Fibropapillomatosis. Animals. 2021; 11(3):861. https://doi.org/10.3390/ani11030861
Chicago/Turabian StylePerrault, Justin R., Milton Levin, Cody R. Mott, Caitlin M. Bovery, Michael J. Bresette, Ryan M. Chabot, Christopher R. Gregory, Jeffrey R. Guertin, Sarah E. Hirsch, Branson W. Ritchie, and et al. 2021. "Insights on Immune Function in Free-Ranging Green Sea Turtles (Chelonia mydas) with and without Fibropapillomatosis" Animals 11, no. 3: 861. https://doi.org/10.3390/ani11030861