
Influence of Environmental Temperature and Hormonal Stimulation on the In Vitro Sperm Maturation in Sterlet Acipenser ruthenus in Advance of the Spawning Season

 , , , and
, , , and 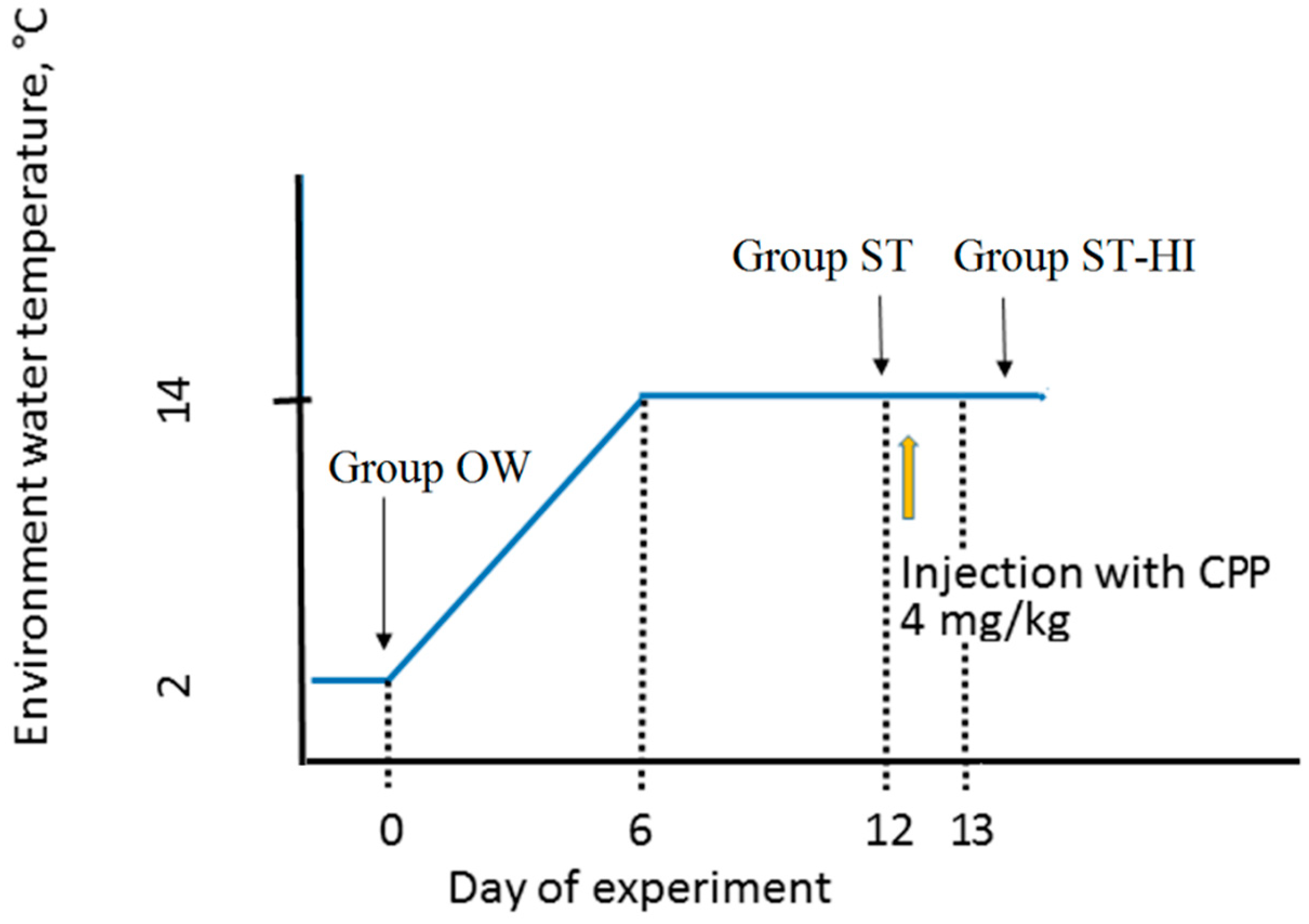{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Fish Rearing Conditions and Experimental Groups
2.3. Collection of Sperm, Blood and Testis Samples
2.4. Histological Analysis
2.5. Plasma Sex Steroid Hormone Evaluation
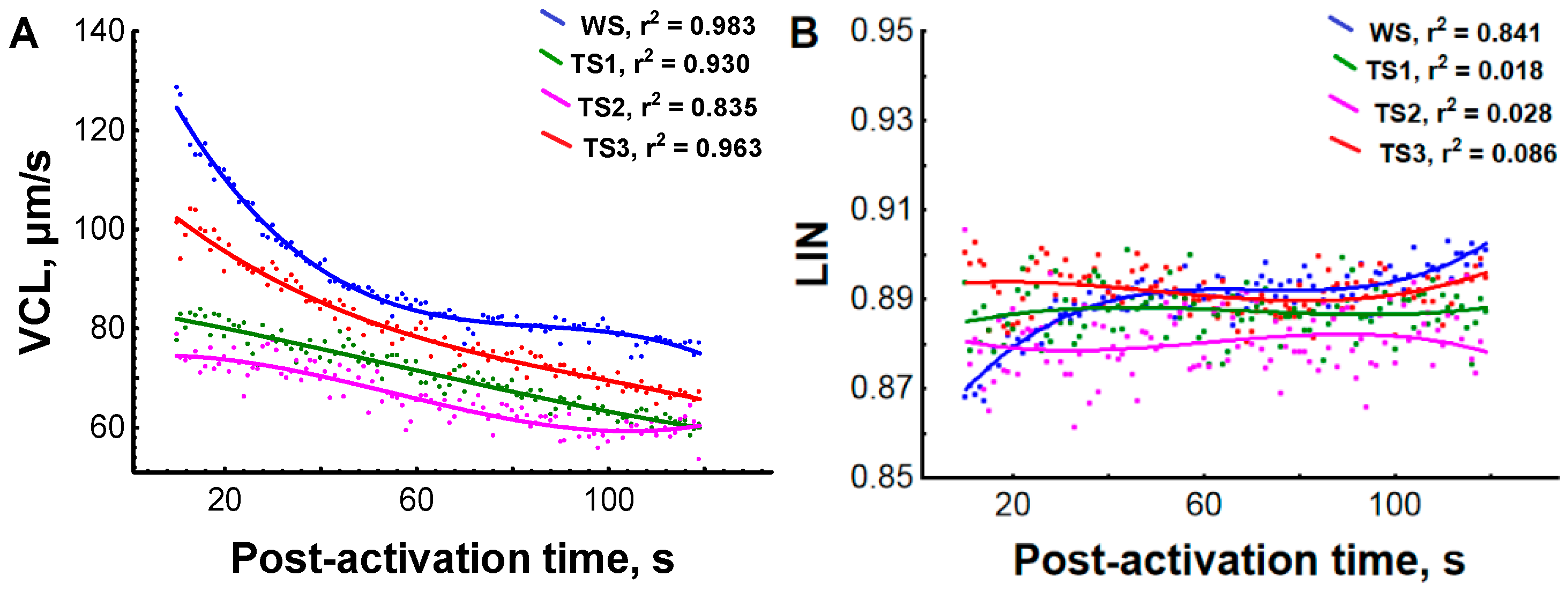
2.6. In Vitro Maturation of Testicular Spermatozoa
2.7. Sperm Motility Analysis
2.8. Statistics
3. Results and Discussion
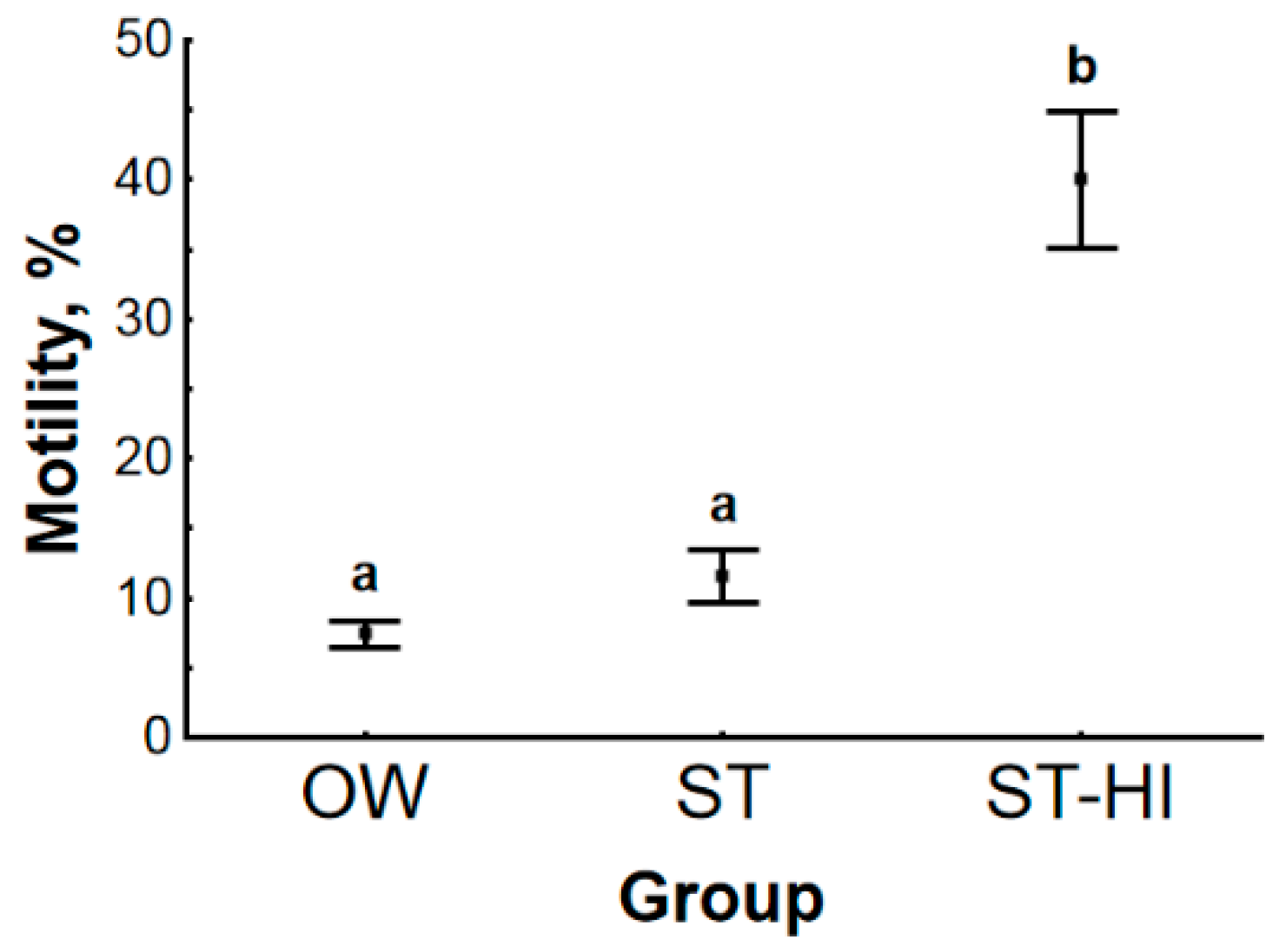
3.1. Effect of Environmental Temperature and Hormonal Stimulation on Sperm Motility Parameters
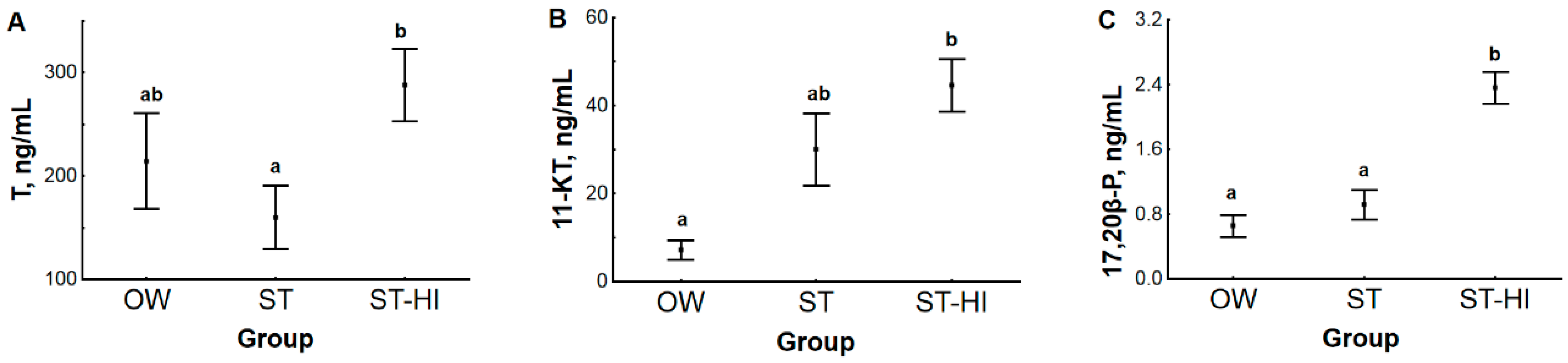
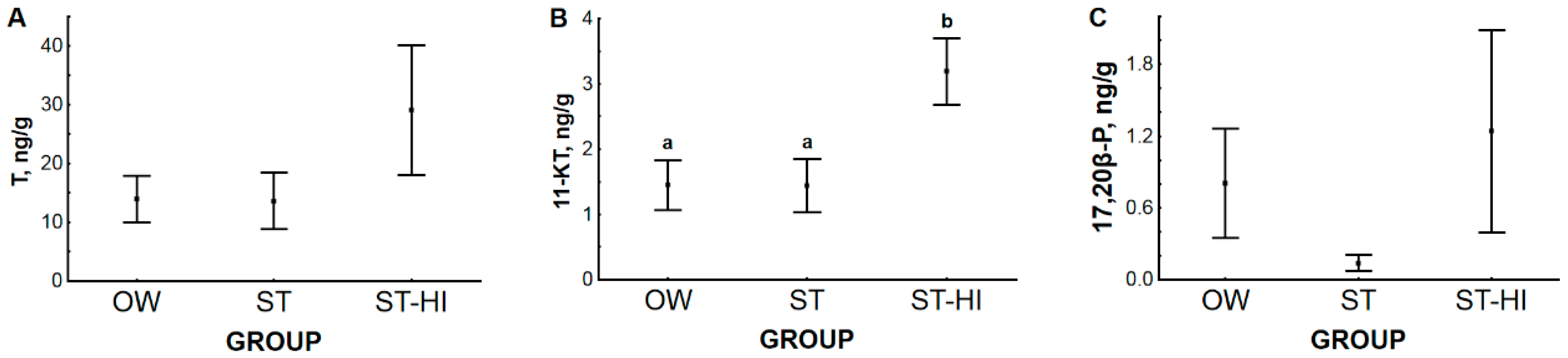
3.2. Effect of Environmental Temperature and Hormonal Stimulation on Content of Sex Steroid Hormones in Blood Plasma and Testes
3.2.1. Hormone Content in Blood Plasma and Testes
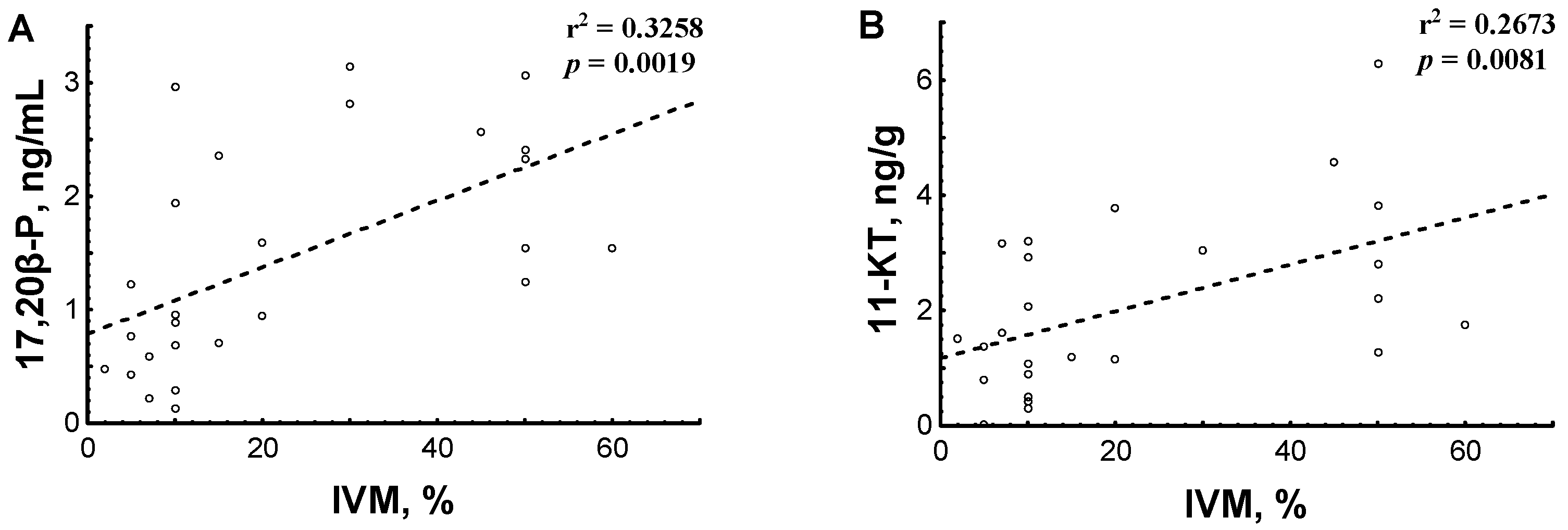
3.2.2. Hormone Content in Relation to Testicular Sperm IVM
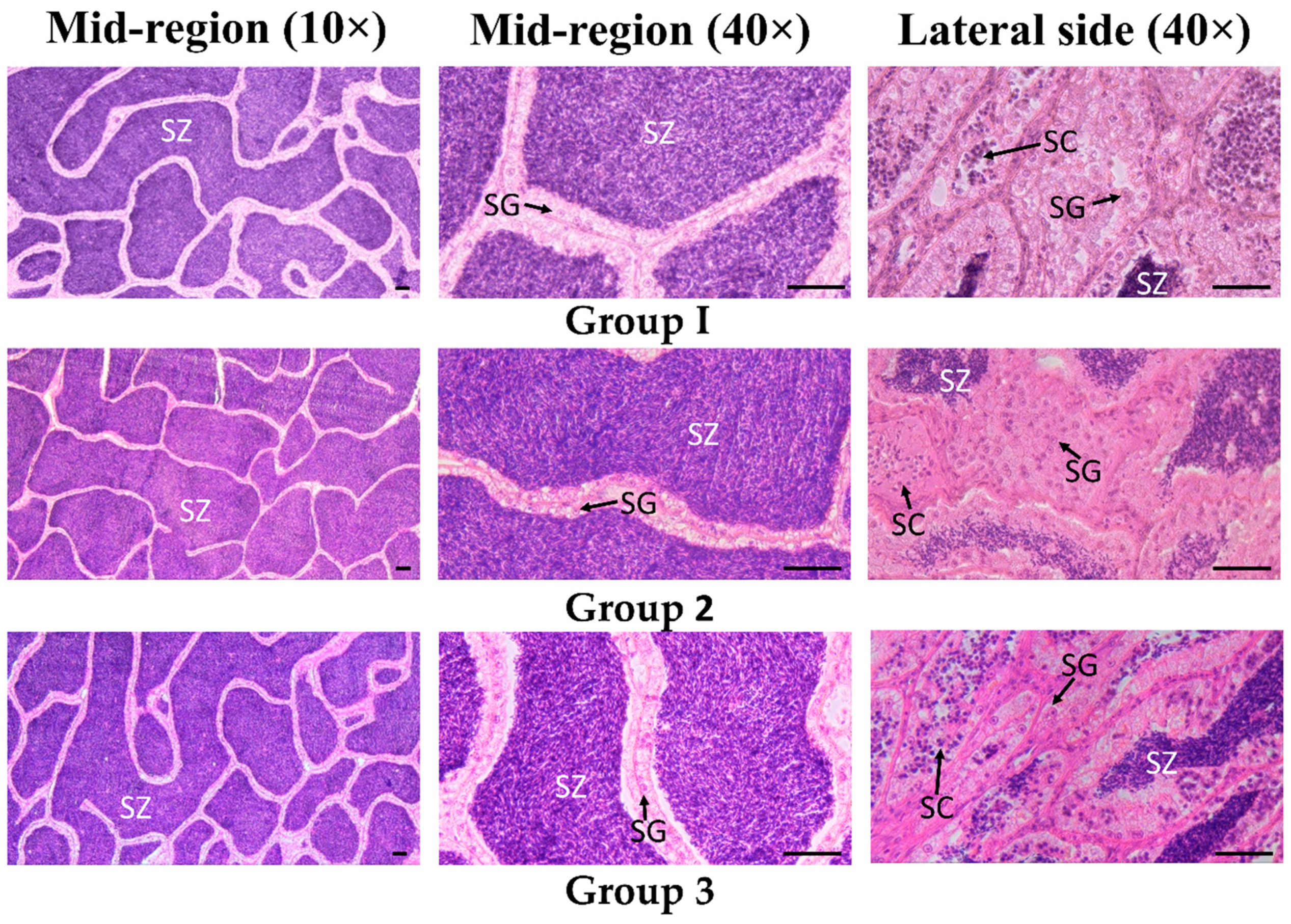
3.3. Testes Histological Identification
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Betancur-R, R.; Wiley, E.; Arratia, G.; Acero, A.; Bailly, N.; Lecointre, G.; Orti, G. Phylogenetic classification of bony fishes. BMC Evol. Biol. 2017, 17, 162. [Google Scholar] [CrossRef] [Green Version]
- Dzyuba, V.; Shelton, W.L.; Kholodnyy, V.; Boryshpolets, S.; Cosson, J.; Dzyuba, B. Fish sperm biology in relation to urogenital system structure. Theriogenology 2019, 132, 153–163. [Google Scholar] [CrossRef]
- Dzyuba, B.; Cosson, J.; Boryshpolets, S.; Bondarenko, O.; Dzyuba, V.; Prokopchuk, G.; Gazo, I.; Rodina, M.; Linhart, O. In vitro sperm maturation in sterlet, Acipenser ruthenus. Reprod. Biol. 2014, 14, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Dzyuba, B.; Boryshpolets, S.; Cosson, J.; Dzyuba, V.; Fedorov, P.; Saito, T.; Psenicka, M.; Linhart, O.; Rodina, M. Motility and fertilization ability of sterlet Acipenser ruthenus testicular sperm after cryopreservation. Cryobiology 2014, 69, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Bronzi, P.; Rosenthal, H. Present and future sturgeon and caviar production and marketing: A global market overview. J. Appl. Ichthyol. 2014, 30, 1536–1546. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Rodina, M.; Gela, D.; Linhart, O. Sperm biology and control of reproduction in sturgeon: (I) testicular development, sperm maturation and seminal plasma characteristics. Rev. Fish Biol. Fish. 2012, 22, 695–717. [Google Scholar] [CrossRef]
