
Combination of Quercetin and Vitamin E Supplementation Promotes Yolk Precursor Synthesis and Follicle Development in Aging Breeder Hens via Liver–Blood–Ovary Signal Axis

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Birds, Management, and Experimental Treatment
2.2. Clinical Blood Parameters
2.3. Reproductive Organ Characteristics of Aging Breeder Hens
2.4. Biochemical Analysis
2.4.1. Triglyceride (TG), Total Cholesterol (T-CHO), Aspartate Aminotransferase (AST) and Alanine Aminotransferase (ALT)
2.4.2. Vitellogenin (VTG), and Very Low Density Lipoprotein y (VLDLy)
2.4.3. Oxidative Stress Parameters Assay in the Liver, Serum, and Ovary
2.5. Morphological and Histological Analyses/Histological Staining
2.6. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
2.7. Statistical Analysis
3. Results
3.1. Effects of Quercetin (Q), Vitamin E (VE), and Their Combination (Q + VE) on Organ Characteristics of Aging Breeder Hens
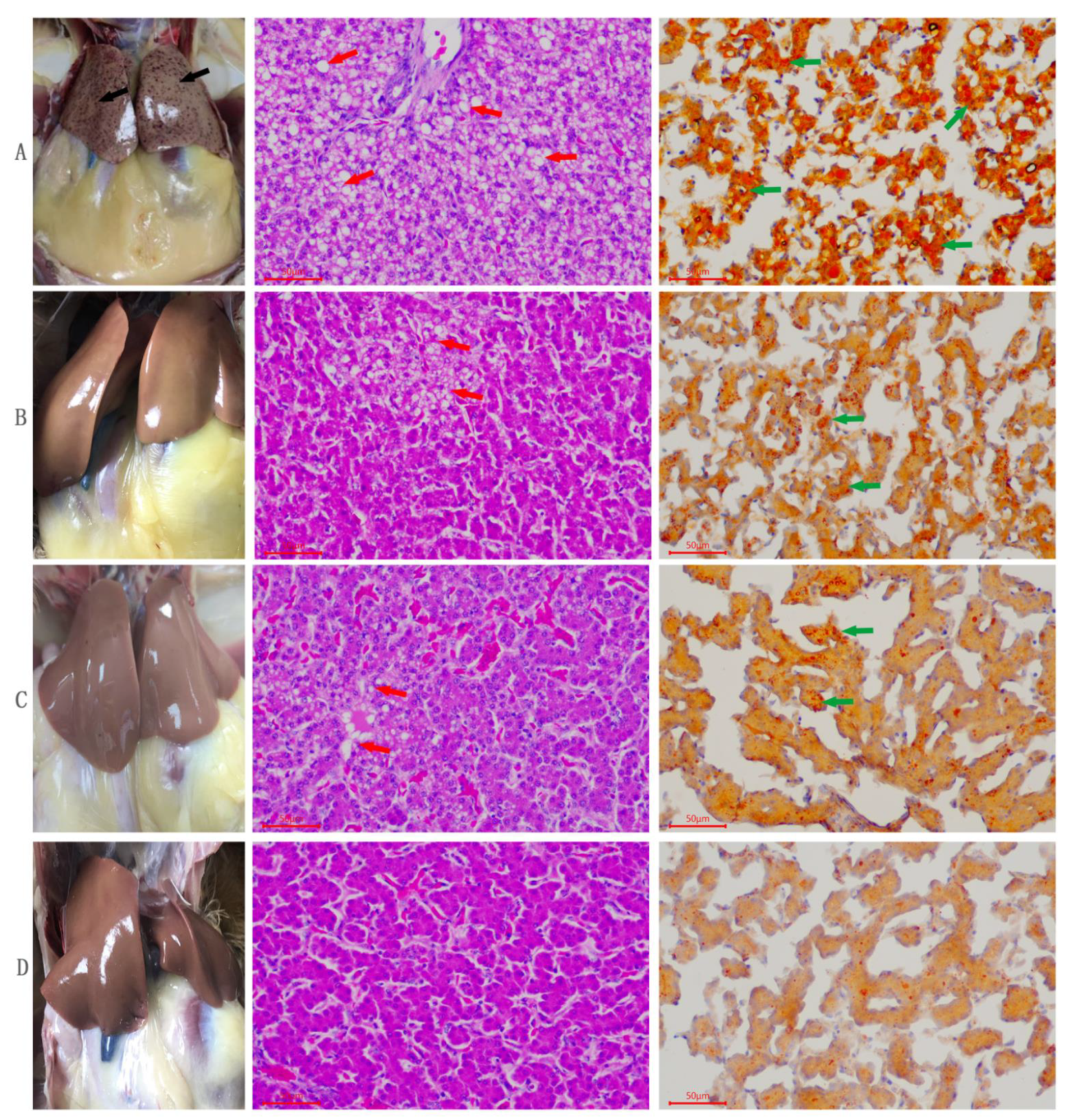
3.2. Effects of Quercetin, Vitamin E, and Their Combination (Q + VE) on Liver Morphological and Histological Changes of Aging Breeder Hens
3.3. Effects of Q, VE, and Q + VE on the Biochemical Changes in the Liver, Serum, and Ovary of the Aging Breeder Hens
3.3.1. TG and TC Levels in the Liver, Serum, and Ovary
3.3.2. VLDLy and VTG Levels in the Liver, Serum, and Ovary
3.4. Effects of Q, VE, and Q + VE on the Antioxidant Capacity and MDA Levels of the Liver, Blood, and Ovary of Aging Breeder Hens
3.4.1. Antioxidant Capacity of the Liver, Blood, and Ovary of Aging Breeder Hens
3.4.2. MDA Levels in the Liver, Blood, and Ovary of Aging Breeder Hens
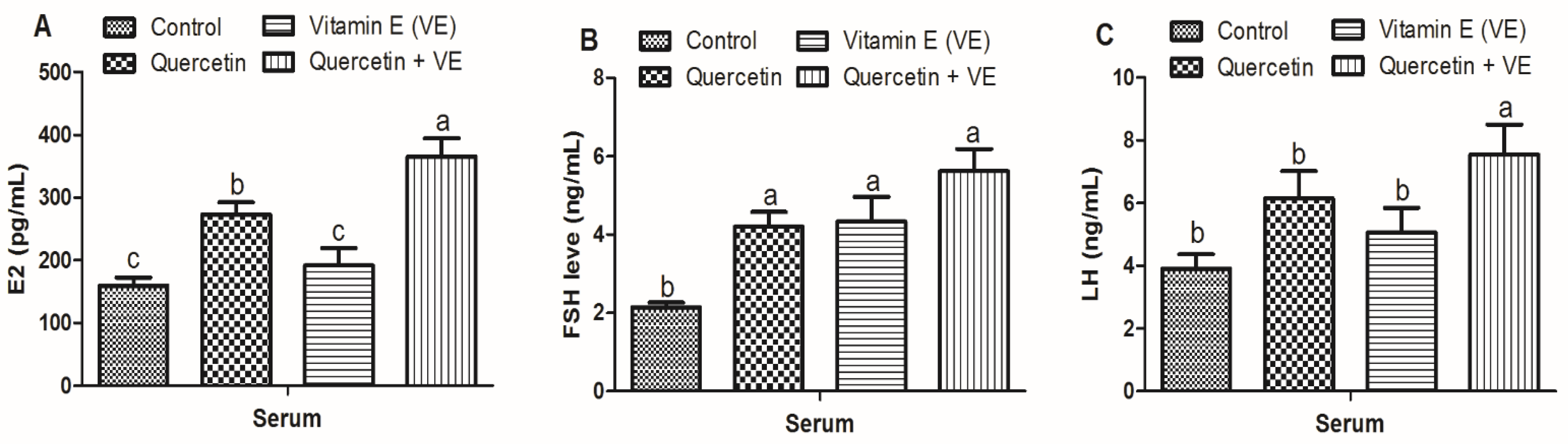
3.5. Effects of Quercetin (Q), Vitamin E (VE), and Their Combination (Q + VE) on Serum Hormone Levels of Aging Breeder Hens
3.6. Effects of Q, VE, and Q + VE on the Levels of mRNA Expression of Hormone Receptors in the Ovary of Aging Breeder Hens
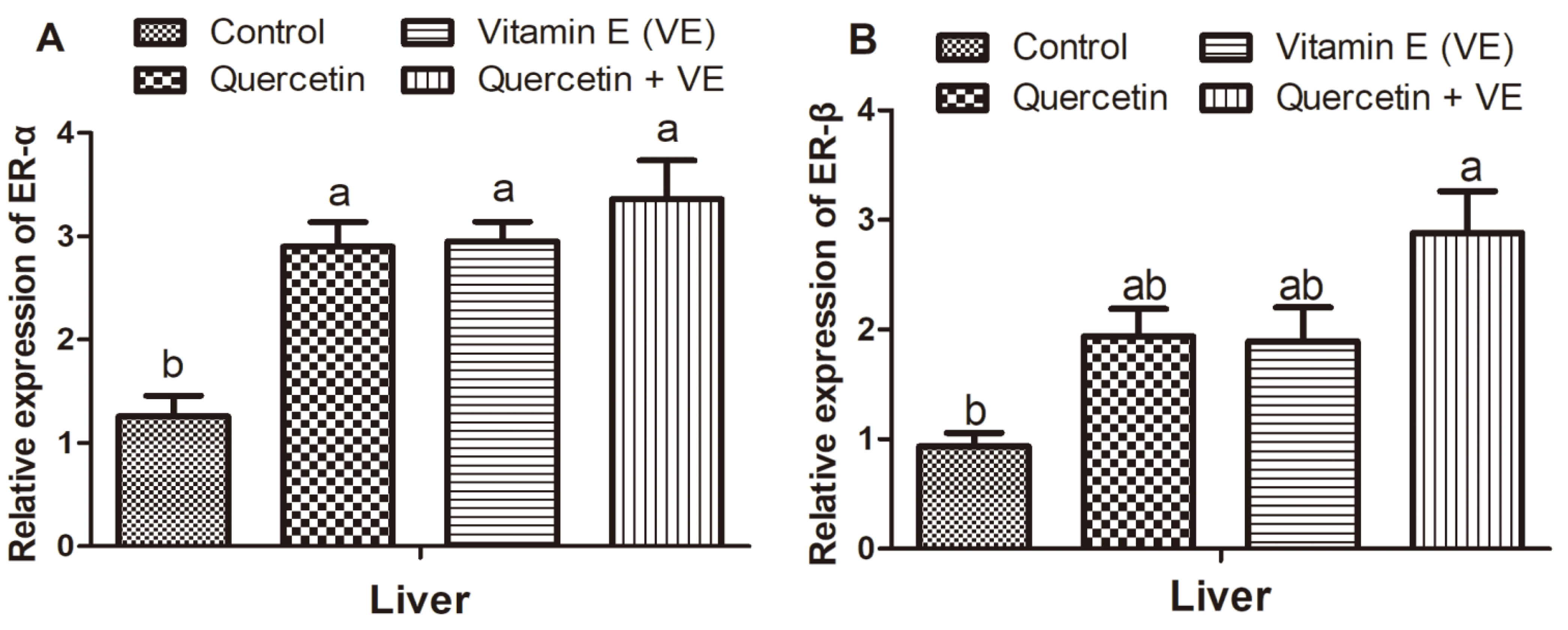
3.7. Effects of Q, VE, and Q + VE on the mRNA Expression of Liver ER-α and ER-β of the Aging Breeder Hens
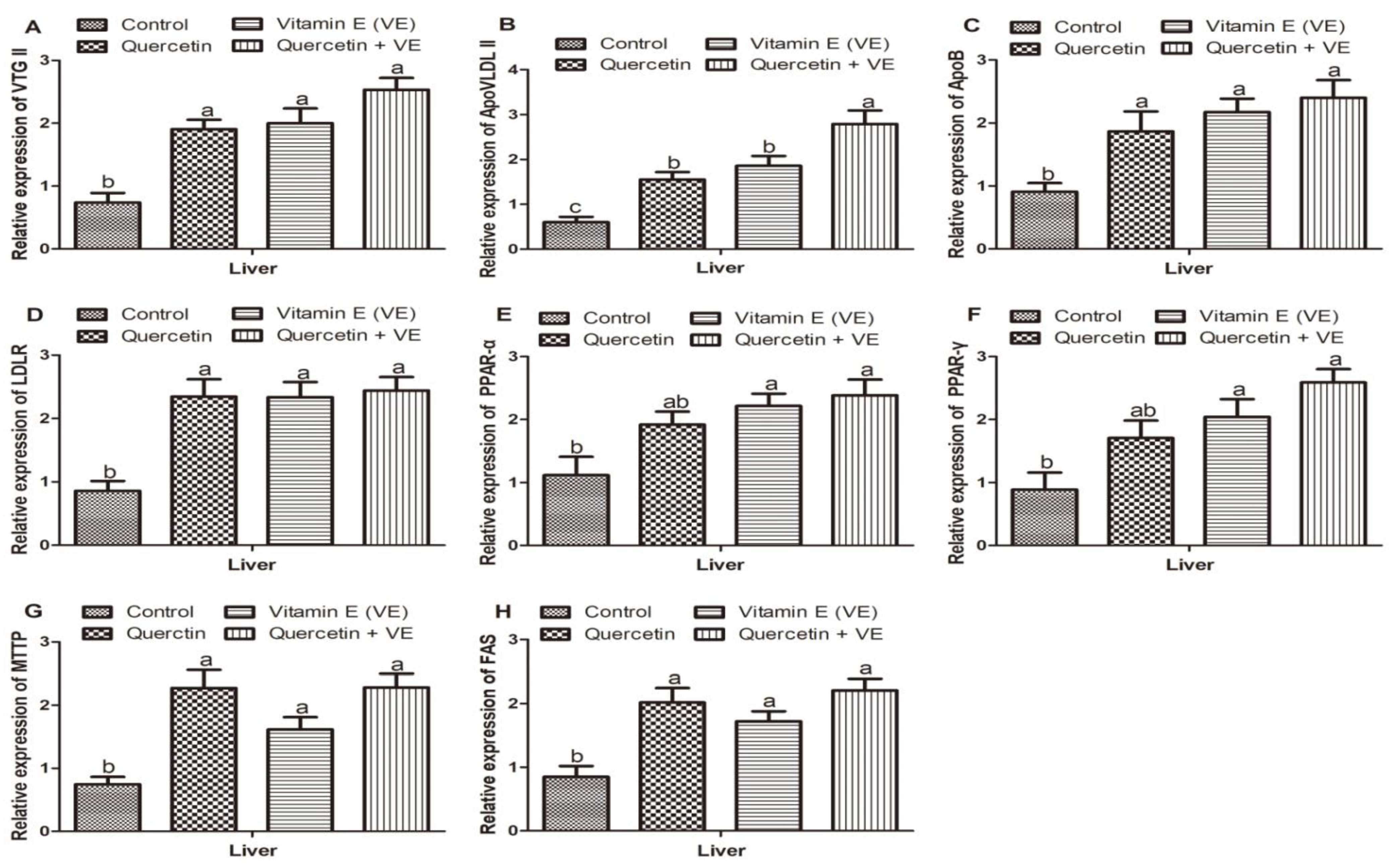
3.8. Effects of Q, VE, and Q + VE on the Expression of mRNAs Related to Yolk Precursor Synthesis in the Liver of Aging Breeder Hens
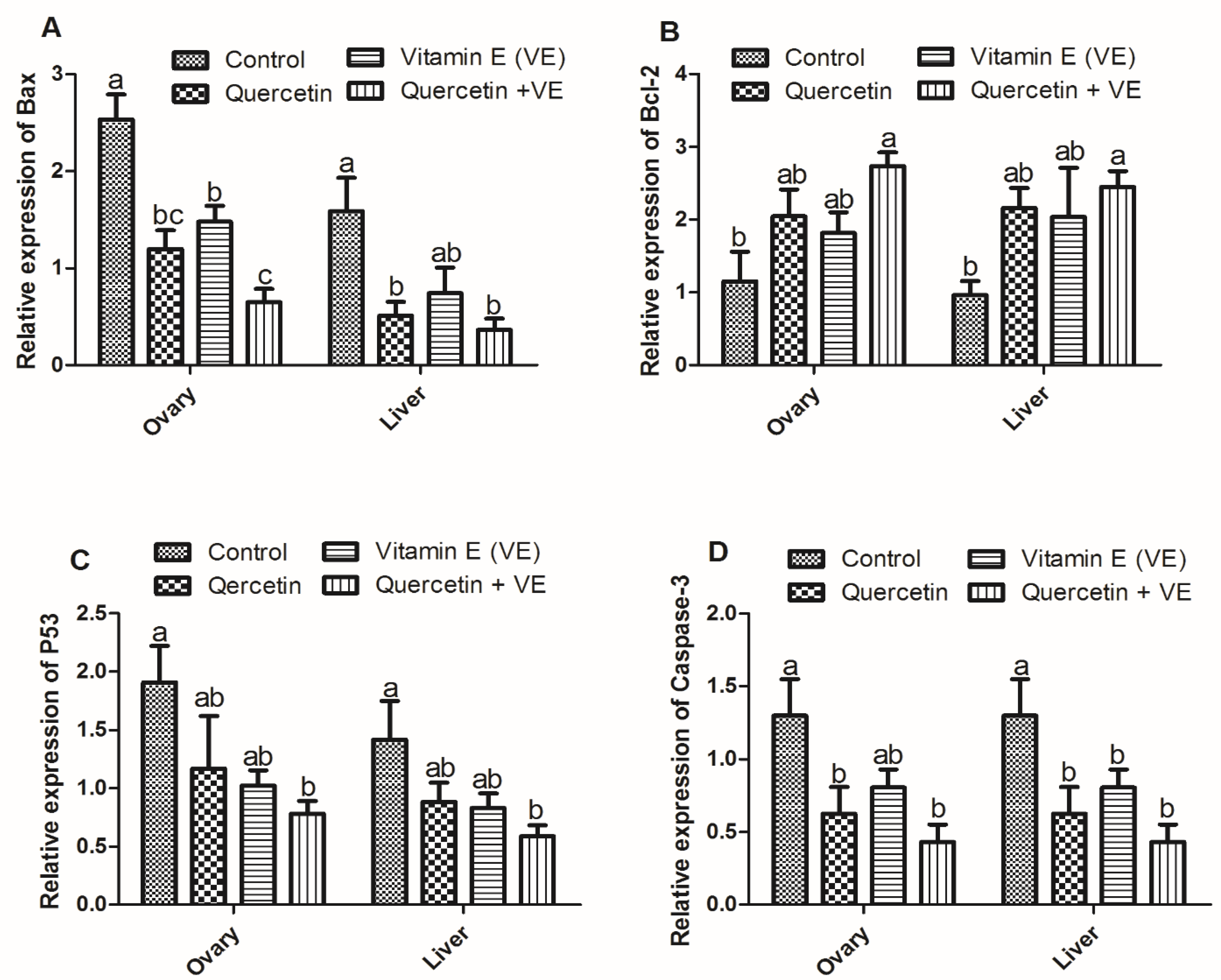
3.9. Effects of Q, VE, and Q + VE on the Expression of Apoptotic Genes in the Liver and Ovary of the Aging Breeder Hens
Relative Expression of Apoptosis-Related Genes in the Ovary and Liver
3.10. Effects of Q, VE, and Q + VE on Serum AST and ALT Levels, and the mRNA Expression of Inflammation and Anti-Inflammation Related Cytokines in the Liver of Aging Breeder Hens
