
Use of Corn Silk Meal in Broiler Diet: Effect on Growth Performance, Blood Biochemistry, Immunological Responses, and Growth-Related Gene Expression

 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sampling
2.3. Serum Biochemical Analysis
2.4. Serum Antibody Titers
2.5. Nutrient Digestibility
2.6. RNA Extraction and Real-Time PCR
2.6.1. RNA Isolation and cDNA Synthesis
2.6.2. Q-PCR (Quantitative Real-Time Polymerase Chain Reaction)
2.7. Statistical Analysis
3. Results
3.1. Growth Performance and Organs Weight
3.2. Blood Biochemical Traits
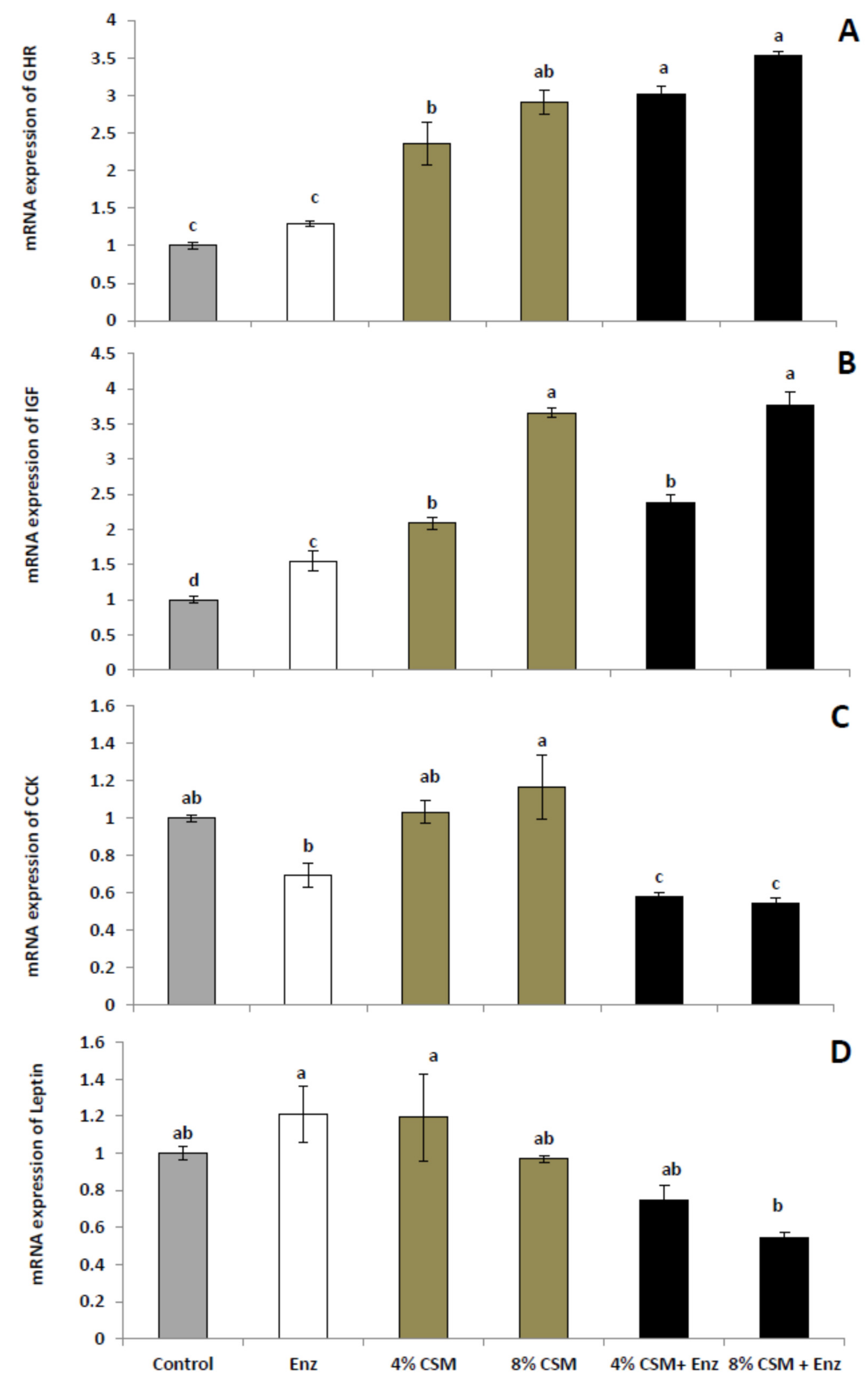
3.3. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sugiharto, S. A review on fungal fermented cassava pulp as a cheap alternative feedstuff in poultry ration. J. World Poult. Res. 2019, 9, 1–6. [Google Scholar] [CrossRef]
- Sugiharto, S.; Yudiarti, T.; Isroli, I.; Widiastuti, E. The potential of tropical agro-industrial by-products as a functional feed for poultry. Iran. J. Appl. Anim. Sci. 2018, 8, 375–385. [Google Scholar]
- Sugiharto, S.; Ranjitkar, S. Recent advances in fermented feeds towards improved broiler chicken performance, gastrointestinal tract microecology and immune responses: A review. Anim. Nutr. 2019, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Haldar, S.; Gan, L.; Tay, S.L.; Ponnalagu, S.; Henry, C.J. Postprandial glycemic and insulinemic effects of the addition of aqueous extracts of dried corn silk, cumin seed powder or tamarind pulp, in two forms, consumed with high glycemic index rice. Foods 2019, 8, 437. [Google Scholar] [CrossRef] [Green Version]
- Rahman, N.A.; Wan Rosli, W.I. Nutritional compositions and antioxidative capacity of the silk obtained from immature and mature corn. J. King Saud Univ. Sci. 2014, 26, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Deng, Z. Protective effects of flavonoids from corn silk on oxidative stress induced by exhaustive exercise in mice. Afr. J. Biotechnol. 2011, 10, 3163–3167. [Google Scholar]
- Ren, S.-H.; Qiao, Q.-Q.; Ding, X.-L. Antioxidative activity of five flavones glycosides from corn silk (Stigma maydis). Czech J. Food Sci. 2013, 2, 148–155. [Google Scholar] [CrossRef]
- El-Ghorab, A.; El-Massry, K.F.; Shibamoto, T. Chemical composition of the volatile extract and antioxidant activities of the volatile and nonvolatile extracts of egyptian corn silk (Zea mays L.). J. Agric. Food Chem. 2007, 55, 9124–9127. [Google Scholar] [CrossRef]
- Ebrahimzadeh, M.A.; Pourmorad, F.; Hafezi, S. Antioxidant activities of iranian corn silk. Turk. J. Biol. 2008, 32, 43–49. [Google Scholar]
- Liu, J.; Wang, C.; Wang, Z.; Zhang, C.; Lu, S.; Liu, J. The antioxidant and free-radical scavenging activities of extract and fractions from corn silk (Zea mays L.) and related flavone glycosides. Food Chem. 2011, 126, 261–269. [Google Scholar] [CrossRef]