- Dettlaff, T.A.; Ginsburg, A.S.; Schmalhausen, O.I. Sturgeon Fishes: Developmental Biology and Aquaculture; Springer-Verlag: Berlin/Heidelberg, Germany, 1993. [Google Scholar]
- Chebanov, M.S.; Galich, E.V. Sturgeon Hatchery Manual; Food and Agriculture Organization of United Nations: Ankara, Turkey, 2013.
- Dzyuba, V.; Dzyuba, B.; Cosson, J.; Boryshpolets, S.; Yamaner, G.; Kholodniy, V.; Rodina, M. The antioxidant system of sterlet seminal fluid in testes and Wolffian ducts. Fish Physiol. Biochem. 2014, 40, 1731–1739. [Google Scholar] [CrossRef]
- Amiri, B.M.; Maebayashi, M.; Adachi, S.; Yamauchi, K. Testicular development and serum sex steroid profiles during the annual sexual cycle of the male sturgeon hybrid the bester. J. Fish Biol. 1996, 48, 1039–1050. [Google Scholar] [CrossRef]
- Rodríguez, L.; Begtashi, I.; Zanuy, S.; Carrillo, M. Development and validation of an enzyme immunoassay for testosterone: Effects of photoperiod on plasma testosterone levels and gonadal development in male sea bass (Dicentrarchus labrax, L.) at puberty. Fish Physiol. Biochem. 2000, 23, 141–150. [Google Scholar] [CrossRef]
- Cuisset, B.; Pradelles, P.; Kime, D.E.; Kühn, E.R.; Babin, P.; Davail, S.; Le Menn, F. Enzyme immunoassay for 11-ketotestosterone using acetylcholinesterase as laberl: Application to the measurement of 11-ketotestosterone in plasma of Siberian sturgeon. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1994, 108, 229–241. [Google Scholar] [CrossRef]
- Nash, J.P.; Cuisset, B.D.; Bhattacharyya, S.; Suter, H.C.; Le Menn, F.; Kime, D.E. An enzyme linked immunosorbant assay (ELISA) for testosterone, estradiol, and 17,20β-dihydroxy-4-pregenen-3-one using acetylcholinesterase as tracer: Application to measurement of diel patterns in rainbow trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 2000, 22, 355–363. [Google Scholar] [CrossRef]
- Wilson-Leedy, J.G.; Ingermann, R.L. Development of a novel CASA system based on open source software for characterization of zebrafish sperm motility parameters. Theriogenology 2007, 67, 661–672. [Google Scholar] [CrossRef]