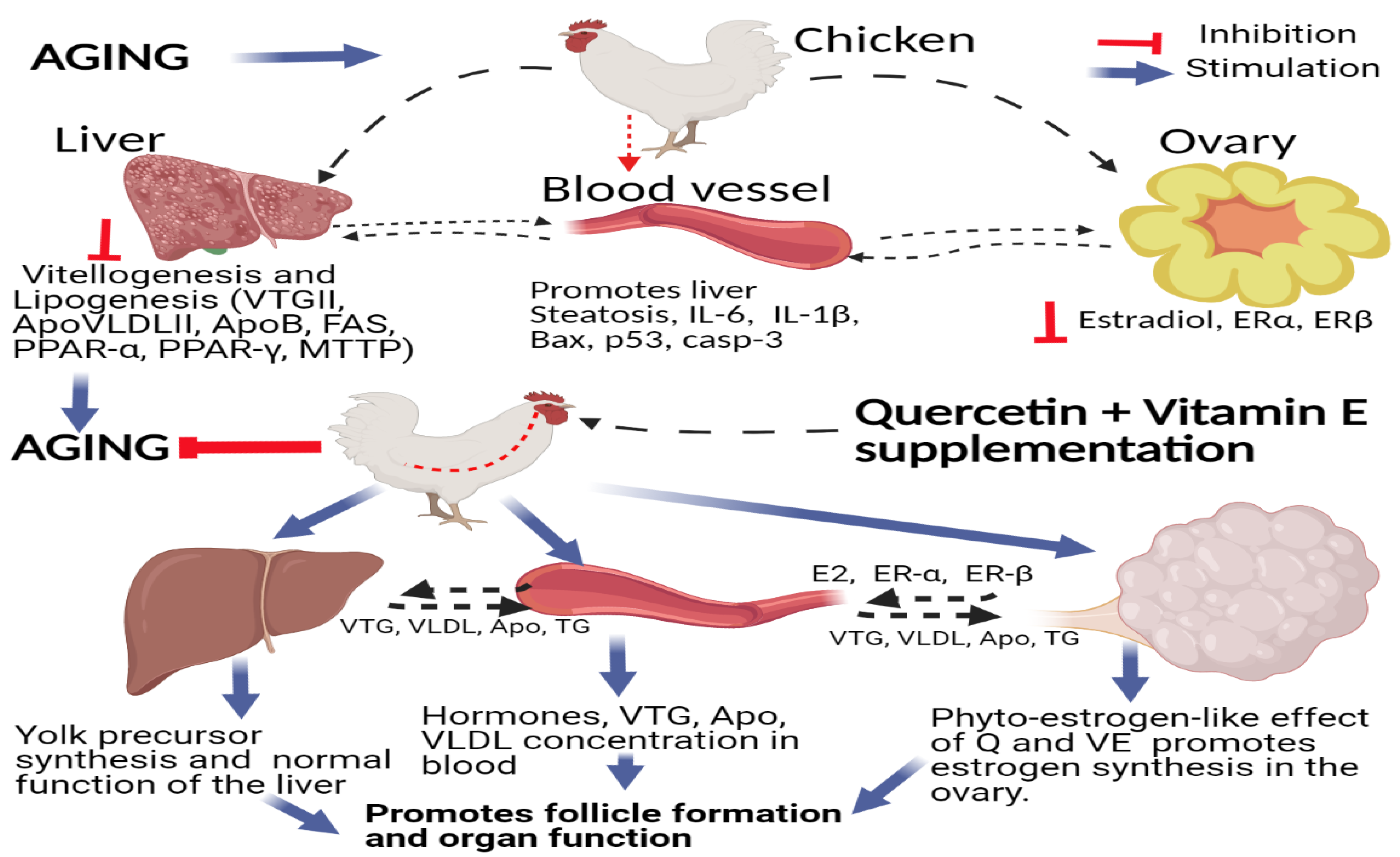
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Durape, N.M. Phytochemicals improve semen quality and fertility. World Poult. 2007, 23, 6. [Google Scholar]
- Mol, B.W.; Zoll, M. Fertility preservation for age-related fertility decline. Lancet. 2015, 385, 507. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.T.; Lin, X.; Mi, Y.L.; Zeng, W.D.; Zhang, C.Q. Age-related changes of yolk precursor formation in the liver of laying hens. J. Zhejiang Univ. Sci. B 2018, 19, 390–399. [Google Scholar] [CrossRef]
- Yang, H.; Qazi, I.H.; Pan, B.; Angel, C.; Guo, S.; Yang, J.; Zhang, Y.; Ming, Z.; Zeng, C.; Meng, Q.; et al. Dietary Selenium Supplementation Ameliorates Female Reproductive Efficiency in Aging Mice. Antioxidants 2019, 8, 634. [Google Scholar] [CrossRef] [Green Version]
- Qazi, I.H.; Cao, Y.; Yang, H.; Angel, C.; Pan, B.; Zhou, G.; Han, H. Impact of Dietary Selenium on Modulation of Expression of Several Non-Selenoprotein Genes Related to Key Ovarian Functions, Female Fertility, and Proteostasis: A Transcriptome-Based Analysis of the Aging Mice Ovaries. Biol. Trace Elem. Res. 2021, 199, 633–648. [Google Scholar] [CrossRef]
- Ishii, M.; Yamauchi, T.; Matsumoto, K.; Watanabe, G.; Taya, K.; Chatani, F. Maternal age and reproductive function in female Sprague-Dawley rats. J. Toxicol. Sci. 2012, 37, 631–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Meir, A.; Burstein, E.; Borrego-Alvarez, A.; Chong, J.; Wong, E.; Yavorska, T.; Naranian, T.; Chi, M.; Wang, Y.; Bentov, Y.; et al. Coenzyme Q10 restores oocyte mitochondrial function and fertility during reproductive aging. Aging Cell 2015, 14, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Okudan, N.; Belviranli, M. Effects of exercise training on hepatic oxidative stress and antioxidant status in aged rats. Arch. Physiol. Biochem. 2016, 122, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Hoare, M.; Das, T.; Alexander, G. Ageing, telomeres, senescence, and liver injury. J. Hepatol. 2010, 53, 950–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buyuk, E.; Nejat, E.; Neal-Perry, G. Determinants of female reproductive senescence: Differential roles for the ovary and the neuroendocrine axis. Semin. Reprod. Med. 2010, 28, 370–379. [Google Scholar] [CrossRef]
