
The Impact of Weaning Stress on Gut Health and the Mechanistic Aspects of Several Feed Additives Contributing to Improved Gut Health Function in Weanling Piglets—A Review



Abstract
:Simple Summary
Abstract
1. Introduction
2. Impact of Weaning Stress in Young Pigs
2.1. Gut Health and Intestinal Barrier
2.2. Weaning Transition and Gut Microbiota
2.3. Intestinal Mucosa Immunity and Oxidative Stress
2.4. Feed Intake and Performance during Weaning Transition
3. Nutritional Intervention and Mechanistic Aspects
3.1. Nucleotides
3.2. Phytogenic Compound
3.3. Probiotics
3.4. Prebiotics
3.5. Fatty Acids
3.6. Organic Acids
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Guo, Y.; Wen, Z.; Jiang, X.; Ma, X.; Han, X. Weaning Stress Perturbs Gut Microbiome and Its Metabolic Profile in Piglets. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Spreeuwenberg, M.A.M.; Verdonk, J.M.A.J.; Gaskins, H.R.; Verstegen, M.W.A. Small Intestine Epithelial Barrier Function Is Compromised in Pigs with Low Feed Intake at Weaning. J. Nutr. 2001, 131, 1520–1527. [Google Scholar] [CrossRef] [Green Version]
- Stokes, C.R.; Bailey, M.; Haverson, K.; Harris, C.; Jones, P.; Inman, C.; Pié, S.; Oswald, I.P.; Williams, B.A.; Akkermans, A.D.; et al. Postnatal development of intestinal immune system in piglets: Implications for the process of weaning. Anim. Res. 2004, 53, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Moeser, A.J.; Pohl, C.S.; Rajput, M. Weaning stress and gastrointestinal barrier development: Implications for lifelong gut health in pigs. Anim. Nutr. 2017, 3, 313–321. [Google Scholar] [CrossRef]
- Xiong, X.; Tan, B.; Song, M.; Ji, P.; Kim, K.; Yin, Y.; Liu, Y. Nutritional Intervention for the Intestinal Development and Health of Weaned Pigs. Front. Vet. Sci. 2019, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Jayaraman, B.; Nyachoti, C.M. Husbandry practices and gut health outcomes in weaned piglets: A review. Anim. Nutr. 2017, 3, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Pácha, J. Development of Intestinal Transport Function in Mammals. Physiol. Rev. 2000, 80, 1633–1667. [Google Scholar] [CrossRef] [PubMed]
- Wijtten, P.J.A.; van der Meulen, J.; Verstegen, M.W.A. Intestinal barrier function and absorption in pigs after weaning: A review. Br. J. Nutr. 2011, 105, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R.; Turpin, D.; Kim, J.-C. Gastrointestinal tract (gut) health in the young pig. Anim. Nutr. 2018, 4, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Boudry, G.; Péron, V.; Luron, I.; Lallès, J.-P.; Sève, B. Weaning Induces Both Transient and Long-Lasting Modifications of Absorptive, Secretory, and Barrier Properties of Piglet Intestine. J. Nutr. 2004, 134, 2256–2262. [Google Scholar] [CrossRef] [PubMed]
- Lallès, J.-P.; Boudry, G.; Favier, C.; Le Floc’H, N.; Luron, I.; Montagne, L.; Oswald, I.; Pié, S.; Piel, C.; Sève, B. Gut function and dysfunction in young pigs: Physiology. Anim. Res. 2004, 53, 301–316. [Google Scholar] [CrossRef] [Green Version]
- Lallès, J.-P. Intestinal alkaline phosphatase: Multiple biological roles in maintenance of intestinal homeostasis and modulation by diet. Nutr. Rev. 2010, 68, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Hampson, D. Alterations in piglet small intestinal structure at weaning. Res. Vet. Sci. 1986, 40, 32–40. [Google Scholar] [CrossRef]
- Madara, J.L. Warner-Lambert/Parke-Davis Award lecture. Pathobiology of the intestinal epithelial barrier. Am. J. Pathol. 1990, 137, 1273–1281. [Google Scholar]
- Nabuurs, M.J. Weaning piglets as a model for studying pathophysiology of diarrhea. Vet. Q. 1998, 20, S42–S45. [Google Scholar] [CrossRef] [PubMed]
- Henderson, P.; van Limbergen, J.; Schwarze, J.; Wilson, D.C. Function of the intestinal epithelium and its dysregulation in inflammatory bowel disease. Inflamm. Bowel Dis. 2011, 17, 382–395. [Google Scholar] [CrossRef] [PubMed]