- Purchase, C.F.; Earle, P.T. Modifications to the IMAGEJ computer assisted sperm analysis plugin greatly improve efficiency and fundamentally alter the scope of attainable data. J. Appl. Ichthyol. 2012, 28, 1013–1016. [Google Scholar] [CrossRef]
- McDonald, J.H. Handbook of Biological Statistics, 2nd ed.; Sparky House Publishing: Baltimore, MD, USA, 2009. [Google Scholar]
- Sieczyński, P.; Glogowski, J.; Cejko, B.; Grygoruk, C. Characteristics of Siberian sturgeon and sterlet sperm motility parameters compared using CASA. Arch. Pol. Fish. 2012, 20, 137–143. [Google Scholar] [CrossRef]
- Boryshpolets, S.; Kholodnyy, V.; Cosson, J.; Dzyuba, B. Fish sperm motility analysis: The central role of the flagellum. Reprod. Fertil. Dev. 2018, 30, 833–841. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Duncan, N.J.; Asturiano, J.F. Hormonal manipulations for the enhancement of sperm production in cultured fish and evaluation of sperm quality. Aquaculture 2017, 472, 21–44. [Google Scholar] [CrossRef]
- Caille, N.; Rodina, M.; Kocour, M.; Gela, D.; Flajšhans, M.; Linhart, O. Quantity, motility and fertility of tench Tinca tinca (L.) sperm in relation to LHRH analogue and carp pituitary treatments. Aquac. Int. 2006, 14, 75–87. [Google Scholar] [CrossRef]
- Cejko, B.I.; Krejszeff, S.; Żarski, D.; Judycka, S.; Targońska, K.; Kucharczyk, D. Effect of carp pituitary homogenate (CPH) and sGnRHa (Ovaprim) on northern pike (Esox lucius) spermiation stimulation and its effect on quantity and quality of sperm. Anim. Reprod. Sci. 2018, 193, 217–225. [Google Scholar] [CrossRef]
- Cejko, B.I.; Żarski, D.; Krejszeff, S.; Kucharczyk, D.; Kowalski, R.K. Effect of hormonal stimulation on milt volume, number of sperm, and sperm motility in the crucian carp, Carassius carassius (L.). Isr. J. Aquacult. Bamid. 2013, 65, 1–7. [Google Scholar]
- Fakriadis, I.; Zanatta, E.M.; Fleck, R.P.D.S.; Sena Mateo, D.L.; Papadaki, M.; Mylonas, C.C. Endocrine regulation of long-term enhancement of spermiation in meagre (Argyrosomus regius) with GnRHa controlled-delivery systems. Gen. Comp. Endocrinol. 2020, 297, 113549. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Hatef, A.; Mylonas, C.C.; Gela, D.; Papadaki, M.; Rodina, M.; Kašpar, V.; Pšenička, M.; Podhorec, P.; Linhart, O. Sperm characteristics and androgens in Acipenser ruthenus after induction of spermiation by carp pituitary extract or GnRHa implants. Fish Physiol. Biochem. 2012, 38, 1655–1666. [Google Scholar] [CrossRef]
