
Lunar Phases and Wildlife–Vehicle Collisions: Application of the Lunar Disk Percentage Method


{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
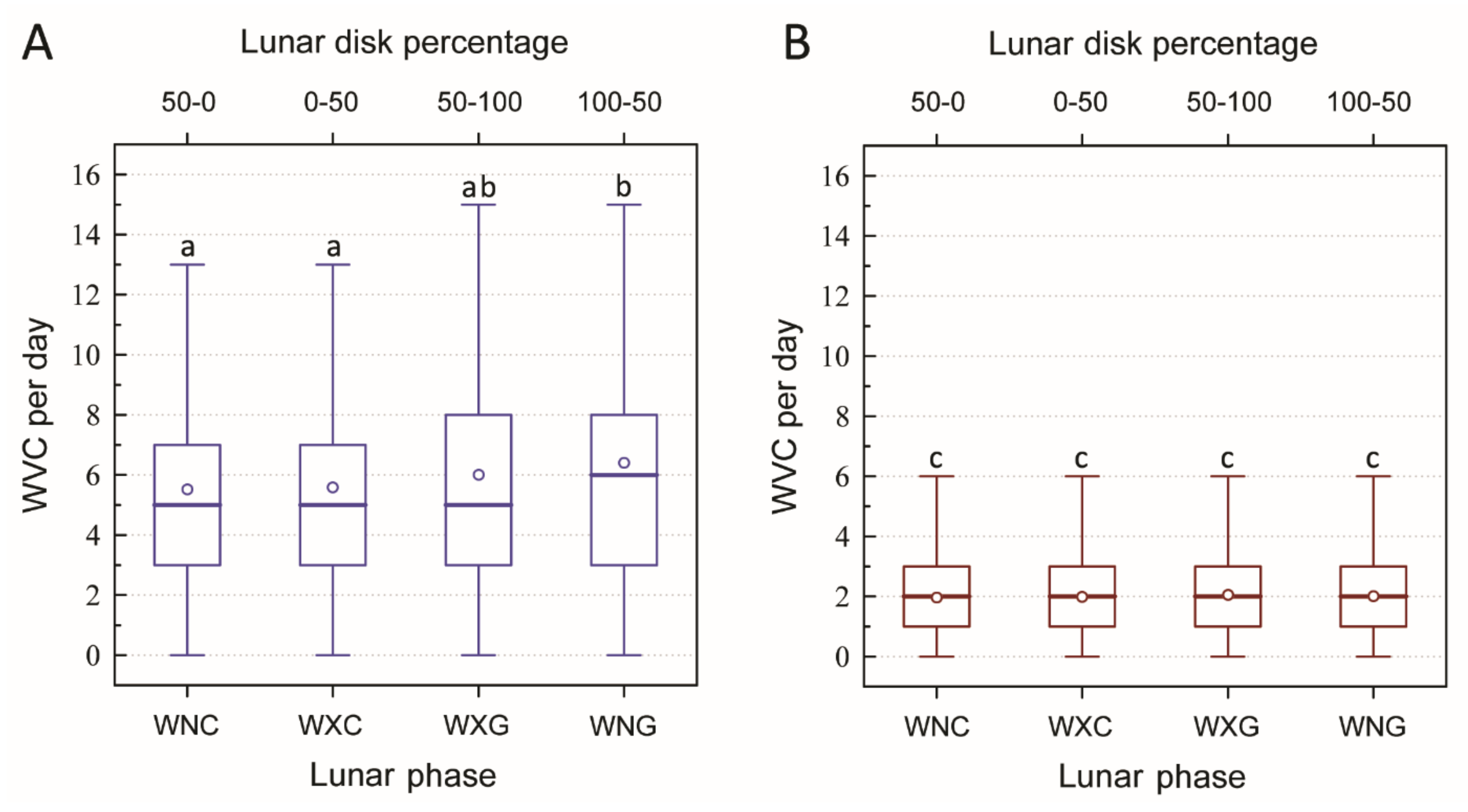
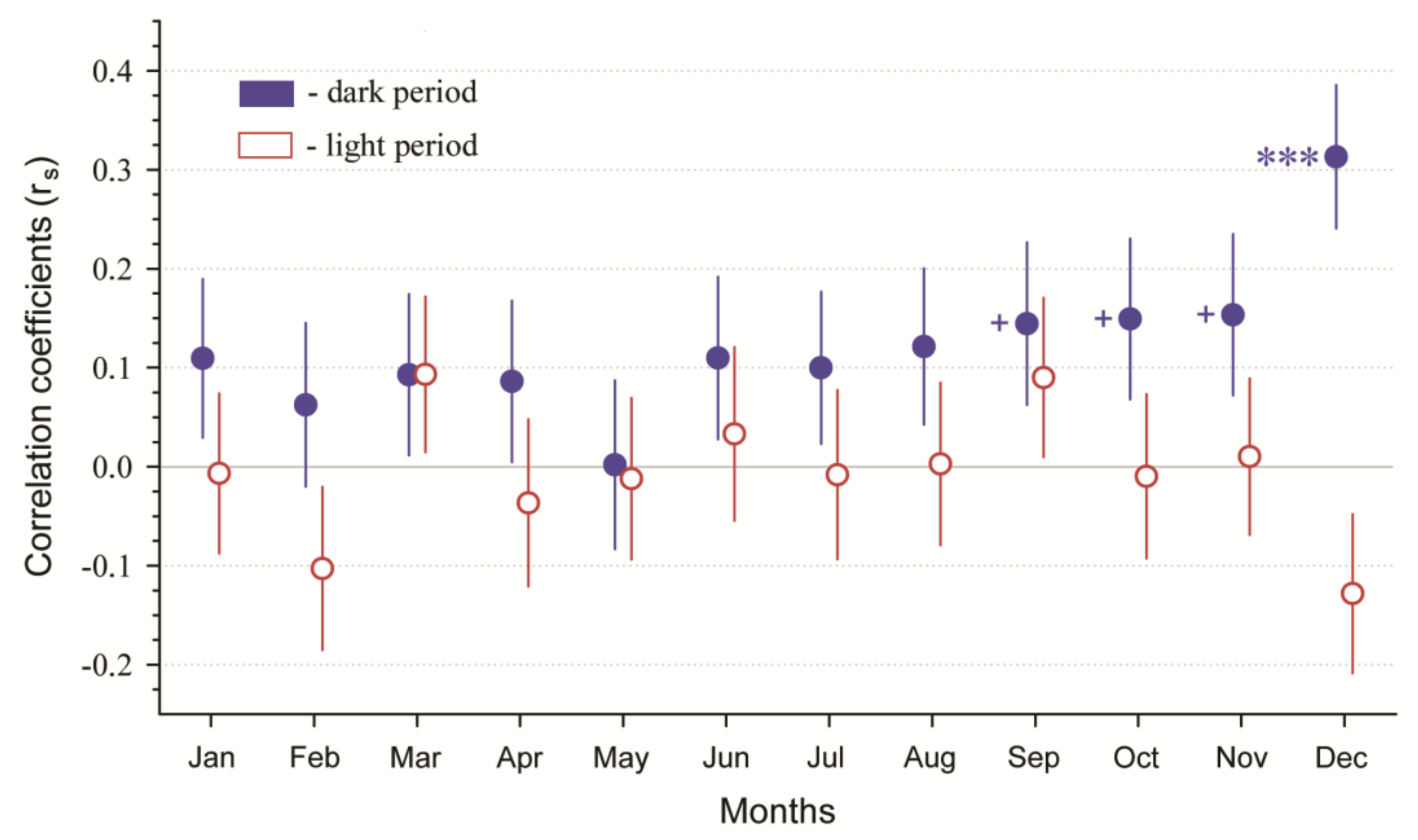
3. Results
4. Discussion
5. Conclusions
- The results of our analysis allow us to accept our first hypothesis that increasing lunar illumination, expressed by increasing the percentage of the visible lunar disk, is correlated with a higher number of WVCs, though trend lines were rather flat and correlation coefficients were low, probably due to the specificity of weather conditions (e.g., high cloudiness) in Lithuania.
- Wildlife–vehicle collisions and LDP were significantly correlated during the dark period (night) during winter, summer, and autumn but not spring. No seasonal influence was observed for the light period (day) between WVCs and LDP. We also observed a general trend of increasing correlation between WVCs and LDP by month, especially from June through December. December was the only month in which the correlation between WVCs and LDP during the dark period was highly significantly different from the other months.