- Yilmaz, O.; Prat, F.; Ibañez, A.J.; Amano, H.; Koksoy, S.; Sullivan, C.V. Estrogen-induced yolk precursors in European sea bass, Dicentrarchus labrax: Status and perspectives on multiplicity and functioning of vitellogenins. Gen. Comp. Endocrinol. 2015, 221, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Amevor, F.K.; Feng, Q.; Kang, X.; Song, W.; Zhu, Q.; Wang, Y.; Li, D.; Zhao, X. Sexual Maturity Promotes Yolk Precursor Synthesis and Follicle Development in Hens via Liver-Blood-Ovary Signal Axis. Animals 2020, 10, 2348. [Google Scholar] [CrossRef]
- Zhang, C.Q.; Shimada, K.; Saito, N.; Kansaku, N. Expression of messenger ribonucleic acids of luteinizing hormone and follicle-stimulating hormone receptors ingranulosa and theca layers of chicken preovulatory follicles. Gen. Comp. Endocrinol. 1997, 105, 402–409. [Google Scholar] [CrossRef]
- Johnson, A. Reproduction in the female. In Sturkie’s Avian Physiology, 6th ed.; Scanes, C.G., Ed.; Academic Press: London, UK, 2015; pp. 653–654. [Google Scholar]
- Alvarenga, R.R.; Zangeronimo, M.G.; Pereira, L.J.; Rodrigues, P.B.; Gomide, E.M. Lipoprotein metabolism in poultry. World’s Poult. Sci. J. 2011, 67, 431–440. [Google Scholar] [CrossRef]
- Stanley, J.A.; Sivakumar, K.K.; Arosh, J.A.; Burghardt, R.C.; Banu, S.K. Edaravone mitigates hexavalent chromium induced oxidative stress and depletion of antioxidant enzymes while estrogen restores antioxidant enzymes in the rat ovary in F1 offspring. Biol. Reprod. 2014, 91, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, A.; Aponte-Mellado, A.; Premkumar, B.J.; Shaman, A.; Gupta, S. The effects of oxidative stress on female reproduction: A review. Reprod. Biol. Endocrinol. 2012, 10, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Lin, X.; Zhang, S.; Guo, C.; Li, J.; Mi, Y.; Zhang, C. Lycopene ameliorates oxidative stress in the aging chicken ovary via activation of Nrf2/HO-1 pathway. Aging 2018, 10, 2016–2036. [Google Scholar] [CrossRef]
- Liu, X.; Lin, X.; Mi, Y.; Li, J.; Zhang, C. Grape seed proanthocyanidin extract prevents ovarian aging by inhibiting oxidative stress in the hens. Oxid. Med. Cell Longev. 2018. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Yin, Y.; Ye, X.; Zeng, M.; Zhao, Q.; Keefe, D.L.; Liu, L. Resveratrol protects against age-associated infertility in mice. Hum. Reprod. 2013, 28, 7–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khedr, N.F. Protective effect of mirtazapine and hesperidin on cyclophosphamide-induced oxidative damage and infertility in rat ovaries. Exp. Biol. Med. 2015, 240, 1682–1689. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.R.; Gu, H.; Chang, L.L.; Wang, Z.Y.; Tong, H.B.; Zou, J.M. Safety evaluation of daidzein in laying hens: Part I. Effects on laying performance, clinical blood parameters, and organs development. Food. Chem. Toxicol. 2013, 55, 684–688. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, Q.; Shao, D.; Zhao, X.; Hu, Y.; Tong, H.B.; Shi, S.R. Effects of daidzein on egg production and gonado expression in the ovary and follicles of laying hens. Eur. Poult. Sci. 2016, 80, 1612–9199. [Google Scholar] [CrossRef]