- Linhart, O.; Mims, S.D.; Gomelsky, B.; Hiott, A.E.; Shelton, W.L.; Cosson, J.; Rodina, M.; Gela, D. Spermiation of paddlefish (Polyodon spathula, Acipenseriformes) stimulated with injection of LHRH analogue and carp pituitary powder. Aquat. Living Resour. 2000, 13, 455–460. [Google Scholar] [CrossRef]
- Billard, R. Biology and control of reproduction of sturgeons in fish farm. Iran. J. Fish. Sci. 2000, 2, 1–20. [Google Scholar]
- Saad, A.; Billard, R. Spermatozoa production and volume of semen collected after hormonal stimulation in the carp, Cyprinus carpio. Aquaculture 1987, 65, 67–77. [Google Scholar] [CrossRef]
- Lofts, B. Testicular Function. In Hormones and Reproduction in Fishes, Amphibians, and Reptiles; Norris, D.O., Jones, R.E., Eds.; Plenum Press: New York, NY, USA, 1987; pp. 283–325. [Google Scholar] [CrossRef]
- Pham, H.Q.; Nguyen, A.T.; Kjørsvik, E.; Nguyen, M.D.; Arukwe, A. Seasonal reproductive cycle of Waigieu seaperch (Psammoperca waigiensis). Aquacult. Res. 2012, 43, 815–830. [Google Scholar] [CrossRef]
- Sisneros, J.A.; Forlano, P.M.; Knapp, R.; Bass, A.H. Seasonal variation of steroid hormone levels in an intertidal-nesting fish, the vocal plainfin midshipman. Gen. Comp. Endocrinol. 2004, 136, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.W.; de França, L.R.; Lareyre, J.-J.; LeGac, F.; Chiarini-Garcia, H.; Nobrega, R.H.; Miura, T. Spermatogenesis in fish. Gen. Comp. Endocrinol. 2010, 165, 390–411. [Google Scholar] [CrossRef] [PubMed]
- Barannikova, I.A.; Bayunova, L.V.; Semenkova, T.B. Serum levels of testosterone, 11-ketotestosterone and oestradiol-17β in three species of sturgeon during gonadal development and final maturation induced by hormonal treatment. J. Fish Biol. 2004, 64, 1330–1338. [Google Scholar] [CrossRef]
- Du, H.; Zhang, X.; Xiaoqian, L.; Zhang, S.; Luo, J.; Liu, Z.; Qiao, X.; Kynard, B.; Wei, Q. Gender and gonadal maturity stage identification of captive Chinese sturgeon, Acipenser sinensis, using ultrasound imagery and sex steroids. Gen. Comp. Endocrinol. 2017, 245, 36–43. [Google Scholar] [CrossRef]
