
Epigenetic Changes in Equine Embryos after Short-Term Storage at Different Temperatures

 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Reproductive Management
2.2. Experimental Design
2.3. Collection and Evaluation of Embryos
2.4. Holding Medium Temperature and pH
2.5. Lipid Peroxidation
2.6. Quantitative Real-Time PCR
2.7. Gene-Specific DNA Methylation Analysis
2.8. Global Methylation-Immunofluorescence Staining for 5mC and 5hmC
2.9. Statistical Analysis
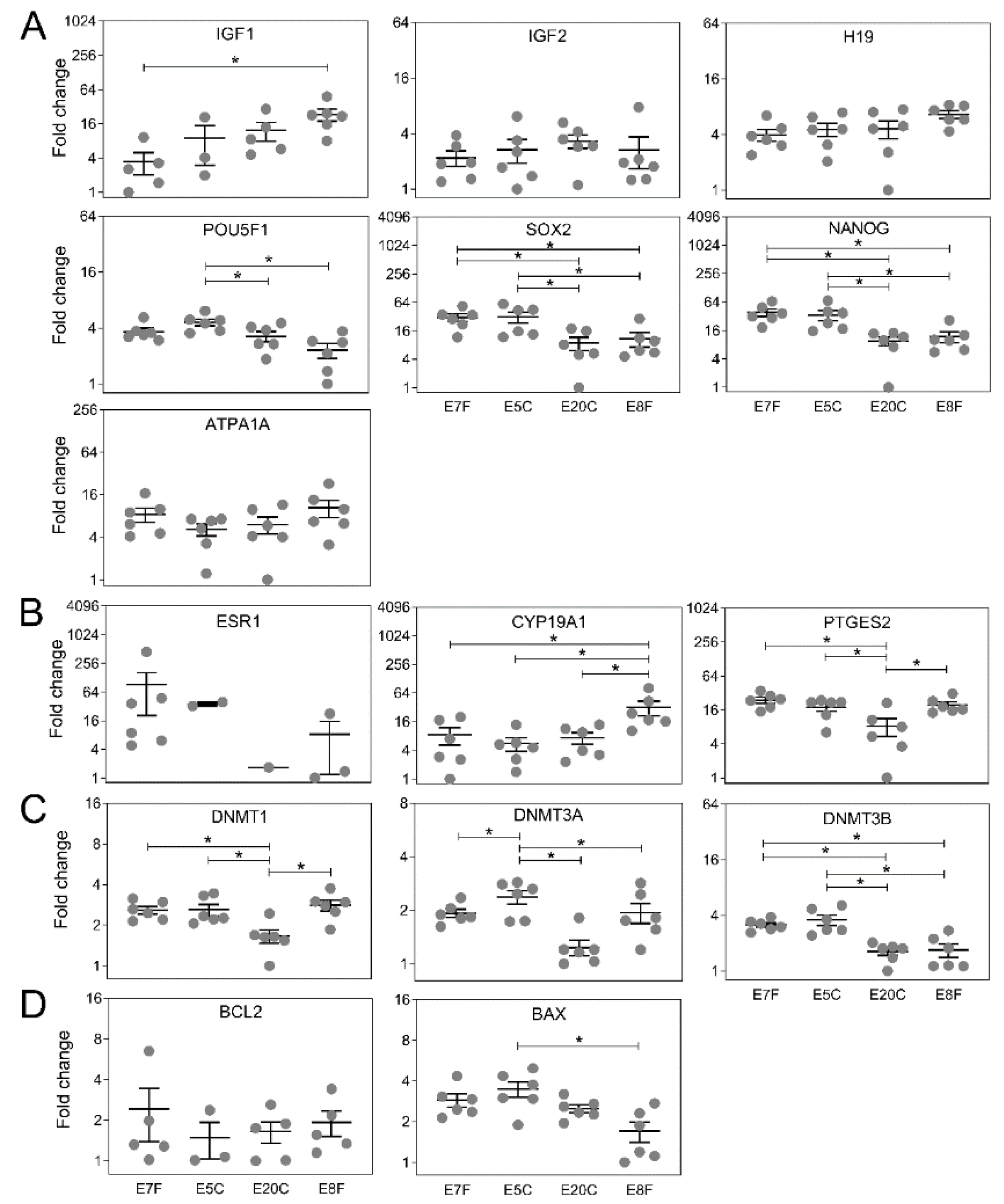
3. Results
3.1. Recovery Rate and Embryo Morphology
3.2. Medium Temperature, pH, and Lipid Peroxidation
3.3. Gene Expression
3.4. Gene-Specific DNA Methylation
3.5. Global methylation–5hmC and 5mC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Squires, E. Current reproductive technologies impacting equine embryo production. J. Equine Vet. Sci. 2020, 89, 102981. [Google Scholar] [CrossRef] [PubMed]
- Hinrichs, K.; Choi, Y.-H. Equine embryo biopsy, genetic testing, and cryopreservation. J. Equine Vet. Sci. 2012, 32, 390–396. [Google Scholar] [CrossRef]
- Foss, R.; Ortis, H.; Hinrichs, K. Effect of potential oocyte transport protocols on blastocyst rates after intracytoplasmic sperm injection in the horse. Equine Vet. J. 2013, 45, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Squires, E.L.; Carnevale, E.M.; McCue, P.M.; Bruemmer, J.E. Embryo technologies in the horse. Theriogenology 2003, 59, 151–170. [Google Scholar] [CrossRef]
- Watson, A.J.; Westhusin, M.E.; de Sousa, P.A.; Betts, D.H.; Barcroft, L.C. Gene expression regulating blastocyst formation. Theriogenology 1999, 51, 117–133. [Google Scholar] [CrossRef]
- Rader, K.; Choi, Y.-H.; Hinrichs, K. Intracytoplasmic sperm injection, embryo culture, and transfer of in vitro–produced blastocysts. Vet. Clin. Equine Pract. 2016, 32, 401–413. [Google Scholar] [CrossRef]
- O’Neill, C. The epigenetics of embryo development. Anim. Front. 2015, 5, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [Green Version]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef]
- Reik, W.; Dean, W.; Walter, J. Epigenetic reprogramming in mammalian development. Science 2001, 293, 1089–1093. [Google Scholar] [CrossRef] [Green Version]
- Vrooman, L.A.; Rhon-Calderon, E.A.; Chao, O.Y.; Nguyen, D.K.; Narapareddy, L.; Dahiya, A.K.; Putt, M.E.; Schultz, R.M.; Bartolomei, M.S. Assisted reproductive technologies induce temporally specific placental defects and the preeclampsia risk marker sFLT1 in mouse. Development 2020, 147. [Google Scholar] [CrossRef]