- Our LDP method (ten 10% intervals of the visible lunar disc percentage) allowed for a more refined analysis of the effects of moonlight on WVCs than the broader-scale lunar phase method, which essentially only compares two broad periods of lunar disc percentage periods (50–0–50% and 50–100–50%). Specifically, the LDP method allowed for a more sensitive analysis monthly and seasonal patterns between lunar illumination and WVCs.
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Stewart, S.I.B.; Wilmer, R.B.; Hammer, G.H.; Hawbaker, T.J.; Miller, C.; Radeloff, V.C. Wildland-urban interface maps vary with purpose and context. J. For. 2009, 107, 78–83. [Google Scholar]
- Venter, O.E.; Sanderson, W.; Magrach, A.; Allan, J.R.; Beher, J.; Jones, K.R.; Possingham, H.P.; Laurance, W.F.; Wood, P.; Fekete, B.M.; et al. Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat. Commun. 2016, 7, 12258. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, S.L.; Sivy, K.J.; Pozzanghera, C.B.; DuBour, A.; Overduijn, K.; Smith, M.M.; Zhou, J.; Little, J.M.; Prugh, L.R. Socioeconomic benefits of large carnivore recolonization through reduced wildlife-vehicle collisions. Conserv. Lett. 2016, 10, 431–439. [Google Scholar] [CrossRef]
- Ignatavičius, G.; Ulevičius, A.; Valskys, V.; Trakimas, G.; Galinskaitė, L.; Busher, P.E. Temporal patterns of ungulate-vehicle collisions in a sparsely populated country. Eur. J. Wildl. Res. 2020, 66, 58. [Google Scholar] [CrossRef]
- Kreling, S.E.S.; Gaynor, K.M.; Coon, C.A.C. Roadkill distribution at the wildland-urban interface. J. Wildl. Manag. 2019, 83, 1427–1436. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Alexander, L.E. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef] [Green Version]
- Abra, F.D.; Granziera, B.M.; Huijser, M.P.; Ferraz, K.M.P.M.B.; Haddad, C.M.; Paolino, R.M. Pay or prevent? Human safety, costs to society and legal perspectives on animal-vehicle collisions in São Paulo state, Brazil. PLoS ONE 2019, 14, e0215152. [Google Scholar] [CrossRef] [Green Version]
- Cserkész, T.; Farkas, J. Annual trends in the number of wildlife-vehicle collisions on the main linear transport corridors (highway and railway) of Hungary. North-West. J. Zool. 2015, 11, 41–50. [Google Scholar]
- Cervinka, J.; Riegert, J.; Grill, S.; Salek, M. Large-scale evaluation of carnivore road mortality: The effect of landscape and local scale characteristics. Mammal Res. 2015, 60, 233–243. [Google Scholar]
- Morelle, K.; Lehaire, F.; Lejeune, P. Spatio-temporal patterns of wildlife-vehicle collisions in a region with a high-density road network. Nat. Conserv. 2013, 5, 53–73. [Google Scholar] [CrossRef] [Green Version]
- Rodrıguez-Morales, B.; Dıaz-Varela, E.R.; Marey-Perez, M.F. Spatiotemporal analysis of vehicle collisions involving wild boar and roe deer in NW Spain. Accid. Anal. Prev. 2013, 60, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Steiner, W.; Leisch, F.; Hackländer, K. A review on the temporal pattern of deer–vehicle accidents: Impact of seasonal, diurnal and lunar effects in cervids. Accid. Anal. Prev. 2014, 66, 168–181. [Google Scholar] [CrossRef]
- Šprem, N.; Dudukovic, K.T.; Konjevi, D. Wildlife-vehicle collisions in Croatia—A hazard for humans and animals. Coll. Antropol. 2013, 37, 531–535. [Google Scholar] [PubMed]
- Haikonen, B.A.H.; Summala, H. Deer-vehicle crashes: Extensive peak at 1 hour after sunset. Am. J. Prev. Med. 2001, 21, 209–213. [Google Scholar] [CrossRef]