- Barter, P.; Gotto, A.M.; LaRosa, J.C.; Maroni, J.; Szarek, M.; Grundy, S.M.; Kastelein, J.J.; Bittner, V.; Fruchart, J.C. HDL cholesterol, very low levels of LDL cholesterol, and cardiovascular events. N. Engl. J. Med. 2007, 357, 1301–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.K.; Purohit, S.S.; Das Prajapati, N. A Handbook on Medicinal Plants. A Complete Source Book; Agrobios: Jodhpur, India, 2009. [Google Scholar]
- Alam, E.A. Evaluation of antioxidant and antibacterial activities of egyptian maydis stigma (Zea mays hairs) rich in some bioactive constituents. J. Am. Sci. 2011, 7, 726–729. [Google Scholar]
- Kaup, S.R.; Arunkumar, N.; Bernhardt, L.K.; Vasavi, R.G.; Shetty, S.S.; Pai, S.R.; Arunkumar, B. Antihyperlipedemic activity of cynodon dactylon extract in high-cholesterol diet fed wistar rats. Genom. Med. Biomark. Health Sci. 2011, 3, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Yin, Y.; Yu, Z.; Liu, J.; Chen, F. Comparison of anti-diabetic effects of polysaccharides from corn silk on normal and hyperglycemia rats. Int. J. Biol. Macromol. 2012, 50, 1133–1137. [Google Scholar] [CrossRef]
- Bedford, M.R. The effect of enzymes on digestion1. J. Appl. Poult. Res. 1996, 5, 370–378. [Google Scholar] [CrossRef]
- Dibner, J.J.; Richards, J.D. The digestive system: Challenges and opportunities. J. Appl. Poult. Res. 2004, 13, 86–93. [Google Scholar] [CrossRef]
- Saleh, A.A.; Gálik, B.; Arpášová, H.; Capcarová, M.; Kalafová, A.; Šimko, M.; Juráček, M.; Rolinec, M.; Bíro, D.; Abudabos, A.M. Synergistic effect of feeding aspergillus awamori and lactic acid bacteria on performance, egg traits, egg yolk cholesterol and fatty acid profile in laying hens. Ital. J. Anim. Sci. 2017, 16, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Omogbenigun, F.O.; Nyachoti, C.M.; Slominski, B.A. Dietary supplementation with multienzyme preparations improves nutrient utilization and growth performance in weaned pigs. J. Anim. Sci. 2004, 82, 1053–1061. [Google Scholar] [CrossRef]
- Anuradha, P.; Barun, R. Effect of supplementation of fiber degrading enzymes on performance of broiler chickens fed diets containing de-oiled rice bran. Asian J. Anim. Vet. Adv. 2015, 10, 179–184. [Google Scholar] [CrossRef] [Green Version]
- NRC. Nutrient Requirements of Poultry, 9th ed.; National Academic Press: Washington, DC, USA, 1994. [Google Scholar]
- Ramadan, M.H.; Mahmoud, A.Z.E.; Zeidan, A.A.B.; Ahmed, A.Z.; Hassaneen, A.S.A. Puerperal Metritis in Crossbreed (Tarentaise X Baladi) Cows: Metabolism-Related Biochemical and Haematological changes. J. Adv. Vet. Res. 2020, 10, 96–104. [Google Scholar]
- OIE. A Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 5th ed.; Part 2 Section 21; OIE: Tokyo, Japan, 2009. [Google Scholar]
- AOAC. International Official Methods of Analysis, 18th ed.; Current Through Revision 2 (Online); AOAC International: Gaithersburg, MD, USA, 1994. [Google Scholar]