- Schieve, L.A.; Cohen, B.; Nannini, A.; Ferre, C.; Reynolds, M.A.; Zhang, Z.; Jeng, G.; Macaluso, M.; Wright, V.C. A population-based study of maternal and perinatal outcomes associated with assisted reproductive technology in Massachusetts. Matern. Child Health J. 2007, 11. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.; Moley, K.H. Gametes and embryo epigenetic reprogramming affect developmental outcome: Implication for assisted reproductive technologies. Pediatr. Res. 2005, 58, 437–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Robbins, K.M.; Wells, K.D.; Rivera, R.M. Large offspring syndrome: A bovine model for the human loss-of-imprinting overgrowth syndrome Beckwith-Wiedemann. Epigenetics 2013, 8, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Leese, H.J. Metabolism of the preimplantation embryo: 40 years on. Reproduction 2012, 143, 417–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCaw, B.A.; Stevenson, T.J.; Lancaster, L.T. Epigenetic responses to temperature and climate. Integr. Comp. Biol. 2020, 60, 1469–1480. [Google Scholar] [CrossRef]
- Canovas, S.; Ivanova, E.; Romar, R.; García-Martínez, S.; Soriano-Ubeda, C.; García-Vázquez, F.A.; Saadeh, H.; Andrews, S.; Kelsey, G.; Coy, P. DNA methylation and gene expression changes derived from assisted reproductive technologies can be decreased by reproductive fluids. Elife 2017, 6, e23670. [Google Scholar] [CrossRef] [PubMed]
- Canovas, S.; Ross, P.J.; Kelsey, G.; Coy, P. DNA methylation in embryo development: Epigenetic impact of ART (assisted reproductive technologies). Bioessays 2017, 39, 1700106. [Google Scholar] [CrossRef]
- Bonometti, S.; Menarim, B.C.; Reinholt, B.M.; Ealy, A.D.; Johnson, S.E. Growth factor modulation of equine trophoblast mitosis and prostaglandin gene expression. J. Anim. Sci. 2019, 97, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Breton-Larrivée, M.; Elder, E.; McGraw, S. DNA methylation, environmental exposures and early embryo development. Anim. Reprod. 2019, 16, 465–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Li, E. Chromatin modification and epigenetic reprogramming in mammalian development. Nat. Rev. Genet. 2002, 3, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Suetake, I.; Shinozaki, F.; Miyagawa, J.; Takeshima, H.; Tajima, S. DNMT3L stimulates the DNA methylation activity of Dnmt3a and Dnmt3b through a direct interaction. J. Biol. Chem. 2004, 279, 27816–27823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, Y.; Yoshida, N.; Suzuki, T.; Shimozawa, N.; Asami, M.; Matsuda, T.; Kojima, N.; Perry, A.C.F.; Takada, T. DNA methylation dynamics in mouse preimplantation embryos revealed by mass spectrometry. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lucas, E. Epigenetic effects on the embryo as a result of periconceptional environment and assisted reproduction technology. Reprod. Biomed. Online 2013, 27, 477–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.G.; Lane, M.; Gilchrist, R.B. Metabolism of the bovine cumulus-oocyte complex and influence on subsequent developmental competence. Soc. Reprod. Fertil. Suppl. 2007, 64, 179. [Google Scholar] [CrossRef] [PubMed]
