1. Introduction
Milk is recognized as being highly nutritious, with global yearly consumption estimated at 113 kg of raw milk per capita in 2018 [
1]. However, there is increasing concern over the environmental impact of food production and, consequently, dairy supply chains worldwide have committed to lowering greenhouse (GHG) emissions to meet the commitments of the Paris Agreement. Many milk processing companies worldwide (e.g., Fonterra, New Zealand; Nestlé, Switzerland; Arlo Foods, Denmark; First Milk, United Kingdom; Emmi AG, Switzerland) have pledged to reduce GHG emissions at the farm level. In the United States, the Net Zero Initiative by a group of dairy industry stakeholders, including farmers, cooperatives, processing companies, and industry organizations, is helping dairy farms decrease GHG emissions by promoting optimized feed production, enhanced feed efficiency, decreased enteric methane (CH
4) emissions, improved manure and nutrient management, and energy generation [
2].
Gerber et al. [
3] reported a global average GHG intensity of 2.4 kg carbon dioxide equivalent (CO
2e)/kg of fat and protein corrected milk (FPCM), with CH
4, nitrous oxide (N
2O), and carbon dioxide (CO
2) from animals, manure, feed production, and fuel use included in the life cycle assessment from “cradle to retail”. The GHG intensity of milk varied greatly among different regions of the world, ranging from 1 to 2 kg CO
2e/kg FPCM for the industrialized regions of the world, to between 3 and 5 kg CO
2e/kg FPCM for South Asian, West Asian, North African, and Central and South American countries, with a high of 7.5 kg CO
2e/kg FPCM for sub-Saharan Africa [
3]. More than 70% of the GHG emissions from milk production occurs prior to the farm-gate [
4], with enteric CH
4 comprising 35% to 55% [
4,
5,
6] of farm emissions. Therefore, abatement of farm-based emissions significantly reduces the GHG emission intensity of milk.
Diet formulation shows promise in terms of mitigating enteric CH
4 emissions [
7,
8]. However, a change in ingredient composition of diets to lower CH
4 emissions may also alter the GHG emissions from feed produced on- and off-farm, as well as emissions from transportation of imported feed to the farm. As the GHG emissions associated with feed production and transportation comprise almost 30% of GHG intensity of milk production to the farm-gate in industrialized countries [
4], it is important to understand the implications for GHG intensity of milk when recommending changes in diet composition to decrease enteric CH
4 production.
Soybean meal (SBM), which contains 50 to 55% crude protein (CP) on a dry matter (DM) basis, is the most widely used protein source in livestock diets globally. Canada produces approximately 1.5 million tonnes of soybean meal per year, which is used as livestock feed [
9]. Additionally, Canada is a major producer of canola seed, with 8.4 million ha planted in 2020 (55% in Saskatchewan (SK)), resulting in 5.8 million tonnes of meal for livestock feeding [
10]. Canola meal (CM), which contains 40.9% ± 2.8% CP (DM basis) [
11]), can be a cost-effective high quality protein source for dairy diets. Dairy diets are supplemented with protein to meet the nitrogen requirement of the rumen microbes and the amino acid requirements of the cow. In a recent study, Benchaar et al. [
12] concluded that replacing solvent-extracted SBM with solvent-extracted CM in iso-nitrogenous diets of lactating dairy cows decreased enteric CH
4 emissions expressed relative to gross energy (GE) intake (i.e., Y
m value) by 13%.
Changes in diet formulation may alter milk production of cows, and because GHG intensity is calculated as the ratio between emissions and product (i.e., meat and milk), an increase in animal performance could lower the GHG intensity of milk [
13]. At low dietary inclusion rates (<10% of dietary DM) in iso-nitrogenous diets, no difference in dairy cow performance was observed between SBM and CM [
14,
15,
16]. However, greater CM incorporation may increase milk production; Benchaar et al. [
12] reported that milk production increased linearly as CM proportion in the diet increased up to 24% of DM.