- Saleh, A.A. Effect of Low-Protein in Iso-Energetic Diets on Performance, Carcass Characteristics, Digestibilities and Plasma Lipids of Broiler Chickens. Egypt. Poult. Sci. J. 2016, 36, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Prakatur, I.; Miskulin, M.; Pavic, M.; Marjanovic, K.; Blazicevic, V.; Miskulin, I.; Domacinovic, M. Intestinal Morphology in Broiler Chickens Supplemented with Propolis and Bee Pollen. Animals 2019, 9, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Tang, L.; Lu, L.; Zhang, L.; Xi, L.; Liu, H.-C.; Odle, J.; Luo, X. Differential expression of heat shock transcription factors and heat shock proteins after acute and chronic heat stress in laying chickens (Gallus gallus). PLoS ONE 2014, 9, e102204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamel, E.R.; Manaa, E.; Farid, A.S. The effects of dietary date pit on the productive and economic efficiency of Japanese quail. Alex. J. Vet. Sci. 2016, 51, 211–221. [Google Scholar] [CrossRef]
- Ashwell, C.M.; Czerwinski, S.M.; Brocht, D.M.; McMurtry, J.P. Hormonal regulation of leptin expression in broiler chickens. Am. J. Physiol. 1999, 276, R226–R232. [Google Scholar] [CrossRef]
- El-Naggar, K.; El-Kassas, S.; Abdo, S.E.; Kirrella, A.A.K.; Al wakeel, R.A. Role of gamma-aminobutyric acid in regulating feed intake in commercial broilers reared under normal and heat stress conditions. J. Therm. Biol. 2019, 84, 164–175. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Nessa, F.; Ismail, Z.; Mohamed, N. Antimicrobial activities of extracts and flavonoid glycosides of corn silk (Zea mays L.). Int. J. Biotechnol. Wellness Ind. 2012, 1, 115–121. [Google Scholar] [CrossRef]
- Kaur, G.; Alam, M.S.; Jabbar, Z.; Javed, K.; Athar, M. Evaluation of antioxidant activity of cassia siamea flowers. J. Ethnopharmacol. 2006, 108, 340–348. [Google Scholar] [CrossRef]
- Arafa, S.A.; Neven, M.M.; Nematalla, K.M. Effect of corn silk on characteristics and keeping quality of cake. J. Biol. Chem. Environ. Sci. 2012, 7, 371–400. [Google Scholar]
- Horvatovic, M.P.; Glamocic, D.; Zikic, D.; Hadnadjev, T.D. Performance and some intestinal functions of broilers fed diets with different inclusion levels of sunflower meal and supplemented or not with enzymes. Braz. J. Poult. Sci. 2015, 17, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Latif, M.A.; El-Far, A.H.; Elbestawy, A.R.; Ghanem, R.; Mousa, S.A.; Abd El-Hamid, H.S. Exogenous dietary lysozyme improves the growth performance and gut microbiota in broiler chickens targeting the antioxidant and non-specific immunity mrna expression. PLoS ONE 2017, 12, e0185153. [Google Scholar] [CrossRef]
- Slominski, B.A.; Gdala, J.; Boros, D.; Campbell, L.D.; Guenter, W.; Jones, O. Variability in chemical and nutritive composition of canadian wheat and the potential for its minimization by enzyme use. In Proceedings of the XXI World Poultry Congress, Montreal, QC, Canada, 20–24 August 2000. [Google Scholar]
- Ravn, J.L.; Glitsø, V.; Pettersson, D.; Ducatelle, R.; Van Immerseel, F.; Pedersen, N.R. Combined endo-β-1,4-xylanase and α-l-arabinofuranosidase increases butyrate concentration during broiler cecal fermentation of maize glucurono-arabinoxylan. Anim. Feed Sci. Technol. 2018, 236, 159–169. [Google Scholar] [CrossRef]
