
Effects of Pasture Grass, Silage, and Hay Diet on Equine Fecal Microbiota



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethnic Announcement
2.2. Animal Selection and Experimental Design
2.3. Fecal Sampling
2.4. DNA Preparation
2.5. High-Throughput Sequencing
2.6. Bioinformatics Analysis
3. Results
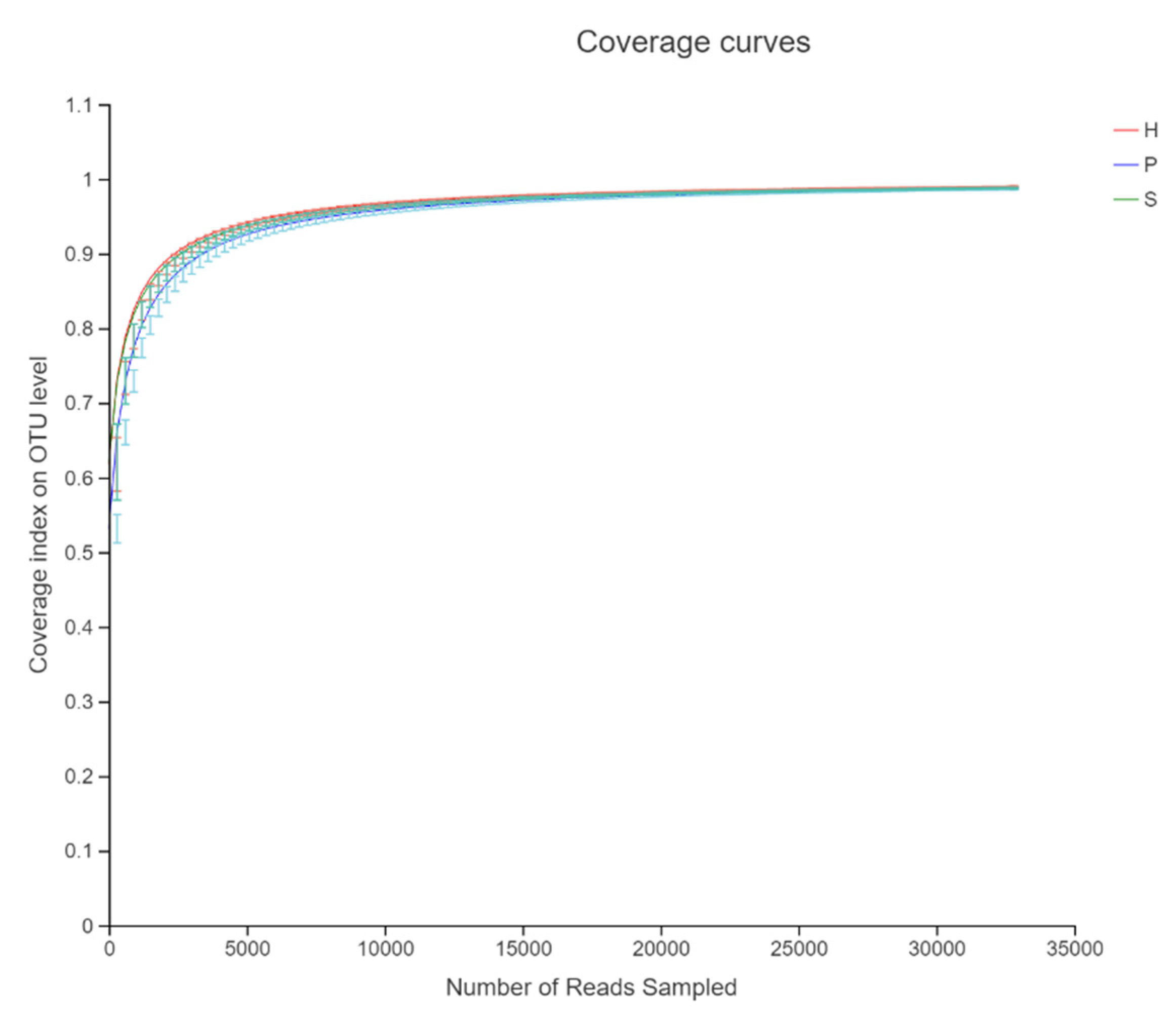
3.1. Sequencing Quality Data
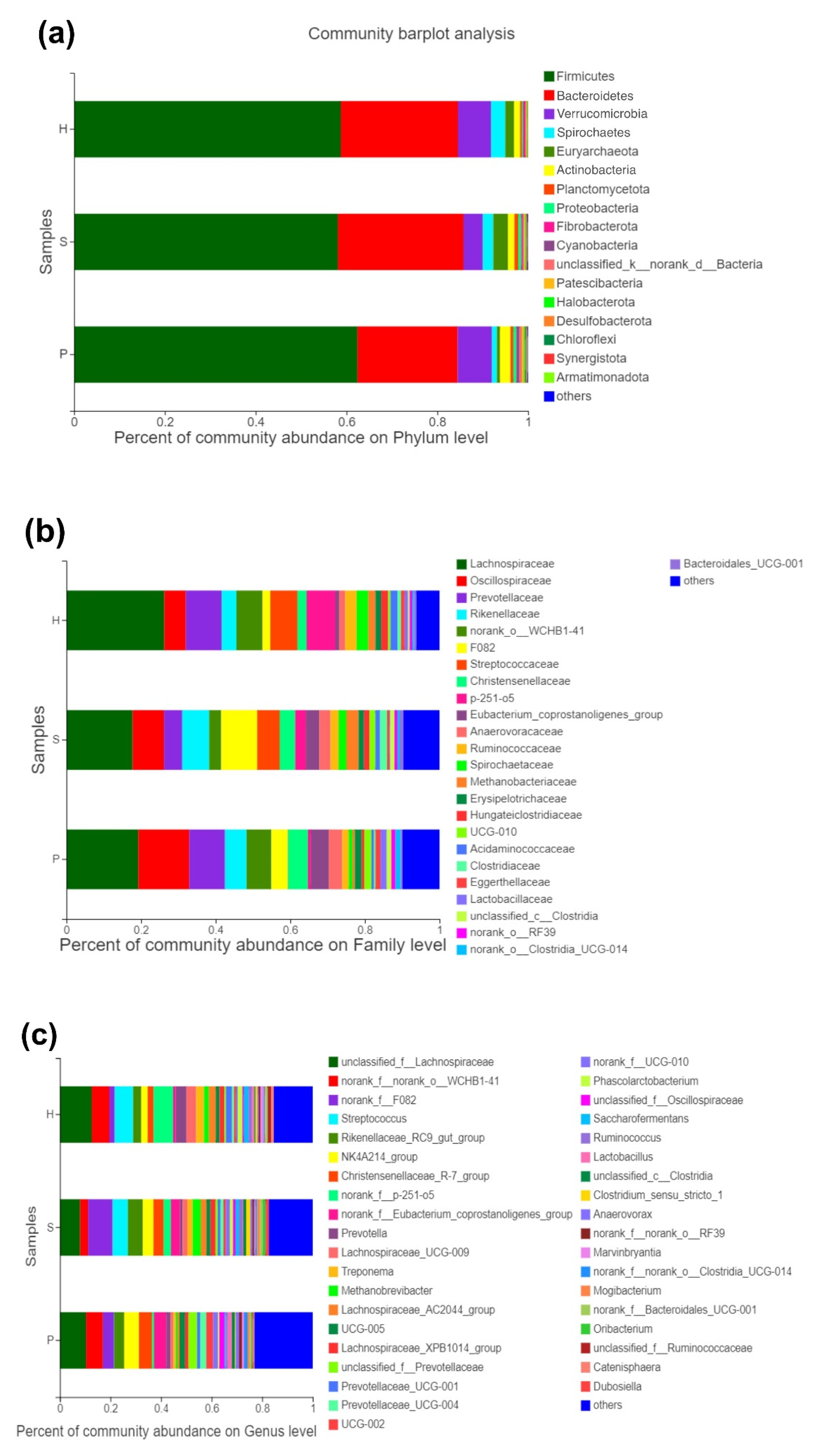
3.2. Microbial Composition Analysis
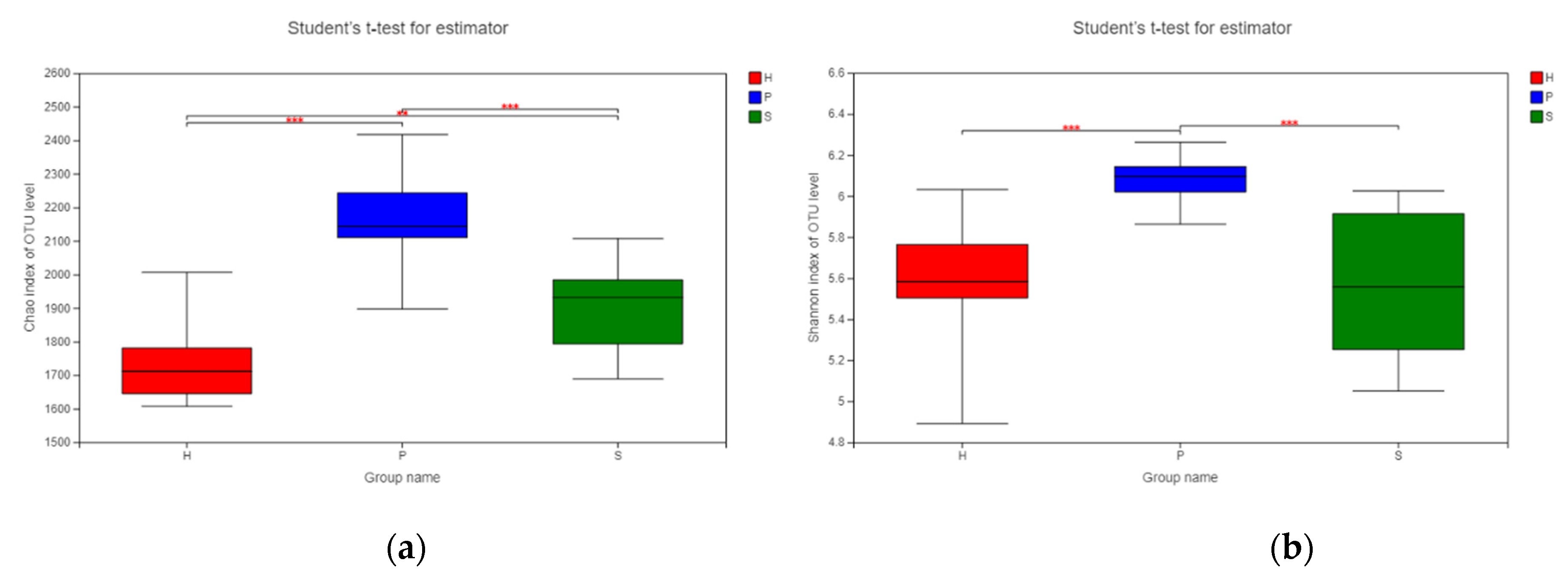
3.3. Alpha Diversity Analysis
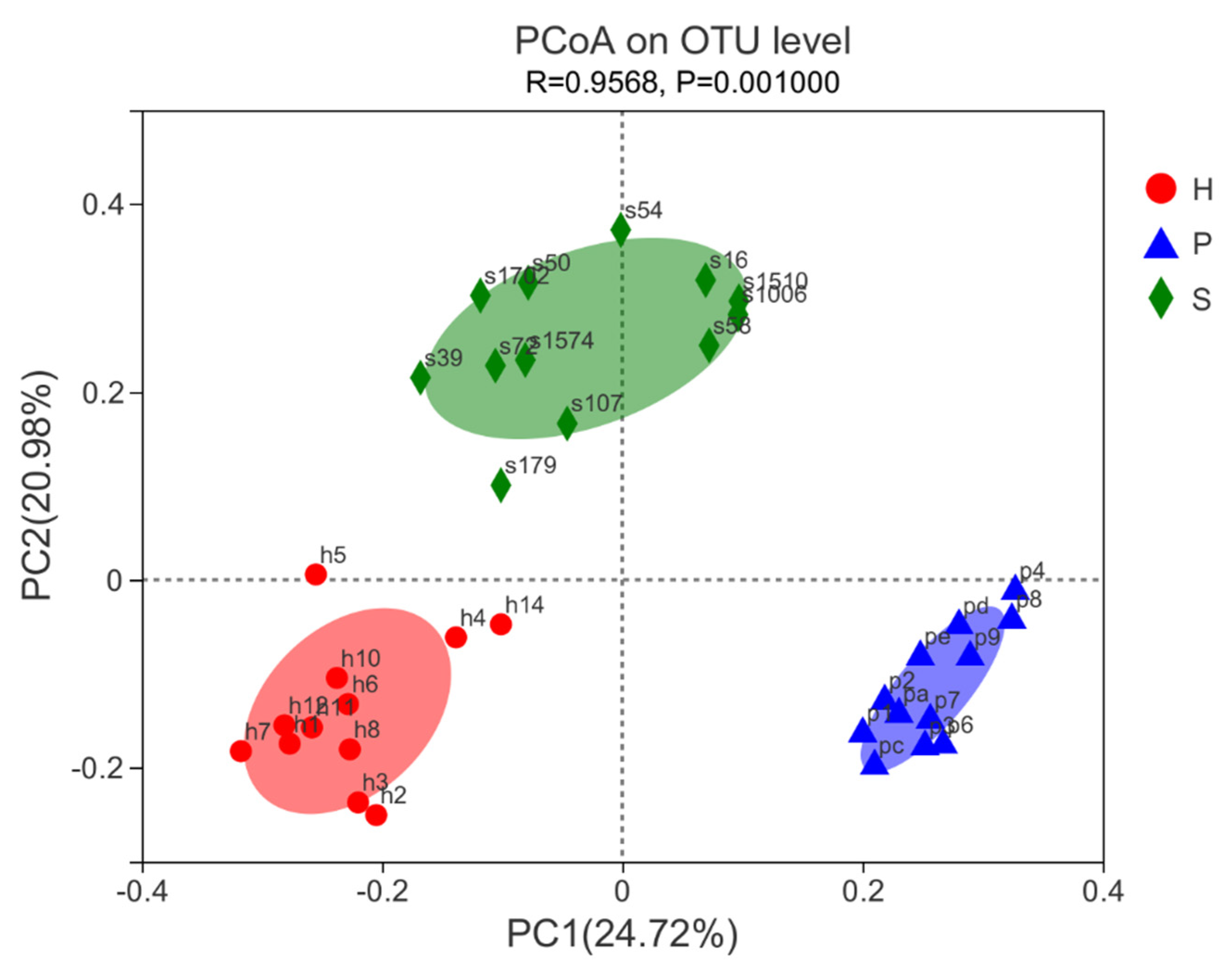
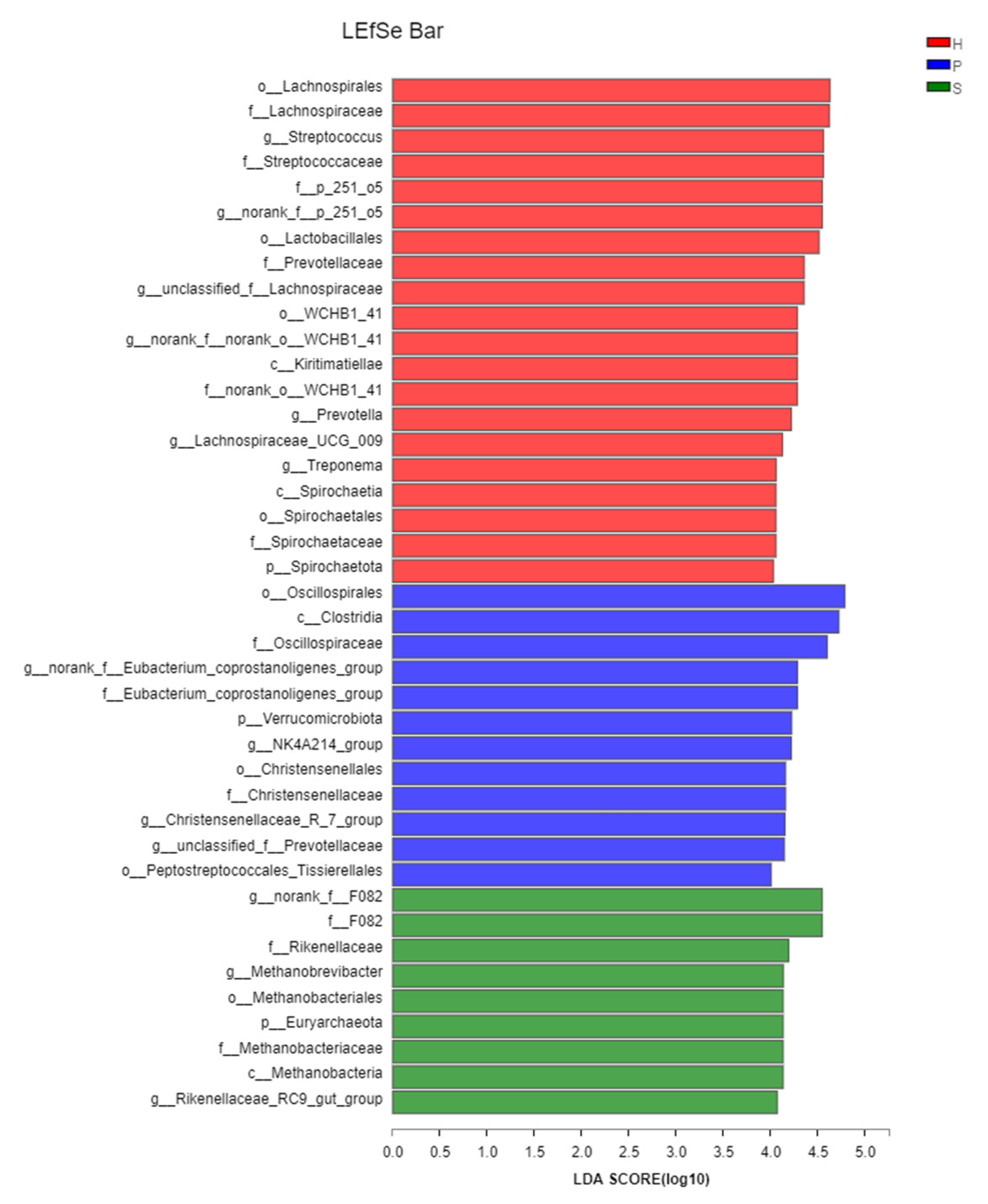
3.4. Beta Diversity and LEfSe Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hintz, H.; Cymbaluk, N. Nutrition of the horse. Annu. Rev. Nutr. 1994, 14, 243–267. [Google Scholar] [CrossRef]
- Julliand, V.; de Fombelle, A.; Drogoul, C.; Jacotot, E. Feeding and microbial disorders in horses: Part 3—Effects of three hay: Grain ratios on microbial profile and activities. J. Equine Vet. Sci. 2001, 21, 543–546. [Google Scholar] [CrossRef]
- Hintz, H.F.; Hogue, D.E.; Walker, E.F., Jr.; Lowe, J.E.; Schryver, H.F. Apparent digestion in various segments of the digestive tract of ponies fed diets with varying roughage-grain ratios. J. Anim. Sci. 1971, 32, 245–248. [Google Scholar] [CrossRef]
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.J.; Wu, E. The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dougal, K.; de la Fuente, G.; Harris, P.A.; Girdwood, S.E.; Pinloche, E.; Geor, R.J.; Nielsen, B.D.; Schott II, H.C.; Elzinga, S.; Newbold, C.J. Characterisation of the faecal bacterial community in adult and elderly horses fed a high fibre, high oil or high starch diet using 454 pyrosequencing. PLoS ONE 2014, 9, e87424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julliand, V.; Grimm, P. The impact of diet on the hindgut microbiome. J. Equine Vet. Sci. 2017, 52, 23–28. [Google Scholar] [CrossRef]