- Urrego, R.; Rodriguez-Osorio, N.; Niemann, H. Epigenetic disorders and altered gene expression after use of assisted reproductive technologies in domestic cattle. Epigenetics 2014, 9, 803–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, M.; Smith, O.E.; Therrien, J.; Rigoglio, N.N.; Miglino, M.A.; Silva, L.A.; Meirelles, F.V.; Smith, L.C. Resiliency of equid H19 imprint to somatic cell reprogramming by oocyte nuclear transfer and genetically induced pluripotency. Biol. Reprod. 2020, 102, 211–219. [Google Scholar] [CrossRef]
- Grant, D.M.; Macedo, A.; Toms, D.; Klein, C. Fibrinogen in equine pregnancy as a mediator of cell adhesion, an epigenetic and functional investigation. Biol. Reprod. 2020, 102, 170–184. [Google Scholar] [CrossRef]
- Aurich, C. Reproductive cycles of horses. Anim. Reprod. Sci. 2011, 124, 220–228. [Google Scholar] [CrossRef]
- Aurich, J.E.; Schönherr, U.; Hoppe, H.; Aurich, C. Effects of antioxidants on motility and membrane integrity of chilled-stored stallion semen. Theriogenology 1997, 48, 185–192. [Google Scholar] [CrossRef]
- McKinnon, A.O.; Squires, E.L. Equine embryo transfer. Vet. Clin. North Am. Equine Pract. 1988, 4, 305–333. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Beckelmann, J.; Budik, S.; Helmreich, M.; Palm, F.; Walter, I.; Aurich, C. Sex-dependent insulin like growth factor-1 expression in preattachment equine embryos. Theriogenology 2013, 79, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Hackett, C.H.; Greve, L.; Novakofski, K.D.; Fortier, L.A. Comparison of gene-specific DNA methylation patterns in equine induced pluripotent stem cell lines with cells derived from equine adult and fetal tissues. Stem Cells Dev. 2012, 21, 1803–1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhne, M.; Ille, N.; Erber, R.; Razavi, M.S.A.; Walter, I.; Aurich, C. Effects of periovulatory gonadotrophin treatment on luteal function and endometrial expression of selected genes in cyclic pony mares. Theriogenology 2016, 86, 2147–2155. [Google Scholar] [CrossRef] [PubMed]
- Scarlet, D.; Ille, N.; Ertl, R.; Alves, B.G.; Gastal, G.D.A.; Paiva, S.O.; Gastal, M.O.; Gastal, E.L.; Aurich, C. Glucocorticoid metabolism in equine follicles and oocytes. Domest. Anim. Endocrinol. 2017, 59, 11–22. [Google Scholar] [CrossRef]
- Scarlet, D.; Ertl, R.; Aurich, C.; Steinborn, R. The orthology clause in the next generation sequencing era: Novel reference genes identified by RNA-seq in humans improve normalization of neonatal equine ovary RT-qPCR data. PLoS ONE 2015, 10, e0142122. [Google Scholar] [CrossRef] [PubMed]