- Webb, M.A.H.; Feist, G.W.; Foster, E.P.; Schreck, C.B.; Fitzpatrick, M.S. Potential classification of sex and stage of gonadal maturity of wild white sturgeon using blood plasma indicators. Trans. Am. Fish. Soc. 2002, 131, 132–142. [Google Scholar] [CrossRef]
- Wildhaber, M.L.; Papoulias, D.M.; DeLonay, A.J.; Tillitt, D.E.; Bryan, J.L.; Annis, M.L. Physical and hormonal examination of Missouri River shovelnose sturgeon reproductive stage: A reference guide. J. Appl. Ichthyol. 2007, 23, 382–401. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, H.; Dong, Y.; Tian, Z.; Dong, T.; Hu, H.; Niu, C. Dimorphic expression of sex-related genes in different gonadal development stages of sterlet, Acipenser ruthenus, a primitive fish species. Fish Physiol. Biochem. 2017, 43, 1557–1569. [Google Scholar] [CrossRef]
- Vizziano, D.; Barrios, F.; Astigarraga, I.; Breton, B.; Williot, P. Unusual conditions for Siberian sturgeon (Acipenser baerii Brandt) spawning. J. Appl. Ichthyol. 2006, 22, 325–330. [Google Scholar] [CrossRef]
- Semenkova, T.; Barannikova, I.; Kime, D.E.; McAllister, B.G.; Bayunova, L.; Dyubin, V.; Kolmakov, N. Sex steroid profiles in female and male stellate sturgeon (Acipenser stellatus Pallas) during final maturation induced by hormonal treatment. J. Appl. Ichthyol. 2002, 18, 375–381. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Scott, A.P.; Vermeirssen, E.L.M.; Zohar, Y. Changes in plasma gonadotropin II and sex steroid hormones, and sperm production of striped bass after treatment with controlled-release gonadotropin-releasing hormone agonist-delivery systems. Biol. Reprod. 1997, 57, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Fostier, A.; Jalabert, B.; Billard, R.; Breton, B.; Zohar, Y. The gonadal steroids. In Fish Physiology; Hoar, W.S., Randall, D.J., Donaldson, E.M., Eds.; Academic Press: Orlando, FL, USA, 1983; Volume 9A, pp. 277–372. [Google Scholar]
- Arai, R.; Tamaoki, B.-I. Steroid biosynthesis in vitro by testes of rainbow trout. Salmo gairdneri. Gen. Comp. Endocrinol. 1967, 8, 305–313. [Google Scholar] [CrossRef]
- Bukovskaya, O.; Lambert, J.G.D.; Kime, D.E. In vitro steroidogenesis by gonads of the Russian sturgeon, Acipenser gueldenstaedti Brandt. Fish Physiol. Biochem. 1997, 16, 345–353. [Google Scholar] [CrossRef]