- Salem, S.E.; Maddox, T.W.; Berg, A.; Antczak, P.; Ketley, J.M.; Williams, N.J.; Archer, D.C. Variation in faecal microbiota in a group of horses managed at pasture over a 12-month period. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, M.L.; Swecker, W.S., Jr.; Jensen, R.V.; Ponder, M.A. Characterization of the fecal bacteria communities of forage-fed horses by pyrosequencing of 16s rrna v4 gene amplicons. FEMS Microbiol. Lett. 2012, 326, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Scott, K.P.; Duncan, S.H.; Flint, H.J. Understanding the effects of diet on bacterial metabolism in the large intestine. J. Appl. Microbiol. 2007, 102, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Hudson, J.M.; Cohen, N.D.; Gibbs, P.G.; Thompson, J. Feeding practices associated with colic in horses. J. Am. Vet. Med. Assoc. 2001, 219, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, S.; Julliand, V.; Leblond, A. Risk factors associated with colic in horses. Vet. Res. 2002, 33, 641–652. [Google Scholar] [CrossRef] [Green Version]
- Archer, D.; Proudman, C. Epidemiological clues to preventing colic. Vet. J. 2006, 172, 29–39. [Google Scholar] [CrossRef]
- Frank, N. Equine metabolic syndrome. J. Equine Vet. Sci. 2009, 29, 259–267. [Google Scholar] [CrossRef]
- Wylie, C.E.; Collins, S.N.; Verheyen, K.L.; Newton, J.R. Risk factors for equine laminitis: A case-control study conducted in veterinary-registered horses and ponies in great britain between 2009 and 2011. Vet. J. 2013, 198, 57–69. [Google Scholar] [CrossRef]
- Larsson, A.; Müller, C.E. Owner reported management, feeding and nutrition-related health problems in arabian horses in sweden. Livest. Sci. 2018, 215, 30–40. [Google Scholar] [CrossRef]
- Ellis, A. Biological basis of behaviour in relation to nutrition and feed intake in horses. EAAP Sci. Ser. 2010, 128, 53–74. [Google Scholar]
- Harris, P.A.; Ellis, A.D.; Fradinho, M.J.; Jansson, A.; Julliand, V.; Luthersson, N.; Santos, A.S.; Vervuert, I. Review: Feeding conserved forage to horses: Recent advances and recommendations. Animal 2017, 11, 958–967. [Google Scholar] [CrossRef] [Green Version]
- Särkijärvi, S.; Ahtila, L.; Jokela, P.; Saastamoinen, M. Silage digestibility in equine diets. In Proceedings of the XIIIth International Silage Conference, Auchincruive, Scotland, 11–13 September 2002. [Google Scholar]