- McDermott, J.H. Antioxidant nutrients: Current dietary recommendations and research update. J. Am. Pharm. Assoc. 2000, 40, 785–799. [Google Scholar] [CrossRef]
- Liu, H.N.; Liu, Y.; Hu, L.L.; Suo, Y.L.; Zhang, L.; Jin, F.; Feng, X.A.; Teng, N.; Li, Y. Effects of dietary supplementation of quercetin on performance, egg quality, cecal microflora populations, and antioxidant status in laying hens. Poult. Sci. 2014, 93, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.X.; Maria, T.C.; Zhou, B.; Xiao, F.L.; Wang, M.; Mao, Y.J.; Li, Y. Quercetin improves immune function in Arbor Acre broilers through activation of NF-κB signaling pathway. Poult. Sci. 2020, 99, 2305. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.X.; Chaudhry, M.T.; Yao, J.Y.; Wang, S.N.; Zhou, B.; Wang, M.; Han, C.Y.; You, Y.; Li, Y. Effects of phyto-oestrogen quercetin on productive performance, hormones, reproductive organs and apoptotic genes in laying hens. J. Anim. Physiol. Anim. Nutr. 2018, 102, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Hu, M.J.; Wang, Y.Q.; Cui, Y.L. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.Y.; Latshaw, J.D.; Lee, H.O.; Min, D.B. α-tocopherol content and oxidative stability of egg yolk as related to dietary α-tocopherol. J. Food Sci. 1998, 63, 919–922. [Google Scholar] [CrossRef]
- Abedi, P.; Tabatabaei Vakili, S.; Mamouei, M.; Aghaei, A. Effect of different levels of dietary vitamin E on reproductive and productive performances in Japanese quails (Coturnix coturnix japonica). Vet. Res. Forum. 2017, 8, 353–359. [Google Scholar] [PubMed]
- Lin, Y.F.; Chang, S.J.; Hsu, A.L. Effects of supplemental vitamin E during the laying period on the reproductive performance of Taiwan native chickens. Br. Poult. Sci. 2004, 45, 807–814. [Google Scholar] [CrossRef]
- Lin, Y.F.; Chang, S.J.; Yang, J.R.; Lee, Y.P.; Hsu, A.L. Effects of supplemental vitamin E during the mature period on the reproduction performance of Taiwan Native Chicken cockerels. Br. Poult. Sci. 2005, 46, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Bollengier-Lee, S.; Williams, P.E.; Whitehead, C.C. Optimal dietary concentration of vitamin E for alleviating the effect of heat stress on egg production in laying hens. Br. Poult. Sci. 1999, 40, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Grobas, S.; Méndez, J.; Lopez, B.C.; De, B.C.; Mateos, G.G. Effect of vitamin E and A supplementation on egg yolk alpha-tocopherol concentration. Poult. Sci. 2002, 81, 376–381. [Google Scholar] [CrossRef]