- Kenkel, C.D.; Aglyamova, G.; Alamaru, A.; Bhagooli, R.; Capper, R.; Cunning, R.; deVillers, A.; Haslun, J.A.; Hédouin, L.; Keshavmurthy, S. Development of gene expression markers of acute heat-light stress in reef-building corals of the genus Porites. PLoS ONE 2011, 6, e26914. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Heinzmann, J.; Mattern, F.; Aldag, P.; Bernal-Ulloa, S.M.; Schneider, T.; Haaf, T.; Niemann, H. Extended in vitro maturation affects gene expression and DNA methylation in bovine oocytes. Mhr Basic Sci. Reprod. Med. 2015, 21, 770–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.-C.; Dahiya, R. MethPrimer: Designing primers for methylation PCRs. Bioinformatics 2002, 18, 1427–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, F.; Dean, W. Using immunofluorescence to observe methylation changes in mammalian preimplantation embryos. In Nuclear Reprogramming; Springer: Berlin/Heidelberg, Germany, 2006; pp. 129–138. [Google Scholar]
- Heras, S.; Smits, K.; de Schauwer, C.; van Soom, A. Dynamics of 5-methylcytosine and 5-hydroxymethylcytosine during pronuclear development in equine zygotes produced by ICSI. Epigenetics Chromatin 2017, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colchen, S.; Battut, I.; Fiéni, F.; Tainturier, D.; Siliart, B.; Bruyas, J.F. Quantitative histological analysis of equine embryos at exactly 156 and 168 h after ovulation. J. Reprod. Fertil. Suppl. 2000, 56, 527–537. [Google Scholar]
- Panzani, D.; Rota, A.; Marmorini, P.; Vannozzi, I.; Camillo, F. Retrospective study of factors affecting multiple ovulations, embryo recovery, quality, and diameter in a commercial equine embryo transfer program. Theriogenology 2014, 82, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Aurich, C.; Budik, S. Early pregnancy in the horse revisited–does exception prove the rule? J. Anim. Sci. Biotechnol. 2015, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Waelchli, R.O.; MacPhee, D.J.; Kidder, G.M.; Betteridge, K.J. Evidence for the presence of sodium- and potassium-dependent adenosine triphosphatase α1 and β1 subunit isoforms and their probable role in blastocyst expansion in the preattachment horse conceptus. Biol. Reprod. 1997, 57, 630–640. [Google Scholar] [CrossRef] [Green Version]
- Budik, S.; Walter, I.; Tschulenk, W.; Helmreich, M.; Deichsel, K.; Pittner, F.; Aurich, C. Significance of aquaporins and sodium potassium ATPase subunits for expansion of the early equine conceptus. Reproduction 2008, 135, 497–508. [Google Scholar] [CrossRef] [Green Version]
- Esteves, C.L.; Sharma, R.; Dawson, L.; Taylor, S.E.; Pearson, G.; Keen, J.A.; McDonald, K.; Aurich, C.; Donadeu, F.X. Expression of putative markers of pluripotency in equine embryonic and adult tissues. Vet. J. 2014, 202, 533–535. [Google Scholar] [CrossRef]
- Gagliardi, A.; Mullin, N.P.; Ying Tan, Z.; Colby, D.; Kousa, A.I.; Halbritter, F.; Weiss, J.T.; Felker, A.; Bezstarosti, K.; Favaro, R.; et al. A direct physical interaction between Nanog and Sox2 regulates embryonic stem cell self-renewal. EMBO J. 2013, 32. [Google Scholar] [CrossRef] [Green Version]