- Domingues, J.L. Use of conserved roughage in the horse feeding. Rev. Bras. Zootec. 2009, 38, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Vik, J.; Farstad, M. Hest, hestehold og fôring: Status for hesteholdet i norge. Kommentert frekvensrapport. Bygdeforskning 2012, 2, 1–59. [Google Scholar]
- Vandenput, S.; Istasse, L.; Nicks, B.; Lekeux, P. Airborne dust and aeroallergen concentrations in different sources of feed and bedding for horses. Vet. Q. 1997, 19, 154–158. [Google Scholar] [CrossRef]
- Muhonen, S. Metabolism and Hindgut Ecosystem in Forage Fed Sedentary and Athletic Horses. Ph.D. Thesis, Acta Universitatis Agriculturae Sueciae, Uppsala, Sweden, 30 September 2008. [Google Scholar]
- Müller, C.; Udén, P. Preference of horses for grass conserved as hay, haylage or silage. Anim. Feed Sci. Technol. 2007, 132, 66–78. [Google Scholar] [CrossRef]
- Müller, C.E. Silage and haylage for horses. Grass Forage Sci. 2018, 73, 815–827. [Google Scholar] [CrossRef]
- Muhonen, S.; Julliand, V.; Lindberg, J.; Bertilsson, J.; Jansson, A. Effects on the equine colon ecosystem of grass silage and haylage diets after an abrupt change from hay. Anim. Sci. J. 2009, 87, 2291–2298. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Von Rosen, D.; Udén, P. Effect of forage conservation method on microbial flora and fermentation pattern in forage and in equine colon and faeces. Livest. Sci. 2008, 119, 116–128. [Google Scholar] [CrossRef]
- Guo, Q. Review of feeding silage to horses in northwest China for fifteen years. In Proceedings of the China Animal Science and Veterinary Medicine Convention, Beijing, China, 27 August 2014. (In Chinese). [Google Scholar]
- Carroll, C.; Huntington, P. Body condition scoring and weight estimation of horses. Equine Vet. J. 1988, 20, 41–45. [Google Scholar] [CrossRef]
- Dougal, K.; Harris, P.A.; Girdwood, S.E.; Creevey, C.J.; Curtis, G.C.; Barfoot, C.F.; Argo, C.M.; Newbold, C.J. Changes in the total fecal bacterial population in individual horses maintained on a restricted diet over 6 weeks. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Venable, E.B.; Fenton, K.A.; Braner, V.M.; Reddington, C.E.; Halpin, M.J.; Heitz, S.A.; Francis, J.M.; Gulson, N.A.; Goyer, C.L.; Bland, S.D. Effects of feeding management on the equine cecal microbiota. J. Equine Vet. Sci. 2017, 49, 113–121. [Google Scholar] [CrossRef]