While CM is routinely used as a protein source in dairy diets to promote milk production, prior to recommending its use to decrease enteric CH
4 production, it is important to conduct a farm-scale life cycle assessment that considers all emission sources to determine the net effects on the GHG intensity of milk. Mathematical models that estimate whole farm GHG emissions can be useful tools for assessing changes in farm management practices. The whole-farm model and software tool Holos, developed by Agriculture and Agri-Food Canada, has been used previously to estimate GHG emissions from Canadian dairy [
5,
17] and beef [
18] farms.
The objective of the present study was to determine whether feeding CM rather than SBM to lactating dairy cows should be recommended as a GHG mitigation strategy. The study focused on Canada because dietary mitigation strategies are dependent upon local conditions, although the concepts and methodology used are universally applicable for evaluating GHG mitigating strategies. We compared the use of SBM and CM as protein supplements in the diets fed to lactating cows on the GHG emission intensity of milk production for a representative dairy farm in eastern (Quebec, QC) and western (Alberta, AB) Canada. The net impact of the CH
4-mitigating property of CM compared with SBM as reported by Benchaar et al. [
12] was examined. We also considered whether the place of origin of the SBM and CM within Canada affects the GHG emission intensity of milk.
4. Discussion
Benchaar et al. [
12] reported that feeding an isonitrogenous diet supplemented with CM (23.7% of DM) rather than SBM (17.0% of DM) to lactating dairy cows decreased the Y
m by 13%. The CH
4-mitigating effect of CM was accompanied by a 7% increase in DMI and a 5% increase in FPCM. While Oba et al. [