- Hothorn, T.; Müller, J.; Held, L.; Mysterud, A. Temporal patterns of deer–vehicle collisions consistent with deer activity pattern and density increase but not general accident risk. Accid. Anal. Prev. 2015, 81, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kämmerle, J.L.; Brieger, F.; Kröschel, M.; Hagen, R.; Storch, I.; Suchant, R. Temporal patterns in road crossing behaviour in roe deer (Capreolus capreolus) at sites with wildlife warning reflectors. PLoS ONE 2017, 12, e0184761. [Google Scholar] [CrossRef] [PubMed]
- Redelmeier, D.A.; Shafir, E. The full moon and motorcycle related mortality: Population based double control study. BMJ 2017, 359. [Google Scholar] [CrossRef] [Green Version]
- Ignatavičius, G.; Valskys, V. The influence of time factors on the dynamics of roe deer collisions with vehicles. Landsc. Ecol. Eng. 2018, 14, 221–229. [Google Scholar] [CrossRef]
- Kruuse, M.; Enno, S.E.; Oja, T. Temporal patterns of wild boar-vehicle collisions in Estonia, at the northern limit of its range. Eur. J. Wildl. Res. 2016, 62, 787–791. [Google Scholar] [CrossRef]
- Putzu, N.; Bonetto, D.; Civallero, V.; Fenoglio, S.; Meneguz, P.G.; Preacco, N.; Tizzani, P. Temporal patterns of ungulate-vehicle collisions in a subalpine Italian region. Ital. J. Zool. 2014, 81, 463–470. [Google Scholar] [CrossRef]
- Kotler, B.P.; Brown, J.S.; Hasson, O. Factors Affecting Gerbil Foraging Behavior and Rates of Owl Predation. Ecology 1991, 72, 2249–2260. [Google Scholar] [CrossRef] [Green Version]
- Kronfeld-Schor, N.; Dominoni, D.; Iglesia, H.; Levy, O.; Herzog, E.D.; Dayan, T.; Helfrich-Forster, C. Chronobiology by moonlight. Proc. R. Soc. 2013, B. [Google Scholar] [CrossRef] [Green Version]
- Penteriani, V.; Kuparinen, A.; Delgado, M.; Palomares, F.; López-Bao, J.V.; Fedriani, J.M.; Calzada, J.; Moreno, S.; Villafuerte, R.; Campioni, L.; et al. Responses of a top and a meso predator and their prey to moon phases. Oecologia 2013, 173, 753–766. [Google Scholar] [CrossRef] [Green Version]
- Pratas-Santiago, L.; Goncalves, A.L.S.; Nogueira, A.J.A.; Spironello, W.R. Dodging the moon: The moon effect on activity allocation of prey in the presence of predators. Ethology 2017, 123, 467–474. [Google Scholar] [CrossRef]
- Sullivan, J.D.; Ditchkoff, S.S.; Collier, B.A.; Ruth, C.R.; Raglin, J.B. Movement with the moon: White-tailed deer activity and solunar events. J. Southeast. Assoc. Fish Wildl. Agencies 2016, 3, 225–232. [Google Scholar]
- Prugh, L.R.; Golden, C.D. Does moonlight increase predation risk? Meta-analysis reveals divergent responses of nocturnal mammals to lunar cycles. J. Anim. Ecol. 2014, 83, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Rusak, B. Vertebrate Behavioral Rhythms. In Handbook of Behavioral Neurobiology; Aschoff, J., Ed.; Plenum Press: New York, NY, USA, 1981; Volume 4, pp. 183–213. [Google Scholar]
- Colino-Rabanal, V.J.; Langen, T.A.; Peris, S.; Lizana, M. Ungulate: Vehicle collision rates are associated with the phase of the moon. Biodivers. Conserv. 2018, 27, 681–694. [Google Scholar] [CrossRef]
- Penteriani, V.; Kuparinen, A.; Delgado, M.M.; Lourenço, R.; Campioni, L. Individual status, foraging effort and need for conspicuousness shape behavioural responses of a predator to moon phases. Anim. Behav. 2011, 82, 413–420. [Google Scholar] [CrossRef]
- Vrkljan, J.; Hozjan, D.; Baric, D.; Ugarkovic, D.; Krapinec, K. Temporal patterns of vehicle collisions with roe deer and wild boar in the dinaric area. Croat. J. For. Eng. J. Theory Appl. For. Eng. 2020, 41, 347–357. [Google Scholar] [CrossRef]