- Biddle, A.S.; Tomb, J.-F.; Fan, Z. Microbiome and blood analyte differences point to community and metabolic signatures in lean and obese horses. Front. Vet. Sci. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, H.L.; Pitta, D.; Indugu, N.; Vecchiarelli, B.; Engiles, J.B.; Southwood, L.L. Characterization of the fecal microbiota of healthy horses. Am. J. Vet. Res. 2018, 79, 811–819. [Google Scholar] [CrossRef]
- Tang, L.; Li, Y.; Srivathsan, A.; Gao, Y.; Li, K.; Hu, D.; Zhang, D. Gut microbiomes of endangered Przewalski’s horse populations in short-and long-term captivity: Implication for species reintroduction based on the soft-release strategy. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one fastq preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. Flash: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Goebel, B.M. Taxonomic note: A place for DNA-DNA reassociation and 16s rrna sequence analysis in the present species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Uparse: Highly accurate otu sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive bayesian classifier for rapid assignment of rrna sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods. 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12. [Google Scholar] [CrossRef] [Green Version]
- Garber, A.; Hastie, P.; Murray, J.-A. Factors influencing equine gut microbiota: Current knowledge. J. Equine Vet. Sci. 2020, 88. [Google Scholar] [CrossRef]
- Hu, D.; Chao, Y.; Li, Y.; Peng, X.; Wang, C.; Wang, Z.; Zhang, D.; Li, K. Effect of gender bias on equine fecal microbiota. J. Equine Vet. Sci. 2021, 97. [Google Scholar] [CrossRef]