19] did not measure enteric CH
4 production, they reported no difference in milk production and DMI for lactating cows fed diets containing CM (13.0% of DM) or SBM (10.2% of DM). Using these data to model the GHG intensity of meat and milk produced by typical dairy farms in QC and AB showed that whether CM should be recommended as a GHG mitigation strategy depended upon two main factors: firstly, the emission factor of the CM relative to that of SBM, which varied with where the crops were grown and the meals produced; and, secondly, the CH
4-mitigating property of CM.
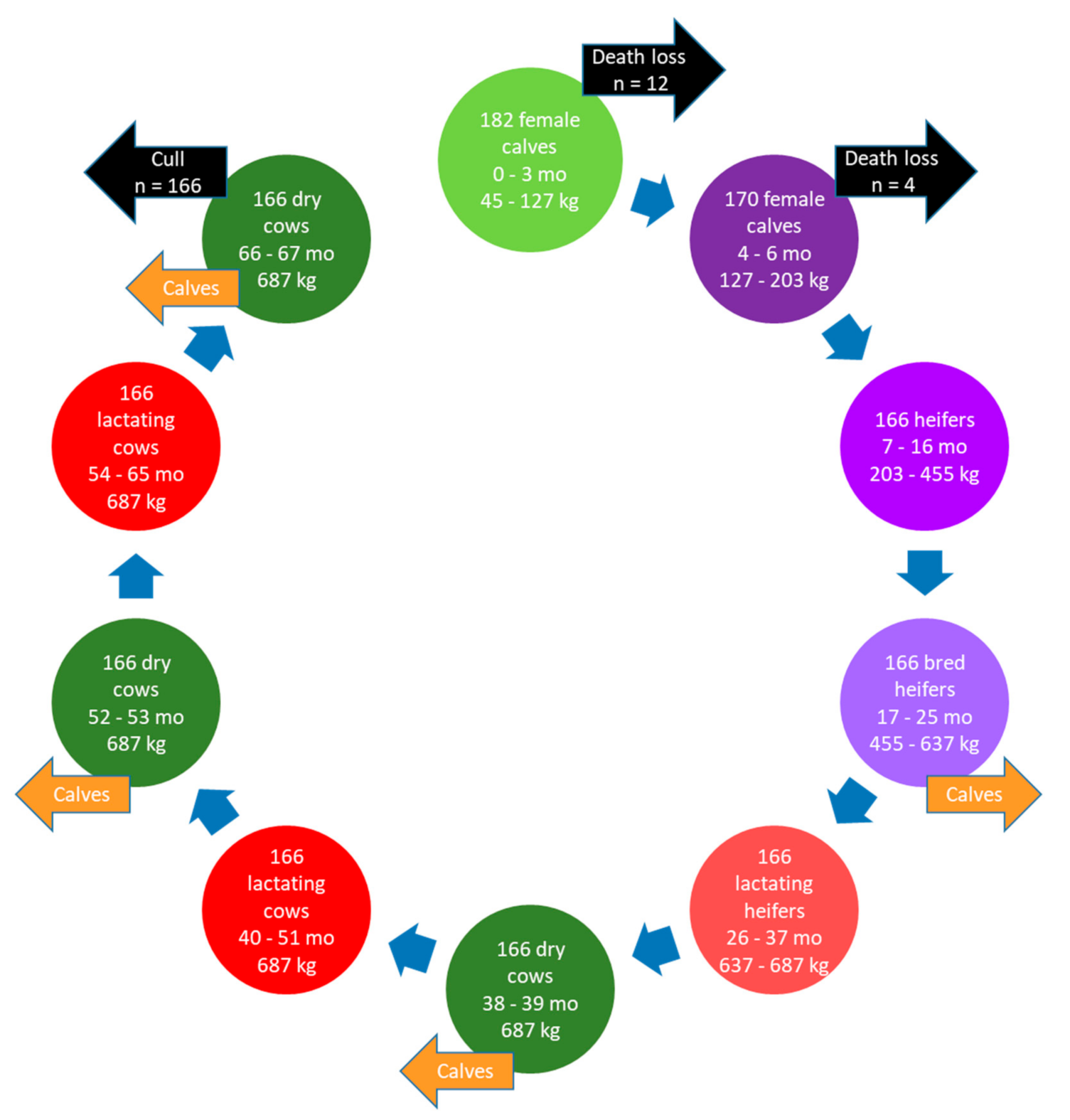
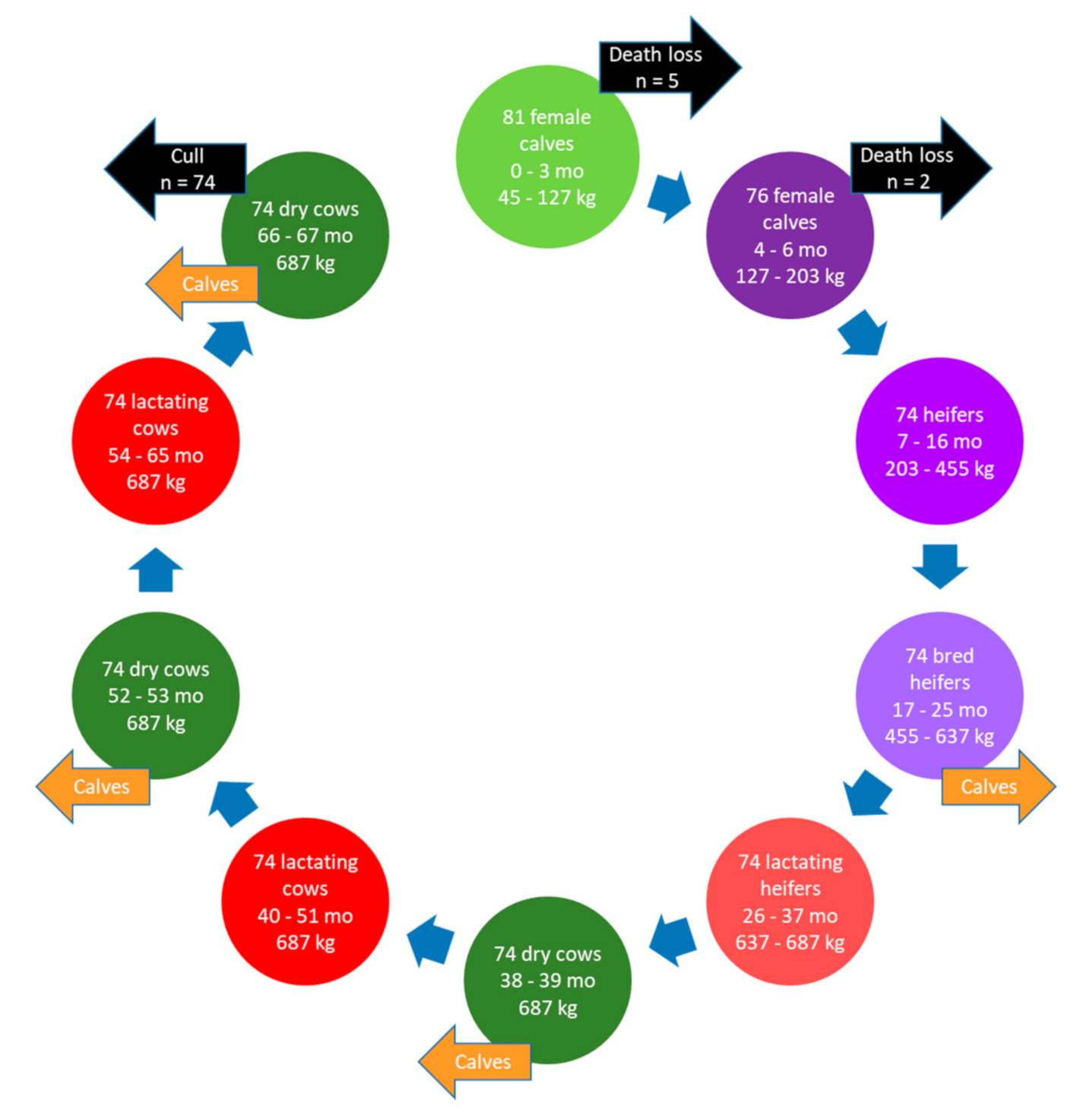
Herd size, dietary ingredients, housing, farm management, and environmental growing conditions of the farms simulated in the present study to evaluate CM as a GHG mitigation strategy differed substantially, but were typical of farms in these respective areas of the country. The AB farm grew barley grain in addition to the forages required for the herd and, therefore, purchased and imported less feed onto the farm. Consequently, the proportion of total GHG emissions from purchased feeds (plus transportation) was greater for the QC (range: 26.9% to 36.2%) versus AB (range: 14.4% to 16.2%) farm and, inversely, the proportion of total GHG emissions from on-farm feed production was greater in AB. However, for both farms, enteric CH
4 was the greatest contributor to kg CO
2e/kg FPCM, ranging from 34% to 40% for the various scenarios. Previous studies also report enteric CH
4 as the greatest contributor to the total GHG emissions from milk production in intensive dairying (range of 31.7% to 57.3% for individual farms in ON) [
45]).
After allocation to meat, the GHG intensities of milk to the farm-gate ranged from 0.85 to 1.02 kg CO