- Saleh, A.A.; Kirrella, A.A.; Abdo, S.E.; Mousa, M.M.; Badwi, N.A.; Ebeid, T.A.; Nada, A.L.; Mohamed, M.A. Effects of dietary xylanase and arabinofuranosidase combination on the growth performance, lipid peroxidation, blood constituents, and immune response of broilers fed low-energy diets. Animals 2019, 9, 467. [Google Scholar] [CrossRef] [Green Version]
- Saleh, A.A.; Hayashi, K.; Ijiri, D.; Ohtsuka, A. Effect of feeding aspergillus, awamori and canola seed on the growth performance and muscle fatty acid profile in broiler chicken. Anim. Sci. J. Nihon Chikusan Gakkaiho 2015, 86, 305–311. [Google Scholar]
- Teo, A.Y.; Tan, H.M. Evaluation of the performance and intestinal gut microflora of broilers fed on corn-soy diets supplemented with Bacillus subtilis PB6 (CloSTAT). J. Appl. Poult. Res. 2007, 16, 296–303. [Google Scholar] [CrossRef]
- Liu, J.; Yang, F.; Zhang, Y.; Li, J. Studies on the cell-immunosuppressive mechanism of oridonin from Isodon serra. Int. Immunopharmacol. 2007, 7, 945–954. [Google Scholar] [CrossRef]
- Zhang, Y.; Sui, D.Y.; Zhou, J.S.; Zhou, H.L. Microwave-assisted extraction and antihyperlipidemic effect of total flavonoids from corn silk. Afr. J. Biotechnol. 2011, 10, 14583–14586. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wu, L.; Ma, Z.; Cheng, J.; Liu, J. Anti-diabetic, anti-oxidant and anti-hyperlipidemic activities of flavonoids from corn silk on Stz-induced diabetic mice. Molecules 2016, 21, 7. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.H.; Kim, S.R.; Kang, H.J.; Kim, M.H.; Ha, A.W.; Kim, W.K. Corn silk extract improves cholesterol metabolism in C57BL/6J mouse fed high-fat diets. Nutr. Res. Pract. 2016, 10, 501–506. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.-H.; Tang, W.; Jin, M.-L.; Li, J.-E.; Xie, M.-Y. Recent advances in bioactive polysaccharides from Lycium barbarum L., Zizyphus jujuba mill, Plantago spp., and Morus spp.: Structures and functionalities. Food Hydrocoll. 2016, 60, 148–160. [Google Scholar] [CrossRef]
- Catap, E.S.; Jimenez, M.R.R.; Tumbali, M.P.B. Immunostimulatory and anti-oxidative properties of corn silk from Zea mays L. in Nile tilapia, Oreochromis niloticus. Int. J. Fish. Aquac. 2015, 7, 30–36. [Google Scholar]
- Sacheck, J.M.; Ohtsuka, A.; McLary, S.C.; Goldberg, A.L. IGF-I stimulates muscle growth by suppressing protein breakdown and expression of atrophy-related ubiquitin ligases, atrogin-1 and MuRF1. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E591–E601. [Google Scholar] [CrossRef] [Green Version]
- Scanes, C.G. Perspectives on the endocrinology of poultry growth and metabolism. Gen. Comp. Endocrinol. 2009, 163, 24–32. [Google Scholar] [CrossRef]
- Tomas, F.M.; Pym, R.A.; McMurtry, J.P.; Francis, G.L. Insulin-like growth factor (IGF)-I but not IGF-II promotes lean growth and feed efficiency in broiler chickens. Gen. Comp. Endocrinol. 1998, 110, 262–275. [Google Scholar] [CrossRef]
- Kita, K.; Nagao, K.; Okumura, J. Nutritional and tissue specificity of IGF-I and IGFBP-2 gene expression in growing chickens—A review. Asian Australas. J. Anim. Sci. 2005, 18, 747–754. [Google Scholar] [CrossRef]