- Bollengier-Lee, S.; Mitchell, M.A.; Utomo, D.B.; Williams, P.E.; Whitehead, C.C. Influence of high dietary vitamin E supplementation on egg production and plasma characteristics in hens subjected to heat stress. Br. Poult. Sci. 1998, 39, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Puthpongsiriporn, U.; Scheideler, S.E.; Sell, J.L.; Beck, M.M. Effects of vitamin E and C supplementation on performance, in vitro lymphocyte proliferation, and antioxidant status of laying hens during heat stress. Poult. Sci. 2001, 80, 1190–1200. [Google Scholar] [CrossRef] [PubMed]
- Çiftci, M.; Ertas, O.N.; Guler, T. Effects of vitamin E and vitamin C dietary supplementation on egg production and egg quality of laying hens exposed to a chronic heat stress. Rev. Med. Vet. 2005, 156, 107–111. [Google Scholar]
- Tian, Y.; Niu, J.; Zhu, Q.; Zhang, Y.; Yin, H.; Wang, Y.; Li, D.; Shu, G.; Ye, L.; Zhang, Z.; et al. Breeding of Tianfu broilers. Heilongjiang Anim. Husb. Vet. Med. 2021, 6, 36–41. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using RT-PCR and the 2 (−Delta C (T)) methods. Methods 2001, 25, 402–440. [Google Scholar] [CrossRef] [PubMed]
- Banu, S.K.; Stanley, J.A.; Sivakumar, K.K.; Arosh, J.A.; Burghardt, R.C. Resveratrol protects the ovary against chromium-toxicity by enhancing endogenous antioxidant enzymes and inhibiting metabolic clearance of estradiol. Toxicol. Appl. Pharmacol. 2016, 303, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Q.; Gao, B.W.; Wang, J.; Ren, Q.L.; Chen, J.F.; Ma, Q.; Zhang, Z.J.; Xing, B.S. Critical role of foxO1 in granulosa cell apoptosis caused by oxidative stress and protective effects of grape seed procyanidin B2. Oxid. Med. Cell Longev. 2016, 6147345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalia, A.M.; Loh, T.C.; Sazili, A.Q.; Jahromi, M.F.; Samsudin, A.A. Effects of vitamin E, inorganic selenium, bacterial organic selenium, and their combinations on immunity response in broiler chickens. BMC Vet. Res. 2018, 14, 249. [Google Scholar] [CrossRef]
- Dong, Y.; Lei, J.; Zhang, B. Effects of dietary quercetin on the antioxidative status and cecal microbiota in broiler chickens fed with oxidized oil. Poult. Sci. 2020, 99, 4892–4903. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Zhao, L.H.; Mosenthin, R.; Zhang, J.Y.; Ji, C.; Ma, Q.G. Protective Effect of Vitamin E on laying performance, antioxidant capacity, and immunity in laying hens challenged with Salmonella Enteritidis. Poult. Sci. 2019, 98, 5847–5854. [Google Scholar] [CrossRef] [PubMed]