2e/kg FPCM for the QC farm scenarios and from 1.07 to 1.11 kg CO
2e/kg FPCM for the AB farm scenarios. The slightly greater emission intensity of the AB farm was partially attributed to the use of default Y
m values [
29] to estimate CH
4 emissions in the absence of measured values. The measured Y
m values from Benchaar et al. [
12] used for the QC herd were less than the default values used for the AB herd. In addition, the feed conversion efficiency (kg milk/kg DMI) was less for the AB herd compared with the QC herd. Improving feed conversion efficiency corresponds to a general trend for lower emissions per unit of milk produced [
4].
The GHG intensity values reported in the present study are consistent with many other North American studies, despite differences in assumptions and methodologies. Gerber et al. [
46] reported an average farm-gate emission intensity of 1.0 kg CO
2e/kg FPCM for North America. Thoma et al. [
4] and Capper et al. [
47] reported national averages of 1.23 (2008) and 1.35 (2007) kg CO
2e/kg FPCM, respectively (farm-gate), in the United States, while for the Canadian dairy industry (farm-gate), Vergé et al. [
29] reported an average GHG intensity of 1.0 kg CO
2e/kg milk, with slightly greater intensity in AB compared with QC (1.05 vs. 0.97 kg CO
2e/kg milk). However, the opposite regional trend was reported by Thivierge et al. [
48], with intensities of 0.95 to 0.99 kg CO
2e/kg FPCM for AB and 1.12 to 1.22 kg CO
2e/kg FPCM for QC. Jayasundara and Wagner-Riddle [
49] reported an intensity of 1.03 kg CO
2e/FPCM, ranging from 0.89 to 1.36 kg CO
2e/kg FPCM, for ON (farm-gate) in 2011. A more recent survey of 142 farms in ON reported a range of 0.44 to 1.73 kg CO
2e/kg FPCM, with a mean of 1.015 kg CO
2e/kg FPCM [
45].