- Arora, S.; Anubhuti. Role of neuropeptides in appetite regulation and obesity—A review. Neuropeptides 2006, 40, 375–401. [Google Scholar] [CrossRef]
- Guo, S.; Liu, D.; Zhao, X.; Li, C.; Guo, Y. Xylanase supplementation of a wheat-based diet improved nutrient digestion and mrna expression of intestinal nutrient transporters in broiler chickens infected with clostridium perfringens. Poult. Sci. 2014, 93, 94–103. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Manafi, M.; Nazarizadeh, H. Effects of xylanase supplementation and citric acid on performance, ileal nutrients digestibility, and gene expression of intestinal nutrient transporters in broilers challenged with clostridium perfringens. J. Poult. Sci. 2017, 54, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Moneim, E.A.; Shehata, A.M.; Alzahrani, S.O.; Shafi, M.E.; Mesalam, N.M.; Taha, A.E.; Swelum, A.A.; Arif, M.; Fayyaz, M.; Abd El-Hack, M.E. The role of polyphenols in poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1851–1866. [Google Scholar] [CrossRef]


{kind=link}
| Control | Enzyme | 4% CSM | 8% CSM | 4% CSM + Enz | 8% CSM + Enz | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ingredients (g/kg) | Grower | Finisher | Grower | Finisher | Grower | Finisher | Grower | Finisher | Grower | Finisher | Grower | Finisher |
| Yellow corn | 577 | 637 | 576.5 | 636.5 | 550 | 609 | 508 | 569 | 549.5 | 608.5 | 507.5 | 568.5 |
| Soybean meal (46%) | 328 | 260 | 328 | 260 | 322 | 256 | 320 | 248 | 322 | 256 | 320 | 248 |
| Gluten (62%) | 15 | 25 | 15 | 25 | 18 | 26 | 17 | 30 | 18 | 26 | 17 | 30 |
| Soybean oil | 30 | 29 | 30 | 29 | 30 | 30 | 35 | 34 | 30 | 30 | 35 | 34 |
| Dicalcium phosphate | 13 | 15 | 13 | 15 | 13 | 15 | 13 | 15 | 13 | 15 | 13 | 15 |
| DL-Methionine | 3.2 | 2.6 | 3.2 | 2.6 | 3.2 | 2.6 | 3.2 | 2.6 | 3.2 | 2.6 | 3.2 | 2.6 |
| L-Lysine | 2 | 3 | 2 | 3 | 2 | 3 | 2 | 3 | 2 | 3 | 2 | 3 |
| Threonine | 0.8 | 0.9 | 0.8 | 0.9 | 0.8 | 0.9 | 0.8 | 0.9 | 0.8 | 0.9 | 0.8 | 0.9 |
| CaCo3 | 13 | 9 | 13 | 9 | 13 | 9 | 13 | 9 | 13 | 9 | 13 | 9 |
| NaCl | 3.5 | 3.5 | 3.5 | 3.5 | 3.5 | 3.5 | 3.5 | 3.5 | 3.5 | 3.5 | 3.5 | 3.5 |
| Premix * | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| NaHCO3 | 1.5 | 2 | 1.5 | 2 | 1.5 | 2 | 1.5 | 2 | 1.5 | 2 | 1.5 | 2 |
| CSM ** | 40 | 40 | 80 | 80 | 40 | 40 | 80 | 80 | ||||
| NSP enzyme | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | ||||||
| Calculated analysis (%) | ||||||||||||
| Crude Protein | 21.04 | 19.06 | 21.04 | 19.06 | 21.06 | 19.05 | 21.05 | 19.05 | 21.06 | 19.04 | 21.04 | 19.04 |
| ME (Kcal/kg) | 3044 | 3116 | 3043 | 3115 | 3042 | 3114 | 3045 | 3115 | 3044 | 3113 | 3043 | 3115 |
| Calcium | 0.892 | 0.766 | 0.892 | 0.766 | 0.901 | 0.775 | 0.902 | 0.765 | 0.901 | 0.765 | 0.902 | 0.765 |
| Av. P | 0.493 | 0.491 | 0.493 | 0.491 | 0.493 | 0.491 | 0.496 | 0.493 | 0.492 | 0.491 | 0.496 | 0.492 |
| Crude Fiber | 3.304 | 3.081 | 3.303 | 3.080 | 3.484 | 3.266 | 3.754 | 3.518 | 3.483 | 3.265 | 3.753 | 3.516 |