- Sujen, E.S.; Giuseppina, B.; Simona, B.; Francesca, G. The phytoestrogen quercetin impairs steroidogenesis and angiogenesis in swine granulosa cells in vitro. J. Biomed. Biotechnol. 2009, 2009, 419891. [Google Scholar] [CrossRef] [Green Version]
- Ososki, A.L.; Kennelly, E.J. Phytoestrogens: A review of the present state of research. Phytother. Res. 2003, 17, 845–869. [Google Scholar] [CrossRef]
- Zhao, R.Q.; Wang, Y.J.; Zhou, Y.C.; Ni, Y.; Lu, L.; Grossmann, R.; Chen, J. Dietary daidzein influences laying performance of ducks (Anas platyrhynchos) and early post- hatch growth of their hatchlings by mod- ulating gene expression. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2004, 138, 459–466. [Google Scholar] [CrossRef]
- Cos, P.; De, B.T.; Apers, S.; Vanden, B.D.; Pieters, L.; Vlietinck, A.J. Phytoestrogens: Recent development. Planta Med. 2003, 69, 589–599. [Google Scholar] [PubMed] [Green Version]
- Claire D’Andre, H.; Paul, W.; Shen, X.; Jia, X.; Zhang, R.; Sun, L.; Zhang, X. Identification and characterization of genes that control fat deposition in chickens. J. Anim. Sci. Biotechnol. 2013, 4, 43–58. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, T.; Xu, C.; Wang, D.; Ren, J.; Li, Y.; Tian, Y.; Wang, Y.; Jiao, Y.; Kang, X.; et al. Transcriptome profile of liver at different physiological stages reveals potential mode for lipid metabolism in laying hens. BMC Genom. 2015, 16, 763–775. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Gupta, S.; Sharma, R.K. Role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2005, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Naseer, Z.; Ahmad, E.; Epikmen, E.T.; Uçan, U.; Boyacioğlu, M.; İpek, E.; Akosy, M. Quercetin supplemented diet improves follicular development, oocyte quality, and reduces ovarian apoptosis in rabbits during summer heat stress. Theriogenology 2017, 96, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Devine, P.J.; Perreault, S.D.; Luderer, U. Roles of reactive oxygen species and antioxidants in ovarian toxicity. Biol. Reprod. 2012, 86, 27. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Lin, J.; Mi, Y.; Zhang, C. Quercetin attenuates cadmium-induced oxidative damage and apoptosis in granulosa cells from chicken ovarian follicles. Reprod. Toxicol. 2011, 31, 477–485. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content (%) | Nutrient | % |
|---|---|---|---|
| Corn | 56.4 | Metabolic Energy (ME) | 10.98 MJ/kg |