It is often assumed that a decrease in the Y
m value of a diet, as reported by Benchaar et al. [
12] for a lactation diet containing CM, would lower the emission intensity of milk production. However, a decreased Y
m value due to a change in ingredient composition of the diet may not lower total GHG emissions of milk if emissions elsewhere in the system increase. This concept is illustrated by the QC farm scenarios, whereby the enteric CH
4-mitigating effect of CM did not offset the greater emission factor of CM compared with SBM when both meals were produced locally. Furthermore, using SBM from ON rather than QC did not alter the conclusion that, despite a CH
4-mitigating effect of CM, SBM is a better choice than CM produced in QC in terms of minimizing the GHG emissions associated with milk and meat production. However, the comparison between using these protein sources in the lactation diets differed when CM from SK was used; its low up-stream emission factor caused a 6.6% decrease in GHG intensity of milk compared with using SBM produced in QC. This result highlights the importance of considering the source of CM and, consequently, its up-stream emission factor, prior to recommending it as a CH
4 mitigation practice.
Emissions from imported protein meals include the CO
2 from fossil fuel used in producing the crop (fertilizer, field operations, machinery use, and so forth), N
2O emissions from the use of organic and inorganic fertilizers, carbon change in soils, and the CO
2 from the fossil fuels used to process the seed and transport it to the farm. Emissions from transporting CM from SK to the farm in QC were small (1.2%,
Figure 3) compared with the emissions associated with growing and processing the crop.
Globally, most SBM used in livestock feeding is imported from Argentina, Brazil, and the United States, with Canada being self-reliant in SBM. Using a consequential life cycle assessment, Dalgaard et al. [
50] calculated that SBM produced in Argentina and delivered to the Netherlands had a global warming potential of 0.721 kg CO
2e/kg if used to avoid palm oil and 0.344 kg CO
2e/kg to avoid rapeseed oil, respectively. The emission factors for SBM and CM in the present study (
Table 5) from Desjardins et al. [
35] were derived using the traditional attributional approach, and thus not directly comparable to those reported by Dalgaard et al. [
50]. Rotz et al. [
51] assumed an emission factor of 0.37 kg CO
2e/kg DM for SBM produced in the United States. In Canada, large differences in the emission factors of protein sources occur at the provincial scale, and these differences were shown in the present study to affect the GHG intensity of FPCM. Desjardins et al. [
35] reported that the emission intensities in 2011 for canola seed on an area basis were 2700 kg CO
2e/ha for QC and 530 kg CO
2e/ha for SK. The lower emission intensity in SK is due to the high adoption rate of practices such as no-till and reduced summer fallowing that favor the sequestration of carbon in soils. Additionally, the drier climate in SK leads to lower N
2O emissions, and large field sizes favor more efficient use of farm machinery and reduced fossil fuel consumption.
For the AB farm, the assessment of using CM rather than SBM as a protein source in lactation diets did not examine the location effect (emission factor) of producing the meals, because meals from multiple sources are typically not available in that region. The similar emission intensity of FPCM for SBM (A1) and CM
without (A3) scenarios indicates that, unless CM has a CH
4-mitigating effect, there is no basis for recommending CM as a GHG mitigation strategy for dairy farms in AB, although its nutritional properties make it a valuable protein source for dairy cows [
12,
15]. When the CH
4-mitigating property of CM was applied (AB2), the GHG intensity of milk decreased by 3% compared with that of SBM (A1) and by 4% compared with that of CM
without (A3). The decrease in the intensity of milk production when feeding CM
with was due solely to decreased enteric CH
4 emissions, as other sources of emissions remained stable and milk production and DMI were unchanged.