- Morrison, P.K.; Newbold, C.J.; Jones, E.; Worgan, H.J.; Grove-White, D.H.; Dugdale, A.H.; Barfoot, C.; Harris, P.A.; McG Argo, C. The equine gastrointestinal microbiome: Impacts of age and obesity. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Dugdale, A.; Curtis, G.; Cripps, P.; Harris, P.; Argo, C.M. Effect of dietary restriction on body condition, composition and welfare of overweight and obese pony mares. Equine Vet. J. 2010, 42, 600–610. [Google Scholar] [CrossRef]
- Grimm, P.; Combes, S.; Pascal, G.; Cauquil, L.; Julliand, V. Dietary composition and yeast/microalgae combination supplementation modulate the microbial ecosystem in the caecum, colon and faeces of horses. Br. J. Nutr. 2020, 123, 372–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julliand, V.; Grimm, P. Horse species symposium: The microbiome of the horse hindgut: History and current knowledge. Anim. Sci. J. 2016, 94, 2262–2274. [Google Scholar] [CrossRef]
- Welp, A.L.; Bomberger, J.M. Bacterial community interactions during chronic respiratory disease. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.A.; Kittelmann, S.; Rogers, C.W.; Gee, E.K.; Bolwell, C.F.; Bermingham, E.N.; Thomas, D.G. Faecal microbiota of forage-fed horses in new zealand and the population dynamics of microbial communities following dietary change. PLoS ONE 2014, 9, e112846. [Google Scholar] [CrossRef] [Green Version]
- Khoruts, A.; Dicksved, J.; Jansson, J.K.; Sadowsky, M.J. Changes in the composition of the human fecal microbiome after bacteriotherapy for recurrent clostridium difficile-associated diarrhea. J. Clin. Gastroenterol. 2010, 44, 354–360. [Google Scholar] [CrossRef]
- Stewart, H.L.; Southwood, L.L.; Indugu, N.; Vecchiarelli, B.; Engiles, J.B.; Pitta, D. Differences in the equine faecal microbiota between horses presenting to a tertiary referral hospital for colic compared with an elective surgical procedure. Equine Vet. J. 2019, 51, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Muck, R.; Shinners, K.J. Conserved forage (silage and hay): Progress and priorities. Int. Grass. Congr. 2001, 19, 753–762. [Google Scholar]