| Na | 0.204 | 0.216 | 0.204 | 0.216 | 0.207 | 0.219 | 0.201 | 0.213 | 0.207 | 0.219 | 0.201 | 0.213 |
| Cl | 0.251 | 0.251 | 0.251 | 0.251 | 0.254 | 0.253 | 0.257 | 0.256 | 0.254 | 0.253 | 0.257 | 0.256 |
| Digestible Lysine | 1.306 | 1.199 | 1.306 | 1.198 | 1.303 | 1.196 | 1.308 | 1.191 | 1.304 | 1.196 | 1.307 | 1.191 |
| Digestible Methionine | 0.676 | 0.601 | 0.676 | 0.600 | 0.669 | 0.592 | 0.658 | 0.585 | 0.669 | 0.591 | 0.657 | 0.585 |
| Digestible Threonine | 0.853 | 0.758 | 0.852 | 0.758 | 0.844 | 0.751 | 0.842 | 0.745 | 0.844 | 0.748 | 0.842 | 0.745 |
| Chemical analysis (%) | ||||||||||||
| Crude Protein | 21.10 | 19.02 | 21.11 | 19.02 | 21.09 | 19.04 | 21.05 | 19.03 | 21.08 | 19.04 | 21.07 | 19.05 |
| Crude Fiber | 3.297 | 3.051 | 3.298 | 3.050 | 3.348 | 3.195 | 3.632 | 3.498 | 3.351 | 3.212 | 3.650 | 3.489 |
| Ether extract | 4.461 | 5.65 | 4.461 | 5.652 | 4.453 | 5.643 | 4.455 | 5.652 | 4.460 | 5.621 | 4.462 | 5.648 |
| Dry matter | 95.9% |
| Crude protein | 12.51% |
| Crude fiber | 9.5% |
| Metabolizable energy (ME) | 2550 Kcal/Kg |
| Calcium | 0.31% |
| Total phosphorus | 0.32% |
| Available phosphorus | 0.24% |
| Digestible lysine | 0.62% |
| Digestible methionine | 0.23% |
| Digestible threonine | 0.36% |
| Gene | Accession No. | Primer | Ref. | Annealing |
|---|---|---|---|---|
| β-actin | NM_205518.1 | F: ACCTGAGCGCAAGTACTCTGTCT R: CATCGTACTCCTGCTTGCTGAT | [25] | 60 |
| Growth Hormone Receptor (GHR) | NM_001001293.1 | F: AACACAGATACCCAACAGCC R: AGAAGTCAGTGTTTGTCAGGG | [27] | 60 |
| Insulin Like Growth Factor (IGF) | NM_001004384.2 | F: CACCTAAATCTGCACGCT R: CTTGTGGATGGCATGATCT | [27] | 60 |
| Leptin | AF082500 | F: TCCGCCAAGCAGAGGGGT R: CCAGGACGCCATCCAGGCTCTCTGGC | [28] | 58 |
| Cholecystokinin (CCK) | NM_001001741.1 | F: CAGCAGAGCCTGACAGAACC R: AGAGAACCTCCCAGTGGAACC | [29] | 58 |
| Treatments | ANOVA | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | Enz. | 4% CSM | 8% CSM | 4% CSM + Enz. | 8% CSM + Enz | Enz | CSM | Enz × CSM | |
| Initial body weight, g at 7 d | 152.3 ± 0.1 | 152.3 ± 0.03 | 152.1 ± 0.16 | 152.1 ± 0.01 | 152.3 ± 0.02 | 152.3 ± 0.02 | NS | NS | NS |
| Feed intake, g 0–7 d | 116.2 ± 0.41 | 116.4 ± 0.34 | 116.2 ± 0.21 | 116.4 ± 0.32 | 115.8 ± 0.32 | 115.7 ± 0.36 | NS | NS | NS |
| FCR, 7 d | 0.763 ± 0.007 | 0.764 ± 0.002 | 0.764 ± 0.004 | 0.765 ± 0.003 | 0.760 ± 0.004 | 0.760 ± 0.005 | NS | NS | NS |
| Body weight, g at 14 d | 456 ± 4 | 462 ± 5 | 455 ± 5 | 460 ± 6 | 468 ± 8 | 461 ± 7 | NS | NS | NS |
| Body weight gain, g (7–14) d | 304 ± 6 | 310 ± 5 | 303 ± 5 | 308 ± 6 | 316 ± 7 | 309 ± 7 | NS | NS | NS |
| Feed intake, g 7–14 d | 462 ± 5 | 466 ± 3 | 461 ± 7 | 469 ± 4 | 471 ± 7 | 468 ± 5 | NS | NS | NS |
| FCR, 14d | 1.519 ± 0.006 ab | 1.503 ± 0.008 b | 1.522 ± 0.007 a | 1.523 ± 0.009 a | 1.491 ± 0.008 b | 1.515 ± 0.006 ab | * | * | NS |
| Body weight, g at 21 d | 932 ± 12 b | 954 ± 15 a | 930 ± 17 b | 941 ± 16 ab | 966 ± 15 a | 944 ± 14 ab | * | * | NS |
| Body weight gain, g (14–21) d | 476 ± 8 b | 492 ± 8 a | 475 ± 7 b | 481 ± 8 ab | 498 ± 9 a | 483 ± 6 ab | * | * | NS |