- Colombo, L.; Belvedere, P.C.; Arcarese, G. Gonadal steroidogenesis and gametogenesis in teleost fishes—a study on the sea bass, Dicentrarchus labrax L. Ital. J. Zool. 1978, 45, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Idler, D.R.; Macnab, H.C. The biosynthesis of 11-ketotestosterone and 11β-hydroxytestosterone by Atlantic salmon tissues in vitro. Can. J. Biochem. 1967, 45, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Asturiano, J.F.; Sorbera, L.A.; Ramos, J.; Kime, D.E.; Carrillo, M.; Zanuy, S. Group-synchronous ovarian development, spawning and spermiation in the European sea bass (Dicentrarchus labrax L.) could be regulated by shifts in gonadal steroidogenesis. Sci. Mar. 2002, 66, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.; Pang, Y.; Zhu, Y.; Detweiler, C.; Doughty, K. Multiple rapid progestin actions and progestin membrane receptor subtypes. Steroids 2004, 69, 567–573. [Google Scholar] [CrossRef]
- Miura, T.; Miura, C.I. Molecular control mechanisms of fish spermatogenesis. Fish Physiol. Biochem. 2003, 28, 181–186. [Google Scholar] [CrossRef]
- Scott, A.P.; Sumpter, J.P.; Stacey, N. The role of the maturation-inducing steroid, 17,20β-dihydroxypregn-4-en-3-one, in male fishes: A review. J. Fish Biol. 2010, 76, 183–224. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.W.; Miura, T. Spermatogenesis and its endocrine regulation. Fish Physiol. Biochem. 2002, 26, 43–56. [Google Scholar] [CrossRef]
- Miura, T.; Miura, C. Japanese eel: A model for analysis of spermatogenesis. Zool. Sci. 2001, 18, 1055–1063. [Google Scholar] [CrossRef]
- Morisawa, S.; Morisawa, M. Induction of potential for sperm motility by bicarbonate and pH in rainbow trout and chum salmon. J. Exp. Biol. 1988, 136, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Ikeda, K.; Izawa, T. Increases in concentrations of potassium and bicarbonate ions promote acquisition of motility in vitro by Japanese eel spermatozoa. J. Exp. Zool. 1997, 277, 171–180. [Google Scholar] [CrossRef]
- Bayunova, L.; Canario, A.V.M.; Semenkova, T.; Dyubin, V.; Sverdlova, O.; Trenkler, I.; Barannikova, I. Sex steroids and cortisol levels in the blood of stellate sturgeon (Acipenser stellatus Pallas) during final maturation induced by LH-RH-analogue. J. Appl. Ichthyol. 2006, 22, 334–339. [Google Scholar] [CrossRef]
- Prokopchuk, G.; Dzyuba, B.; Rodina, M.; Cosson, J. Control of sturgeon sperm motility: Antagonism between K+ ions concentration and osmolality. Anim. Reprod. Sci. 2016, 164, 82–89. [Google Scholar] [CrossRef]