- Herrler, A.; Krusche, C.A.; Beier, H.M. Insulin and insulin-like growth factor-I promote rabbit blastocyst development and prevent apoptosis. Biol. Reprod. 1998, 59, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.M.; Feng, H.L.; Ma, Y.Z.; Cang, M.; Li, H.J.; Yan, Z.; Zhou, P.; Wen, J.X.; Bou, S.; Liu, D.J. Expression of IGF receptors and its ligands in bovine oocytes and preimplantation embryos. Anim. Reprod. Sci. 2009, 114, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Bragulat, A.F.; Llorente, J.; Morikawa, I.; von Meyeren, M.; Losinno, L. Pregnancy rates for cooled transported equine embryos according to size and arrival temperature. J. Equine Vet. Sci. 2020, 89, 103078. [Google Scholar] [CrossRef]
- Monk, M.; Boubelik, M.; Lehnert, S. Temporal and regional changes in DNA methylation in the embryonic, extraembryonic and germ cell lineages during mouse embryo development. Development 1987, 99, 371–382. [Google Scholar] [CrossRef]
- Edwards, J.R.; Yarychkivska, O.; Boulard, M.; Bestor, T.H. DNA methylation and DNA methyltransferases. Epigenetics Chromatin 2017, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinho, L.S.R.; Rissi, V.B.; Lindquist, A.G.; Seneda, M.M.; Bordignon, V. Acetylation and methylation profiles of H3K27 in porcine embryos cultured in vitro. Zygote 2017, 25, 575–582. [Google Scholar] [CrossRef]
- Bestor, T.H.; Edwards, J.R.; Boulard, M. Notes on the role of dynamic DNA methylation in mammalian development. Proc. Natl. Acad. Sci. USA 2015, 112, 6796–6799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, L.E.; Sinclair, K.D.; Wilmut, I. Large offspring syndrome in cattle and sheep. Rev. Reprod. 1998, 3, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Kleijkers, S.H.M.; van Montfoort, A.P.A.; Smits, L.J.M.; Viechtbauer, W.; Roseboom, T.J.; Nelissen, E.C.M.; Coonen, E.; Derhaag, J.G.; Bastings, L.; Schreurs, I.E.L.; et al. IVF culture medium affects post-natal weight in humans during the first 2 years of life. Hum. Reprod. 2014, 29, 661–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Gonzalez, R.; Moreira, P.; Bilbao, A.; Jiménez, A.; Pérez-Crespo, M.; Ramírez, M.A.; de Fonseca, F.R.; Pintado, B.; Gutiérrez-Adán, A. Long-term effect of in vitro culture of mouse embryos with serum on mRNA expression of imprinting genes, development, and behavior. Proc. Natl. Acad. Sci. USA 2004, 101, 5880–5885. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | NCBI/Ensembl Accession Number | Oligo Sequence (5′–3′) | Amplicon Length (bp) | PCR Efficiency (%) | R2 Value | Reference |
|---|---|---|---|---|---|---|---|
| ATP1A1 | Equus caballus ATPase Na+/K+ transporting subunit alpha 1 | NM_001114532.2, XM_023640223.1, XM_023640224.1 | F: CTTGATGAACTTCAGCGCAAATA | 104 | 94 | 0.990 | This study |
| R: GGTGTAAGGGCATTGGGA | |||||||
| P: TGAGCCGAGGCTTAACAACTGCTC | |||||||
| BAX | Equus caballus BCL2-associated X protein | XM_014729721.1 | F: AGGATGCGTCCACCAAGAAG | 80 | 93.2 | 0.994 | [37] |
| XM_014729717.1 | R: CCTCTGCAGCTCCATGTTACTG | ||||||
| P: CTCAAGCGCATCGGAGATGAGCTG | |||||||