| Feed intake, g 14-21 d | 716 ± 23 | 704 ± 21 | 712 ± 19 | 710 ± 18 | 701 ± 21 | 706 ± 22 | NS | NS | NS |
| FCR, 21d | 1.504 ± 0.007 a | 1.431 ± 0.006 bc | 1.499 ± 0.007 ab | 1.476 ± 0.009 ab | 1.408 ± 0.004 c | 1.462 ± 0.008 ab | * | NS | * |
| Body weight, g at 28 d | 1502 ± 25 b | 1553 ± 29 a | 1506 ± 37 b | 1523 ± 16 ab | 1654 ± 42 a | 1538 ± 36 ab | * | * | NS |
| Body weight gain, g (21–28) d | 570 ± 12 c | 599 ± 14 b | 576 ± 13 c | 582 ± 15 bc | 688 ± 17 a | 594 ± 16 b | * | * | * |
| Feed intake, g 21–28d | 1010 ± 37 | 998 ± 39 | 1003 ± 28 | 1005 ± 32 | 1009 ± 34 | 1008 ± 38 | NS | NS | NS |
| FCR, 28d | 1.772 ± 0.01 a | 1.650 ± 0.05 bc | 1.741 ± 0.07 ab | 1.727 ± 0.05 ab | 1.467 ± 0.08 c | 1.697 ± 0.07 ab | * | NS | * |
| Body weight, g at 35 d | 1802 ± 51 b | 1854 ± 28 ab | 1796 ± 54 b | 1832 ± 28 ab | 1942 ± 23 a | 1832 ± 32 ab | NS | NS | * |
| Body weight gain, g (28–35) d | 300 ± 12 ab | 301 ± 17 a | 290 ± 13 b | 309 ± 19 a | 280 ± 12 b | 294 ± 17 ab | NS | * | * |
| Feed intake, g (28–35) d | 1067 ± 46 a | 1015 ± 38 b | 1055 ± 43 a | 1062 ± 30 a | 1045 ± 34 ab | 1020 ± 31 b | NS | NS | NS |
| Body weight gain, g (7–35) d | 1650 ± 51.2 b | 1702 ± 28.1 ab | 1643 ± 53.8 b | 1680 ± 28.1 ab | 1790 ± 22.6 a | 1680 ± 31.5 ab | NS | NS | * |
| Feed intake, g (7–35) d | 3371 ± 46 | 3335 ± 38 | 3383 ± 43 | 3398 ± 30 | 3378 ± 34 | 3354 ± 31 | NS | NS | NS |
| FCR, 7–35 | 2.043 ± 0.21 a | 1.959 ± 0.26 ab | 2.059 ± 0.23 a | 2.023 ± 0.31 a | 1.887 ± 0.18 b | 1.997 ± 0.20 ab | NS | NS | * |
| Crude protein retention% | 67.1 ± 0.37 c | 70.67 ± 0.41 b | 67.57 ± 0.23 c | 70.37 ± 0.37 b | 72.63 ± 0.29 a | 72.3 ± 0.26 a | * | * | ** |
| Crude fiber digestibility% | 42.97 ± 0.203 b | 48.13 ± 0.745 a | 43.3 ± 0.208 b | 42.47 ± 0.59 b | 47.67 ± 1.38 a | 45.47 ± 1.41 ab | * | * | * |
| Treatments | ANOVA | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | Enz | 4% CSM | 8% CSM | 4% CSM + Enz | 8% CSM + Enz | Enz | CSM | Enz × CSM | |
| Carcass, g/100 g BW | 59.87 ± 2.4 b | 66.61 ± 2.01 a | 65.8 ± 1.08 a | 66.92 ± 1.13 a | 65.71 ± 2.32 a | 64.85 ± 1.59 ab | * | * | * |
| Breast muscle, g/100 g BW | 21.26 ± 1.21 cd | 20.88 ± 0.63 d | 21.65 ± 0.36 bcd | 25.63 ± 1.08 a | 24.46 ± 1.18 ab | 23.99 ± 0.86 abc | NS | * | ** |
| Thigh muscle, g/100 g BW | 12.59 ± 0.34 b | 13.97 ± 0.66 ab | 14.91 ± 0.87 ab | 15.02 ± 1.21 ab | 15.63 ± 1.48 ab | 17.18 ± 1.41 a | NS | * | ** |
| Bursa, g/100 g BW | 0.093 ± 0.017 b | 0.13 ± 0.015 ab | 0.137 ± 0.016 ab | 0.147 ± 0.019 a | 0.146 ± 0.011 a | 0.141 ± 0.018 ab | NS | * | * |
| Thymus, g/100 g BW | 0.147 ± 0.014 c | 0.201 ± 0.019 bc | 0.244 ± 0.019 abc | 0.314 ± 0.033 a | 0.29 ± 0.047 ab | 0.284 ± 0.054 ab | NS | * | ** |
| Spleen, g/100 g BW | 0.142 ± 0.016 a | 0.086 ± 0.018 b | 0.106 ± 0.008 ab | 0.122 ± 0.016 ab | 0.127 ± 0.006 a | 0.129 ± 0.005 a | * | NS | * |
| Liver, g/100 g BW | 1.987 ± 0.16 | 1.854 ± 0.10 | 2.118 ± 0.15 | 1.947 ± 0.23 | 1.959 ± 0.15 | 1.896 ± 0.04 | NS | NS | NS |