- Paul, K.; Nonoh, J.O.; Mikulski, L.; Brune, A. “Methanoplasmatales,” thermoplasmatales-related archaea in termite guts and other environments, are the seventh order of methanogens. Appl. Environ. Microbiol. 2012, 78, 8245–8253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, B.B.; Jørgensen, H. Effect of dietary fiber on microbial activity and microbial gas production in various regions of the gastrointestinal tract of pigs. Appl. Environ. Microbiol. 1994, 60, 1897–1904. [Google Scholar] [CrossRef] [Green Version]
- Elghandour, M.; Adegbeye, M.J.; Barbabosa-Pilego, A.; Perez, N.R.; Hernández, S.R.; Zaragoza-Bastida, A.; Salem, A.Z.M. Equine contribution in methane emission and its mitigation strategies. J. Equine Vet. Sci. 2019, 72, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Dougal, K.; Harris, P.A.; Edwards, A.; Pachebat, J.A.; Blackmore, T.M.; Worgan, H.J.; Newbold, C.J. A comparison of the microbiome and the metabolome of different regions of the equine hindgut. FEMS Microbiol. Ecol. 2012, 82, 642–652. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.C.; Arroyo, L.G.; Allen-Vercoe, E.; Stämpfli, H.R.; Kim, P.T.; Sturgeon, A.; Weese, J.S. Comparison of the fecal microbiota of healthy horses and horses with colitis by high throughput sequencing of the v3-v5 region of the 16s rrna gene. PLoS ONE 2012, 7, e41484. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Li, B.; Bai, D.; Huang, J.; Shiraigo, W.; Yang, L.; Zhao, Q.; Ren, X.; Wu, J.; Bao, W. Comparison of fecal microbiota of mongolian and thoroughbred horses by high-throughput sequencing of the v4 region of the 16s rrna gene. Asian-Australas. J. Anim. Sci. 2016, 29. [Google Scholar] [CrossRef] [Green Version]
- Bulmer, L.S.; Murray, J.A.; Burns, N.M.; Garber, A.; Wemelsfelder, F.; McEwan, N.R.; Hastie, P.M. High-starch diets alter equine faecal microbiota and increase behavioural reactivity. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Collinet, A.; Grimm, P.; Julliand, S.; Julliand, V. Multidimensional approach for investigating the effects of an antibiotic-probiotic combination on the equine hindgut ecosystem and microbial fibrolysis. Front. Microbiol. 2021, 12, 646294. [Google Scholar] [CrossRef] [PubMed]