| BCL2 | Equus caballus B-cell lymphoma 2 | XM_001490436.2 | F: TTGGAAAGCCTACCACTAATTGC | 74 | 92.6 | 0.998 | [37] |
| R: CCGTGTTTATAGGCACAGGAGAT | |||||||
| P: CCCACCTGAGCGGCTCCACC | |||||||
| CYP19A1 | Equus caballus cytochrome P450 family 19 subfamily A member 1 | NM_001081805.2 | F: GGAGAGGAAACGCTCGTTATTA | 107 | 99.2 | 0.999 | This study |
| XM_005602588.2 | R: CCCATATACTGCAACCCAAATG | ||||||
| XM_005602587.2 | P: ATCACTACTCCTCCCGATTTGGCA | ||||||
| DNMT1 | Equus caballus DNA methyltransferase 1 | XM_014741825.1 | F: GACCACCATCACGTCTCATTT | 97 | 100.5 | 1 | This study |
| R: CTCCTCATCCACAGAATTGTCC | |||||||
| P: AAACGGAAACCCGAGGAAGAGCTG | |||||||
| DNMT3A | Equus caballus DNA methyltransferase 3 α | XM_005600169.2 | F: GATTATTGACGAACGCACAAGAG | 112 | 100 | 0.998 | This study |
| XM_005600168.2 | |||||||
| XM_005600167.2 | |||||||
| XM_005600170.2 | R: GTGTTCCAGGGTGACATTGA | ||||||
| XM_005600171.2 | P: TGCAAATGTCTTCGATGTTCCGGC | ||||||
| DNMT3B | Equus caballus DNA methyltransferase 3 β | XM_001916514.4 | F: CGAGTCTTGTCCCTGTTTGAT | 110 | 100.6 | 0.999 | This study |
| R: GCGATAGACTCTTCACACACTT | |||||||
| P: CGCCACAGGGTACTTGGTTCTCAA | |||||||
| DNMT3L | Equus caballus DNA (cytosine-5-)-methyltransferase 3-like | XM_014736476.1 | F: GCCCTCACTTGGTTGGTTT | 98 | 100.5 | 0.999 | This study |
| R: CTTCCACACAGGCACAGTTT | |||||||
| P: CAAAGTGCCCATCTGCTCTGGAGA | |||||||
| ESR1 | Equus caballus estrogen receptor 1 | NM_001081772.1 | F: CACCCAGGAAAGCTCCTATTT | 110 | 101.8 | 0.999 | This study |
| R: CGAGATGACGTAGCCAACAA | |||||||
| P: TCCACCATGCCCTCTACACATTTCC | |||||||
| H19 | Equus caballus H19, imprinted maternally expressed transcript | NR_027326.2 | F: CCTCTAGCTCTGACTCAAGAATATG | 103 | 94.3 | 0.992 | This study |
| R: CAGGTCCATCTGGTTCCTTTAG | |||||||
| P: ACTCAGGAATCAGCTCTGGAAGGT | |||||||
| IGF1 | Equus caballus insulin-like growth factor 1 | NM_001082498.2 | F: TGCTTCCGGAGCTGTGATCT | 67 | 102 | 1 | [34] |
| XM_005606471.2 | |||||||
| XM_005606472.2 | |||||||
| XM_005606470.2 | R: CCGACTTGGCAGGCTTGA | ||||||
| XM_005606469.2 | P: AGGAGGCTGGAGATGTACTGCGCACC | ||||||
| IGF2 | Equus caballus insulin-like growth factor 2 | NM_001114539.2 | F: AAGTCCGAGAGGGACGTG | 100 | 99.9 | 0.998 | This study |
| R: ATTGCTTCCAGGCGTTGT | |||||||
| P: CCCGTGGTCAAGCTCTTCCAGT | |||||||
| NANOG | Equus caballus Nanog homeobox | XM_014740545.1 | F: ACAGCCCCGATTCATCCA | 72 | 102.3 | 0.999 | [35] |
| R: TCTTTGCCTCGCTCGTCTCT | |||||||
| XM_001498808.1 | P: CAGTCCCAGAGTAAAACCGCTGCCC | ||||||
| POU5F1 | Equus caballus POU class 5 homeobox 1 | XM_014734675.1 | F: CGGGCACTGCAGGAACAT | 73 | 100.8 | 0.999 | [35] |
| R: CCGAAAGAGAAAGCGAACTAGTATTG | |||||||
| XM_001490108.5 | P: TTCTCCAGGTTGCCTCTCACTCGGTTC | ||||||
| PSMB4 | Equus caballus proteasome subunit beta type IV | XM_001492317.4 | F: CTTGGTGTAGCCTATGAAGCCC | 82 | 93.1 | 0.991 | [38] |
| XM_005610132.1 | |||||||