The decrease in Y
m value when feeding CM to dairy cows reported by Benchaar et al. [
12] needs further substantiation, as relatively few studies have examined the CH
4 mitigation potential of protein sources. Gidlund et al. [
52] reported a 6.6% numerical decrease in CH
4 emissions (g/kg DMI) when feeding high proportions of CM (24% of DM) versus SBM (15% of DM) to dairy cows fed grass silage-based diets. The mitigation effect was attributed to a shift in rumen fermentation towards an increased molar proportion of propionate, an alternative hydrogen sink to CH
4 in the rumen. However, Reynolds et al. [
15] reported no effect on CH
4 emissions when partially replacing SBM (9% of dietary DM) with CM (10% of diet DM) in the diet of lactating dairy cows. Thus, further research is needed before recommending feeding CM as a CH
4-mitigating practice for milk production.
Flysjö et al. [
53] indicated that the most important parameters affecting the GHG emissions of milk production were DMI and Y
m, which affect the quantity of enteric CH
4 produced, and the amount of N applied and the emission factor for direct N
2O emissions from soils, which determine the N
2O from crop production and contribute substantially to the emission factor of imported feeds. Our study shows that the emission factors for SBM and CM, as well as the Y
m value of the lactation diets, have significant effects on the GHG intensity of milk production, and the sensitivity analysis puts the relative impact of these factors into perspective. Although the sensitivity analysis examined only one factor at a time and did not consider interactions, it is considered a valuable tool in life cycle assessment [
44].
According to the sensitivity analysis, a 10% decrease in Ym of the lactation diet had a much greater impact on the GHG intensity of milk than a 10% decrease in the emission factor of the imported protein meals. However, for dairy farms, the emission factors of the imported protein meals vary substantially depending upon where the meals are produced, whereas the diet Ym values vary only slightly. For example, the emission factor for CM in the present study varied by 424% (0.29 to 1.23 CO2e/kg DM) depending upon where it was produced, whereas the Ym value of the lactation diets only varied by 13%.
Although the present study focused on the effects of protein source (emission factor of the meals and Y
m of the diet), the sensitivity analysis also included the impact of increasing milk production and improving feed conversion efficiency. It is well documented that improving animal performance decreases the GHG intensity of meat and milk production because fewer animals and less feed are needed to produce a certain amount of product [
7]. Our analysis showed that, in comparison with the effects of Y
m value and protein source emission factor, changes in feed conversion efficiency due to increased milk production or decreased DMI (due to increased digestibility) clearly have larger potential to reduce the GHG intensity of milk, as indicated by sensitivity indices >0.63. Increased milk production can be achieved through genetic selection, nutrition, and management [
54]. However, the impact of increased milk production in the present study is a “best-case” scenario, because it assumed no increase in DMI. In reality, increased milk production per cow is usually accompanied by more feed consumed per cow, but with a greater portion of the feed partitioned toward milk instead of maintenance and body growth. A decrease in DMI due to increased digestibility decreased the GHG intensity of milk because less feed was required and, therefore, emissions from the animal, manure, crops, and imported feeds were also decreased.
This LCA highlights the relative impacts of dietary protein supplements on the GHG intensity of milk by accounting for differences in up-stream emissions of producing SBM and CM and a potential enteric CH
4-mitigating effect of CM. While the overall impact of the dietary protein source on the GHG intensity of milk was small (a decrease of 6.6% in the east and 3% in the west), it was not inconsequential. In comparison, the GHG intensity of milk produced in ON decreased by 1% annually over a 20-year period (2011 vs. 1991) owing to numerous improvements in production efficiency. On the other hand, it should be noted that there is considerable uncertainty associated with the inputs, farm characteristics, emission factors, and assumptions used to model the emissions in the present study, thus caution must be used when interpreting the results [
44].








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}