- Daly, K.; Proudman, C.J.; Duncan, S.H.; Flint, H.J.; Dyer, J.; Shirazi-Beechey, S.P. Alterations in microbiota and fermentation products in equine large intestine in response to dietary variation and intestinal disease. Br. J. Nutr. 2012, 107, 989–995. [Google Scholar] [CrossRef] [Green Version]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the genetic basis of fibrolytic specialization by lachnospiraceae and ruminococcaceae in diverse gut communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Brulc, J.M.; Antonopoulos, D.A.; Miller, M.E.; Wilson, M.K.; Yannarell, A.C.; Dinsdale, E.A.; Edwards, R.E.; Frank, E.D.; Emerson, J.B.; Wacklin, P.; et al. Gene-centric metagenomics of the fiber-adherent bovine rumen microbiome reveals forage specific glycoside hydrolases. Proc. Natl. Acad. Sci. USA 2009, 106, 1948–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouns, F.; Kettlitz, B.; Arrigoni, E. Resistant starch and “the butyrate revolution”. Trends Food Sci. Technol. 2002, 13, 251–261. [Google Scholar] [CrossRef]
- Xu, Z.; Knight, R. Dietary effects on human gut microbiome diversity. Br. J. Nutr. 2015, 113, S1–S5. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.N.; Amand, A.L.S.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [Green Version]
- De Fombelle, A.; Julliand, V.; Drogoul, C.; Jacotot, E. Feeding and microbial disorders in horses: 1-effects of an abrupt incorporation of two levels of barley in a hay diet on microbial profile and activities. J. Equine Vet. Sci. 2001, 21, 439–445. [Google Scholar] [CrossRef]
- Steelman, S.M.; Chowdhary, B.P.; Dowd, S.; Suchodolski, J.; Janečka, J.E. Pyrosequencing of 16s rrna genes in fecal samples reveals high diversity of hindgut microflora in horses and potential links to chronic laminitis. BMC Vet. Res. 2012, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mshelia, E.S.; Adamu, L.; Wakil, Y.; Turaki, U.A.; Gulani, I.A.; Musa, J. The association between gut microbiome, sex, age and body condition scores of horses in maiduguri and its environs. Microb. Pathog. 2018, 118, 81–86. [Google Scholar] [CrossRef]