| XM_008515015.1 | |||||||
| XM_005613704.1 | R: CCAGAATTTCTCGCAGCAGAG | ||||||
| PTGES2 | Equus caballus prostaglandin-endoperoxide synthase 2 | NM_001081775.2 | F: GAGGTGTATCCGCCCACAGT | 81 | 92.3 | 0.996 | [36] |
| R: AGCAAACCGCAGGTGCTC | |||||||
| P: TCAGATGGAAATGATCTACCCGCCTCA | |||||||
| SNRPD3 | Equus caballus small nuclear ribonucleoprotein D3 polypeptide. | XM_001489060.4 | F: ACGCACCTATGTTAAAGAGCATG | 120 | 99.4 | 0.996 | [38] |
| XM_008511652.1 | R: CACGTCCCATTCCACGTC | ||||||
| SOX2 | Equus caballus SRY box 2 | XM_003363345.3 | F: TGCGAGCGCTGCACAT | 91 | 99.3 | 0.998 | [35] |
| R: AGCGTGTACTTATCCTTCTTCATGAG | |||||||
| P: ATAAATACCGTCCTCGGCGGAAAACCAA |
| Gene | Primer Sequence (5′–3′) | Amplicon Length (bp) | Genomic Localization (EquCab3.0) | Number of CpGs in Inner Amplicon | Number of Sequenced CpGs |
|---|---|---|---|---|---|
| CYP19A1 | Outer forward: TTTTAGTTTTGATTGGTTGTTTTT | 317 | 1:140151317-140151633 | - | - |
| Outer reverse: CTAAACCCCATAAAACATCTCTTAC | |||||
| Inner forward: TTTTTTTTGTAAGATTAGTGAGTATATTTA | 212 | 1:140151386-140151597 | 4 | 3 | |
| Inner reverse: TTTCCAAAATTAAAAAACATAACC | |||||
| DNMT1 | Outer forward: AATTTTTTTTAAGAGTTTGGTATGG | 264 | 7:51536766-51537029 | - | - |
| Outer reverse: ACCAATCCTCCTCTTTATACTAAAA | |||||
| Inner forward: GAGTTTGGTATGGTATATAAGTGTTGA | 229 | 7:51536778-51537006 | 9 | 8 | |
| Inner reverse: AAAAAACTAACCCTAAACTCACATC | |||||
| DNMT3A | Outer forward: GGGATTGATTAGATTTTTTAGAGAAG | 316 | 15:71656615-71656930 | - | - |
| Outer reverse: TAATAACACTAAATCCCTCCAAAAC | |||||
| Inner forward: TAGGAGTTTAGTGGGGGAATAGT | 200 | 15:71656665-71656864 | 6 | 4 | |
| Inner Reverse: ATAAAATAAATAAAACCCCTACACC | |||||
| DNMT3B | Outer forward: TTAAAGGGGGAATAGTAGAAGTTTA | 388 | 22:24214245-24214632 | - | - |
| Outer reverse: CAACTCCAAAAATATTTAAAATCAC | |||||
| Inner forward: TATAGAGGATGGATTTGGGATTTTA | 240 | 22:24214309-24214548 | 10 | 8 | |
| Inner reverse: ACTAAACACTCCCTACCCTAATACC | |||||
| ESR1 | Outer forward: TTGTGGTAGGTATGAATATTTATGTG | 334 | 31:15363114-15363447 | - | - |
| Outer reverse: ATTACATATACAACCAACCACAAAC | |||||
| Inner forward: AATTTTTAGTGGGAGGAAGTATAGTAT | 226 | 31:15363156-15363381 | 9 | 8 | |
| Inner reverse: ACATAAACTAACAAAAAACATCCC | |||||
| NANOG | Outer forward: TGGAAATATGGTGAATTTATAGGTAT | 387 | 6:36542573-36542959 | - | - |
| Outer reverse: AACTTAAATATCCAAACAAAAAACC | |||||
| Inner forward: TTGGTAGATAGGATTAATTGAGAATT | 237 | 6:36542585-36542821 | 8 | 7 | |
| Inner reverse: CAAACAAAAAACCTTAAAAAAATAC | |||||
| PTGES2 | Forward: GATTTATTTAAGAGTGGGGGAGGT | 205 | 25:32059289-32059493 | 17 | 11 |
| Reverse: CAATATAAAACCCCAACC | |||||
| SOX2 | Outer forward: ATTTTTAATATAGAATAAATTATGGAGAAG | 302 | 19:22733114-22733417 | - | - |
| Outer reverse: AAATAAAAATAAAACAAAACAAAATAAATA | |||||
| Inner forward: ATAGAATAAATTATGGAGAAGTAAGGAG | 253 | 19:22733122-22733376 | 19 | 13 | |