- Cosson, J.; Linhart, O. Paddlefish, Polyodon spathula, spermatozoa: Effects of potassium and pH on motility. Folia Zool. 1996, 45, 361–370. [Google Scholar]
- Linhart, O.; Cosson, J.; Mims, S.D.; Shelton, W.L.; Rodina, M. Effects of ions on the motility of fresh and demembranated paddlefish (Polyodon spathula) spermatozoa. Reproduction 2002, 124, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, O.; Dzyuba, B.; Rodina, M.; Cosson, J. Role of Ca2+ in the IVM of spermatozoa from the sterlet Acipenser ruthenus. Reprod. Fertil. Dev. 2016, 29, 1319–1328. [Google Scholar] [CrossRef]
- Linhart, O.; Mims, S.D.; Gomelsky, B.; Hiott, A.E.; Shelton, W.L.; Cosson, J.; Rodina, M.; Gela, D.; Bastl, J. Ionic composition and osmolality of paddlefish (Polyodon spathula, Acipenseriformes) seminal fluid. Aquac. Int. 2003, 11, 357–368. [Google Scholar] [CrossRef]
- Flynn, S.R.; Benfey, T.J. Sex differentiation and aspects of gametogenesis in shortnose sturgeon Acipenser brevirostrum Lesueur. J. Fish Biol. 2007, 70, 1027–1044. [Google Scholar] [CrossRef]
- Barannikova, I.A.; Bayunova, L.V.; Semenkova, T.B.; Trenkler, I.V. Changes in the physiological state of hiemal form of the Russian sturgeon Acipenser gueldenstaedtii in the Volga after holding it and hormonal impacts. J. Ichthyol. 2008, 48, 402–407. [Google Scholar] [CrossRef]
- Divers, S.J.; Boone, S.S.; Hoover, J.J.; Boysen, K.A.; Killgore, K.J.; Murphy, C.E.; George, S.G.; Camus, A.C. Field endoscopy for identifying gender, reproductive stage and gonadal anomalies in free-ranging sturgeon (Scaphirhynchus) from the lower Mississippi River. J. Appl. Ichthyol. 2009, 25, 68–74. [Google Scholar] [CrossRef]
- Doroshov, S.I.; Moberg, G.P.; Van Eenennaam, J.P. Observations on the reproductive cycle of cultures white sturgeon, Acipenser transmontanus. Environ. Biol. Fishes 1997, 48, 265–278. [Google Scholar] [CrossRef]
- Matsche, M.A.; Rosemary, K.M.; Brundage, H.M., III; O’Herron, J.C., II. Reproductive demographics, intersex, and altered hormone levels in shortnose sturgeon, Acipenser brevirostrum, from Delaware River, USA. J. Appl. Ichthyol. 2013, 29, 299–309. [Google Scholar] [CrossRef]