| Soybean meal | 26.2 | Crude protein | 17.04 |
| Wheat bran | 2.4 | Crude fat | 3.47 |
| Corn germ meal | 3.5 | Crude fibre | 2.68 |
| Lard | 0.8 | Calcium | 3.4 |
| Limestone (fine) | 2.3 | Total phosphorus | 0.63 |
| Limestone (coarse) | 6 | Available phosphorus | 0.37 |
| Dicalcium phosphate | 1.34 | Lysine | 0.86 |
| Sodium chloride | 0.24 | Methionine | 0.39 |
| Choline chloride | 0.12 | ||
| Vitamin premix 1 + mineral premix 2 | 0.7 | ||
| Total | 100 | ||
| Gene | Sequence (5′-3′) | Product Length (bp) | Annealing Temperature (℃) | Accession Number |
|---|---|---|---|---|
| FSHR | F: ACCACACGTGCCTCTGTGAA R: GCTCCCTTCGGAATGACTCT | 136 | 59.17 | NM_205079.1 |
| LHR | F: CGGATACACAACGATGCCCT R: TTTATCCAGAGGCGGCAGTC | 159 | 59.82 | NM_204936.1 |
| VLDLR/LR8 | F: AACGAGGCAGTCTATGGTGC R: TGTTGAATCCTCCACATCTCAG | 271 | 57.79 | NM_205229.1 |
| VTGII | F: AACTACTCGATGCCCGCAAA R: ACCAGCAGTTTCACCTGTCC | 179 | 58 | NM_001031276.1 |
| ApoVLDL-II | F: CCTTAGCACCACTGTCCCTG R: AGCTCTAGGGGACACCTTGT | 130 | 58 | NM_205483.2 |
| ApoB | F: ACACTTCGGGCTATTGGA R: TGCCTGTATGGCTGCTTT | 129 | 60 | NM_001044633.1 |
| LDLR | F: GGGAACCTCTATTGGGCCG R: CAACATGGGATCCAACGCGA | 270 | 59.85 | NM_204452.1 |
| ER-α | F: TGTGCTGTGTGCAACGACTA R: CAGGCCTGGCAACTCTTTCT | 167 | 57 | NM_205183.2 |
| ER-β | F: GGCTGCAACCCGTGTAAAAG R: GCCCAGCCAATCATGTGAAC | 189 | 58 | NM_204794.2 |
| PPAR-α | F: AGGCCAAGTTGAAAGCAGAA R: TTTCCCTGCAAGGATGACTC | 155 | 60 | NM_001001464.1 |
| PPAR-γ | F: TGACAGGAAAGACGACAGACA R: CTCCACAGAGCGAAACTGAC | 164 | 59 | NM_001001460.1 |
| MTTP | F: GTTCTGAAGGACATGCGTGC R: GATGTCTAGGCCGTACGTGG | 120 | 58 | NM_001109784.2 |
| FAS | F: GCTAAGATGGCATTGCACGG R: TCCATTCAGTTCCAGACGGC | 135 | 58 | NM_205155.1 |
| Bcl- 2 | F: ATCGTCGCCTTCTTCGAGTT R: ATCCCATCCTCCGTTGTCCT | 150 | 59 | Z11961.1 |
| Bax | F: GTGATGGCATGGGACATAGCTC R: TGGCGTAGACCTTGCGGATAA | 90 | 58 | XM_422067.4 |
| Caspase-3 | F: ACTCTGGAATTCTGCCTGATGACA R: CATCTGCATCCGTGCCTGA | 129 | 59 | NM_204725.1 |
| p53 | F:GAGATGCTGAAGGAGATCAATGAG R: GTGGTCAGTCCGAGCCTTTT | 145 | 58 | X13057.1 |
| IL-10 | F: GGAGAGAGCGGAGGTTTCG R: TCCCGTTCTCATCCATCTGC | 118 | 59.86 | XM_025143715.1 |
| IL-6 |
F: CTGCAGGACGAGATGTGCAA R: AGGTCTGAAAGGCGAACAGG | 175 | 60.67 | NM_204628.1 |
| IL-1β | F: TGCCTGCAGAAGAAGCCTCG R: GACGGGCTCAAAAACCTCCT | 204 | 60.25 | NM_204524.1 |
| GAPDH | F: TCCTCCACCTTTGATGCG R: GTGCCTGGCTCACTCCTT | 144 | 60 | NM_204305.1 |
| Parameters | Control | Quercetin | Vitamin E | Q + VE |
|---|---|---|---|---|
| Live Body weight (g) | 2602.81 ± 11.96 | 2855.94 ± 310.09 | 2637.50 ± 223.17 | 2675.00 ± 214.41 |