| Inner reverse: CTATCCTACTAAAATTTCAAAAACC |
| Function | Growth & Development | Embryo-Maternal Communication | Methylation | Apoptosis | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | IGF2- | H19 | POU5F1 | SOX2 | IGF1 | NANOG | ATPA1A | ESR1 | CYP19A1 | PTGES2 | DNMT1 | DNMT3a | DNMT3b | BCL2 | BAX | |
| Growth & Development | IGF2 | 1 | ||||||||||||||
| H19 | 0.4 * | 1 | ||||||||||||||
| POU5F1 | 0.26 | −0.05 | 1 | |||||||||||||
| SOX2 | 0.21 | −0.14 | 0.74 *** | 1 | ||||||||||||
| IGF1 | −0.06 | 0.34 | 0.74 *** | −0.66 ** | 1 | |||||||||||
| NANOG | 0.07 | −0.27 | 0.73 *** | 0.89 *** | −0.63 ** | 1 | ||||||||||
| ATPA1A | −0.04 | 0.06 | −0.66 *** | −0.46 * | 0.56 * | −0.34 | 1 | |||||||||
| Embryo-maternal communication | ESR1 | 0.04 | −0.05 | 0.33 | 0.45 | −0.43 | 0.2 | 0.18 | 1 | |||||||
| CYP19A1 | −0.24 | 0.4 * | −0.74 *** | -0.59 ** | 0.49 * | −0.51 * | 0.5 * | −0.12 | 1 | |||||||
| PTGES2 | 0.14 | 0.16 | −0.1 | 0.14 | 0.03 | 0.26 | 0.37 | 0.15 | 0.1 | 1 | ||||||
| Methylation | DNMT1 | −0.23 | 0.29 | −0.11 | 0.1 | 0.49 * | 0.19 | 0.45 * | 0.27 | 0.47 * | 0.7 *** | 1 | ||||
| DNMT3a | −0.12 | 0.19 | 0.32 | 0.5 | 0.02 | 0.58 ** | 0.14 | 0.54 | −0.04 | 0.45 * | 0.65 *** | 1 | ||||
| DNMT3b | −0.04 | −0.05 | 0.75 *** | 0.82 *** | −0.59 ** | 0.91 *** | −0.36 | 0.29 | −0.39 | 0.3 | 0.29 | 0.57 ** | 1 | |||
| Apoptosis | BCL2 | −0.04 | 0.24 | 0.21 | 0.05 | 0.01 | 0.14 | 0.03 | 0.38 | 0.09 | −0.07 | 0.23 | 0.43 | 0.07 | 1 | |
| BAX | 0.07 | −0.22 | 0.77 *** | 0.55 ** | −0.69 ** | 0.58 ** | −0.6 ** | 0.34 | −0.54 ** | −0.04 | −0.12 | 0.05 | 0.67 *** | −0.02 | 1 | |
| Function | Gene | E7F | E5C | E20C | E8F | p-Value |
|---|---|---|---|---|---|---|
| Growth & Development | SOX2 | 2.2 | 1.1 | 0.0 | 0.0 | 0.62 |
| NANOG | 46.9 a | 63.3 a,b | 53.1 a | 85.7 b | <0.0001 | |
| Embryo-maternal communication | ESR1 | 64.3 a,b | 39.3 b,c | 28.6 c | 78.6 a | <0.0001 |
| CYP19A1 | 100.0 | 100.0 | 100.0 | 100.0 | NS | |
| PTGES2 | 0.0 | 0.0 | 0.0 | 0.0 | NS | |
| Methylation | DNMT1 | 89.3 a,b | 85.7 a,b | 75.0 b | 96.4 a | 0.009 |
| DNMT3a | 78.6 a | 100.0 b | 89.3 a,b | 100.0 b | 0.006 | |
| DNMT3b | 53.6 | 30.4 | 44.6 | 33.9 | 0.052 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gastal, G.D.A.; Scarlet, D.; Melchert, M.; Ertl, R.; Aurich, C. Epigenetic Changes in Equine Embryos after Short-Term Storage at Different Temperatures. Animals 2021, 11, 1325. https://doi.org/10.3390/ani11051325
Gastal GDA, Scarlet D, Melchert M, Ertl R, Aurich C. Epigenetic Changes in Equine Embryos after Short-Term Storage at Different Temperatures. Animals. 2021; 11(5):1325. https://doi.org/10.3390/ani11051325
Chicago/Turabian StyleGastal, Gustavo D. A., Dragos Scarlet, Maria Melchert, Reinhard Ertl, and Christine Aurich. 2021. "Epigenetic Changes in Equine Embryos after Short-Term Storage at Different Temperatures" Animals 11, no. 5: 1325. https://doi.org/10.3390/ani11051325