- Petochi, B.T.; Di Marco, P.; Donadelli, V.; Longobardi, A.; Corsalini, I.; Bertotto, D.; Finoia, M.G.; Marino, G. Sex and reproductive stage identification of sturgeon hybrids (Acipenser naccarii × Acipenser baerii) using different tools: Ultrasounds, histology and sex steroids. J. Appl. Ichthyol. 2011, 27, 637–642. [Google Scholar] [CrossRef]
- Pšenička, M.; Saito, T.; Linhartová, Z.; Gazo, I. Isolation and transplantation of sturgeon early-stage germ cells. Theriogenology 2015, 83, 1085–1092. [Google Scholar] [CrossRef] [Green Version]
- Golpour, A.; Broquard, C.; Milla, S.; Dadras, H.; Baloch, A.R.; Saito, T.; Pšenička, M. Determination of annual reproductive cycle in male sterlet, Acipenser ruthenus using histology and ultrasound imaging. Fish Physiol. Biochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Van Eenennaam, J.P.; Doroshov, S.I. Effects of age and body size on gonadal development of Atlantic sturgeon. J. Fish Biol. 1998, 53, 624–637. [Google Scholar] [CrossRef]
- Yarmohammadi, M.; Pourkazemi, M.; Kazemi, R.; Sadati, M.A.Y.; Hallajian, A.; Saber, M.H. Sex steroid level and sexual dimorphism expression of genes in gonads of the great sturgeon Huso huso Linneaus, 1758 during maturity developmental stages. Aquacult. Res. 2017, 48, 1413–1429. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dzyuba, V.; Cosson, J.; Papadaki, M.; Mylonas, C.C.; Steinbach, C.; Rodina, M.; Tučkova, V.; Linhart, O.; Shelton, W.L.; Gela, D.; et al. Influence of Environmental Temperature and Hormonal Stimulation on the In Vitro Sperm Maturation in Sterlet Acipenser ruthenus in Advance of the Spawning Season. Animals 2021, 11, 1417. https://doi.org/10.3390/ani11051417
Dzyuba V, Cosson J, Papadaki M, Mylonas CC, Steinbach C, Rodina M, Tučkova V, Linhart O, Shelton WL, Gela D, et al. Influence of Environmental Temperature and Hormonal Stimulation on the In Vitro Sperm Maturation in Sterlet Acipenser ruthenus in Advance of the Spawning Season. Animals. 2021; 11(5):1417. https://doi.org/10.3390/ani11051417
Chicago/Turabian StyleDzyuba, Viktoriya, Jacky Cosson, Maria Papadaki, Constantinos C. Mylonas, Christoph Steinbach, Marek Rodina, Vladimira Tučkova, Otomar Linhart, William L. Shelton, David Gela, and et al. 2021. "Influence of Environmental Temperature and Hormonal Stimulation on the In Vitro Sperm Maturation in Sterlet Acipenser ruthenus in Advance of the Spawning Season" Animals 11, no. 5: 1417. https://doi.org/10.3390/ani11051417