- Bolotin, A.; Quinquis, B.; Renault, P.; Sorokin, A.; Ehrlich, S.D.; Kulakauskas, S.; Lapidus, A.; Goltsman, E.; Mazur, M.; Pusch, G.D.; et al. Complete sequence and comparative genome analysis of the dairy bacterium streptococcus thermophilus. Nat. Biotechnol. 2004, 22, 1554–1558. [Google Scholar] [CrossRef] [Green Version]
- Milinovich, G.J.; Klieve, A.V.; Pollitt, C.C.; Trott, D.J. Microbial events in the hindgut during carbohydrate-induced equine laminitis. Vet. Clin. Equine 2010, 26, 79–94. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, K.; Liu, Y.; Li, K.; Hu, D.; Wronski, T. Community composition and diversity of intestinal microbiota in captive and reintroduced Przewalski’s horse (equus ferus przewalskii). Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Stothart, M.; Greuel, R.; Gavriliuc, S.; Henry, A.; Wilson, A.; McLoughlin, P.; Poissant, J. Bacterial drift and dispersal drive microbiome diversity within a population of feral hindgut fermenters. Authorea Prepr. 2020. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pasture Grass | Silage | Hay | |
|---|---|---|---|
| DM 1 (%NM 2) | 15.53 | 25.78 | 91.50 |
| CP 3 (%DM) | 13.11 | 8.54 | 9.36 |
| CF 4 (%DM) | 0.98 | 1.34 | 1.82 |
| Ash (%DM) | 11.24 | 9.91 | 5.04 |
| NDF 5 (%DM) | 47.56 | 66.62 | 59.89 |
| ADF 6 (%DM) | 29.02 | 46.94 | 34.71 |
| Ca 7 (%DM) | 0.57 | 0.28 | 0.16 |
| P 8 (%DM) | 0.13 | 0.22 | 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Wang, X.; Deng, L.; Chen, S.; Zhu, C.; Li, J. Effects of Pasture Grass, Silage, and Hay Diet on Equine Fecal Microbiota. Animals 2021, 11, 1330. https://doi.org/10.3390/ani11051330
Zhu Y, Wang X, Deng L, Chen S, Zhu C, Li J. Effects of Pasture Grass, Silage, and Hay Diet on Equine Fecal Microbiota. Animals. 2021; 11(5):1330. https://doi.org/10.3390/ani11051330
Chicago/Turabian StyleZhu, Yiping, Xuefan Wang, Liang Deng, Shulei Chen, Chunyan Zhu, and Jing Li. 2021. "Effects of Pasture Grass, Silage, and Hay Diet on Equine Fecal Microbiota" Animals 11, no. 5: 1330. https://doi.org/10.3390/ani11051330
APA StyleZhu, Y., Wang, X., Deng, L., Chen, S., Zhu, C., & Li, J. (2021). Effects of Pasture Grass, Silage, and Hay Diet on Equine Fecal Microbiota. Animals, 11(5), 1330. https://doi.org/10.3390/ani11051330