| Oviduct index (%) | 2.15 ± 0.62 c | 2.57 ± 0.45 ac | 2.05 ± 0.56 ac | 2.75 ± 0.52 a |
| Oviduct length (cm) | 53.43 ± 9.63 b | 63.71 ± 10.10a | 57.89 ± 7.54 ab | 64.60 ± 9.06 a |
| Liver index (%) | 2.10 ± 0.54 | 2.32 ± 0.43 | 0.32 ± 0.57 | 2.55 ± 0.69 |
| Ovary index (%) | 1.58 ± 0.41 b | 2.35 ± 0.52 a | 2.24 ± 0.52 a | 2.32 ± 0.53 a |
| Follicle No. (F1-F3, >8 mm in diameter) | 5.25 ± 1.48 b | 7.31 ± 1.99 a | 7.13 ± 1.02 a | 7.38 ± 1.09 a |
| Follicle F1 index (%) | 0.51 ± 0.15 b | 0.65 ± 0.08 ab | 0.67 ± 0.17 a | 0.71 ± 0.18 a |
| Follicle F2 index (%) | 0.31 ± 0.14 b | 0.53 ± 0.09 a | 0.50 ± 0.08 a | 0.57 ± 0.21 a |
| Follicle F3 index (%) | 0.20 ± 0.09 b | 0.32 ± 0.09 a | 0.30 ± 0.07 a | 0.32 ± 0.10 a |
| Follicle diameter (mm) F1 | 28.74 ± 5.56 c | 34.83 ± 5.09 b | 33.54 ± 0.89 b | 39.57 ± 4.12 a |
| Follicle diameter (mm) F2 | 22.90 ± 4.93 c | 31.19 ± 4.84 b | 30.07 ± 4.12 b | 35.06 ± 4.73 a |
| Follicle diameter (mm) F3 | 20.23 ± 5.85 c | 26.48 ± 3.84 b | 25.55 ± 3.83 b | 31.47 ± 3.48 a |
| Abdominal fat index (%) | 4.60 ± 1.55 | 3.73 ± 1.94 | 3.90 ± 3.10 | 2.82 ± 1.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amevor, F.K.; Cui, Z.; Du, X.; Ning, Z.; Shu, G.; Jin, N.; Deng, X.; Tian, Y.; Zhang, Z.; Kang, X.; et al. Combination of Quercetin and Vitamin E Supplementation Promotes Yolk Precursor Synthesis and Follicle Development in Aging Breeder Hens via Liver–Blood–Ovary Signal Axis. Animals 2021, 11, 1915. https://doi.org/10.3390/ani11071915
Amevor FK, Cui Z, Du X, Ning Z, Shu G, Jin N, Deng X, Tian Y, Zhang Z, Kang X, et al. Combination of Quercetin and Vitamin E Supplementation Promotes Yolk Precursor Synthesis and Follicle Development in Aging Breeder Hens via Liver–Blood–Ovary Signal Axis. Animals. 2021; 11(7):1915. https://doi.org/10.3390/ani11071915
Chicago/Turabian StyleAmevor, Felix Kwame, Zhifu Cui, Xiaxia Du, Zifan Ning, Gang Shu, Ningning Jin, Xun Deng, Yaofu Tian, Zhichao Zhang, Xincheng Kang, and et al. 2021. "Combination of Quercetin and Vitamin E Supplementation Promotes Yolk Precursor Synthesis and Follicle Development in Aging Breeder Hens via Liver–Blood–Ovary Signal Axis" Animals 11, no. 7: 1915. https://doi.org/10.3390/ani11071915
APA StyleAmevor, F. K., Cui, Z., Du, X., Ning, Z., Shu, G., Jin, N., Deng, X., Tian, Y., Zhang, Z., Kang, X., Xu, D., You, G., Zhang, Y., Li, D., Wang, Y., Zhu, Q., & Zhao, X. (2021). Combination of Quercetin and Vitamin E Supplementation Promotes Yolk Precursor Synthesis and Follicle Development in Aging Breeder Hens via Liver–Blood–Ovary Signal Axis. Animals, 11(7), 1915. https://doi.org/10.3390/ani11071915