| Abdominal fat, g/100 g BW | 1.11 ± 0.11 | 0.688 ± 0.14 | 0.622 ± 0.07 | 0.711 ± 0.07 | 0.652 ± 0.05 | 0.699 ± 0.04 | NS | NS | NS |
| Treatments | ANOVA | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | Enz | 4% CSM | 8% CSM | 4% CSM + Enz | 8% CSM + Enz | Enz | CSM | Enz × CSM | |
| Total protein, mg/dL | 3.35 ± 0.45 | 3.45 ± 0.15 | 3.575 ± 0.14 | 3.65 ± 0.58 | 3.9 ± 0.42 | 3.8 ± 0.18 | NS | NS | NS |
| Albumin, mg/dL | 2.175 ± 0.048 | 2.225 ± 0.18 | 2.175 ± 0.149 | 2.213 ± 0.131 | 2.1 ± 0.158 | 2.525 ± 0.111 | NS | NS | NS |
| Total cholesterol, mg/dL | 140.0 ± 6.4 ab | 149.3 ± 5.9 a | 130.0 ± 8.5 bc | 125.3 ± 5.5 bc | 119.8 ± 2.4 c | 123.5 ± 2.2 bc | NS | * | * |
| HDL, mg/dL | 30.8 ± 3.3 b | 41.84 ± 1.9 a | 37.86 ± 2.61 ab | 37.51 ± 1.46 ab | 39.92 ± 3.32 a | 42.75 ± 3.4 a | * | * | * |
| LDL, mg/dL | 90.7 ± 3.5 | 86.16 ± 7.2 | 88.44 ± 7.4 | 85.05 ± 5.1 | 73.84 ± 7.8 | 73.85 ± 5.8 | NS | NS | NS |
| ALT, (U/L) | 23.75 ± 6.04 | 24.0 ± 3.24 | 20.5 ± 2.39 | 23.0 ± 3.34 | 21.25 ± 3.30 | 22.5 ± 5.86 | NS | NS | NS |
| AST, (U/L) | 242.3 ± 16.3 | 234 ± 29.2 | 244.3 ± 15.9 | 245.5 ± 14.7 | 229.8 ± 8.5 | 226 ± 12.8 | NS | NS | NS |
| Glucose, mg/dL | 170.5 ± 16.9 a | 144.5 ± 21.56 ab | 174 ± 7.9 a | 159.8 ± 9.3 ab | 121.8 ± 4.7 b | 133.5 ± 7.2 ab | NS | NS | * |
| T3, mg/dL | 0.835 ± 0.198 b | 1.495 ± 0.154 a | 1.358 ± 0.206 a | 1.45 ± 0.048 a | 1.223 ± 0.186 ab | 1.253 ± 0.104 ab | * | * | NS |
| Titers for ND | 1.5 ± 0.29 b | 2.25 ± 0.25 ab | 3.0 ± 0.41 a | 3.25 ± 0.47 a | 2.75 ± 0.62 ab | 3.5 ± 0.29 a | * | * | * |
| Titers for H9N1 | 1.5 ± 0.28 | 1.5 ± 0.28 | 1.75 ± 0.25 | 2.0 ± 0.41 | 2.5 ± 0.29 | 2.25 ± 0.47 | NS | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirrella, A.A.; Abdo, S.E.; El-Naggar, K.; Soliman, M.M.; Aboelenin, S.M.; Dawood, M.A.O.; Saleh, A.A. Use of Corn Silk Meal in Broiler Diet: Effect on Growth Performance, Blood Biochemistry, Immunological Responses, and Growth-Related Gene Expression. Animals 2021, 11, 1170. https://doi.org/10.3390/ani11041170
Kirrella AA, Abdo SE, El-Naggar K, Soliman MM, Aboelenin SM, Dawood MAO, Saleh AA. Use of Corn Silk Meal in Broiler Diet: Effect on Growth Performance, Blood Biochemistry, Immunological Responses, and Growth-Related Gene Expression. Animals. 2021; 11(4):1170. https://doi.org/10.3390/ani11041170
Chicago/Turabian StyleKirrella, Abeer A., Safaa E. Abdo, Karima El-Naggar, Mohamed Mohamed Soliman, Salama Mostafa Aboelenin, Mahmoud A. O. Dawood, and Ahmed A. Saleh. 2021. "Use of Corn Silk Meal in Broiler Diet: Effect on Growth Performance, Blood Biochemistry, Immunological Responses, and Growth-Related Gene Expression" Animals 11, no. 4: 1170. https://doi.org/10.3390/ani11041170
APA StyleKirrella, A. A., Abdo, S. E., El-Naggar, K., Soliman, M. M., Aboelenin, S. M., Dawood, M. A. O., & Saleh, A. A. (2021). Use of Corn Silk Meal in Broiler Diet: Effect on Growth Performance, Blood Biochemistry, Immunological Responses, and Growth-Related Gene Expression. Animals, 11(4), 1170. https://doi.org/10.3390/ani11041170