- Coulson, G.M. Road-kills of macropods on a section of highway in central Victoria. Wildl. Res. 1982, 9, 21–26. [Google Scholar] [CrossRef]
- Siers, S.R.; Reed, R.N.; Savidge, J.A. To cross or not to cross: Modeling wildlife road crossings as a binary response variable with contextual predictors. Ecosphere 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Baleišis, R.; Bluzma, P.; Balčiauskas, L. Lietuvos Kanopiniai Žvėrys (Ungulates of Lithuania); Asveja Publ.: Vilnius, Lithuania, 1998; pp. 1–218. [Google Scholar]
- Wallach, A.D.; Shanas, U.; Inbar, M. Feeding activity and dietary composition of roe deer at the southern edge of their range. Eur. J. Wildl. Res. 2010, 56, 1–9. [Google Scholar] [CrossRef]
- Pagon, N.; Grignolio, S.; Pipia, A.; Bongi, P.; Bertolucci, C.; Apollonio, M. Seasonal variation of activity patterns in roe deer in a temperate forested area. Chronobiol. Int. 2013, 30, 772–785. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, G.J.; Glasser, A.; Wendt, M.; Williams, G.A.; Osborn, D.A.; Gallagher, G.R.; Warren, R.J.; Miller, K.V.; Pardue, M.T. Visual specialization of an herbivore prey species, the white-tailed deer. Can. J. Zool. 2008, 86, 735–743. [Google Scholar] [CrossRef]
- Sokolov, V.E.; Danilkin, A.A. Sibirskaya Kosulya. Ekologicheskie Aspekty Povedeniya (The Siberian Roe Deer: Ecological Aspects of Behavior); Nauka: Moscow, Russia, 1981. [Google Scholar]
- Abbas, F.; Picot, D.; Merlet, J.; Cargnelutti, B.; Lourtet, B.; Angibault, J.M.; Daufresne, T.; Aulagnier, S.; Verheyden, H.A. Typical browser, the roe deer, may consume substantial quantities of grasses in open landscapes. Eur. J. Wildl. Res. 2012, 59, 69–75. [Google Scholar] [CrossRef]
- Stubbe, M. (Ed.) Buch der Hege. Haarwild. 5. Auflage, auf Basis der 4. bearb. und erw. Auflage; Deutscher Landwirtschaftsverlag Berlin DDR: Berlin, Germany, 1989. [Google Scholar]
- Onozuka, D.; Nishimura, K.; Hagihara, A. Full moon and traffic accident-related emergency ambulance transport: A nationwide case-crossover study. Sci. Total Environ. 2018, 644, 801–805. [Google Scholar] [CrossRef]
- Santos, S.R.; Davis, C.A., Jr.; Smarzaro, R. Analyzing traffic accidents based on the integration of official and crowdsourced data. J. Inf. Data Manag. 2017, 8, 67–82. [Google Scholar]
- Chakraborty, U. Effects of different phases of the lunar month on humans. Biol. Rhythm Res. 2014, 45, 383–396. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ignatavičius, G.; Ulevičius, A.; Valskys, V.; Galinskaitė, L.; Busher, P.E.; Trakimas, G. Lunar Phases and Wildlife–Vehicle Collisions: Application of the Lunar Disk Percentage Method. Animals 2021, 11, 908. https://doi.org/10.3390/ani11030908
Ignatavičius G, Ulevičius A, Valskys V, Galinskaitė L, Busher PE, Trakimas G. Lunar Phases and Wildlife–Vehicle Collisions: Application of the Lunar Disk Percentage Method. Animals. 2021; 11(3):908. https://doi.org/10.3390/ani11030908
Chicago/Turabian StyleIgnatavičius, Gytautas, Alius Ulevičius, Vaidotas Valskys, Lina Galinskaitė, Peter E. Busher, and Giedrius Trakimas. 2021. "Lunar Phases and Wildlife–Vehicle Collisions: Application of the Lunar Disk Percentage Method" Animals 11, no. 3: 908. https://doi.org/10.3390/ani11030908
APA StyleIgnatavičius, G., Ulevičius, A., Valskys, V., Galinskaitė, L., Busher, P. E., & Trakimas, G. (2021). Lunar Phases and Wildlife–Vehicle Collisions: Application of the Lunar Disk Percentage Method. Animals, 11(3), 908. https://doi.org/10.3390/ani11030908