- Moeser, A.J.; Ryan, K.A.; Nighot, P.K.; Blikslager, A. Gastrointestinal dysfunction induced by early weaning is attenuated by delayed weaning and mast cell blockade in pigs. Am. J. Physiol. Liver Physiol. 2007, 293, G413–G421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.F.; Blisklager, A.T.; Moeser, A.J. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohl, C.S.; Medland, J.E.; Moeser, A.J. Early life stress origins of gastrointestinal disease: Animal models, intestinal pathophysiology, and translational implications. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G927–G941. [Google Scholar] [CrossRef] [Green Version]
- Medland, J.E.; Pohl, C.S.; Edwards, L.L.; Frandsen, S.; Bagley, K.; Li, Y.; Moeser, A.J. Early life adversity in piglets induces long-term upregulation of the enteric cholinergic nervous system and heightened, sex-specific secretomotor neuron responses. Neurogastroenterol. Motil. 2016, 28, 1317–1329. [Google Scholar] [CrossRef] [Green Version]
- Isaacson, R.; Kim, H.B. The intestinal microbiome of the pig. Anim. Health Res. Rev. 2012, 13, 100–109. [Google Scholar] [CrossRef]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut microbiota dysbiosis in post weaning piglets: Understanding the keys to health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef]
- Yang, Q.; Huang, X.; Zhao, S.; Sun, W.; Yan, Z.; Wang, P.; Li, S.; Huang, W.; Zhang, S.; Liu, L.; et al. Structure and Function of the Fecal Microbiota in Diarrheic Neonatal Piglets. Front. Microbiol. 2017, 8, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downes, J.; Dewhirst, F.E.; Tanner, A.C.R.; Wade, W.G. Description of Alloprevotella rava gen. nov., sp. nov., isolated from the human oral cavity, and reclassification of Prevotella tannerae Moore et al. 1994 as Alloprevotella tannerae gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 1214–1218. [Google Scholar] [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A Central, Enigmatic Component of the Human Gut Microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef] [PubMed]
- Gophna, U.; Konikoff, T.; Nielsen, H.B. Oscillospira and related bacteria—from metagenomic species to metabolic features. Environ. Microbiol. 2017, 19, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; He, Y.; Irwin, P.L.; Jin, T.; Shi, X. Antibacterial Activity and Mechanism of Action of Zinc Oxide Nanoparticles againstCampylobacter jejuni. Appl. Environ. Microbiol. 2011, 77, 2325–2331. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.B.; Borewicz, K.; White, B.A.; Singer, R.S.; Sreevatsan, S.; Tu, Z.J.; Isaacson, R.E. Microbial shifts in the swine distal gut in response to the treatment with antimicrobial growth promoter, tylosin. Proc. Natl. Acad. Sci. USA 2012, 109, 15485–15490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Nie, Y.; Chen, J.; Zhang, Y.; Wang, Z.; Fan, Q.; Yan, X. Gradual Changes of Gut Microbiota in Weaned Miniature Piglets. Front. Microbiol. 2016, 7, 1727. [Google Scholar] [CrossRef]
- Kim, H.B.; Borewicz, K.; White, B.A.; Singer, R.S.; Sreevatsan, S.; Tu, Z.J.; Isaacson, R.E. Longitudinal investigation of the age-related bacterial diversity in the feces of commercial pigs. Vet. Microbiol. 2011, 153, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Pajarillo, E.A.B.; Chae, J.-P.; Balolong, M.P.; Kim, H.B.; Kang, D.-K. Assessment of fecal bacterial diversity among healthy piglets during the weaning transition. J. Gen. Appl. Microbiol. 2014, 60, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Guevarra, R.B.; Lee, J.H.; Lee, S.H.; Seok, M.-J.; Kim, D.W.; Na Kang, B.; Johnson, T.J.; Isaacson, R.E.; Kim, H.B. Piglet gut microbial shifts early in life: Causes and effects. J. Anim. Sci. Biotechnol. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flint, H.J.; Bayer, E.A. Plant Cell Wall Breakdown by Anaerobic Microorganisms from the Mammalian Digestive Tract. Ann. N. Y. Acad. Sci. 2008, 1125, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Ivarsson, E.; Roos, S.; Liu, H.; Lindberg, J.E. Fermentable non-starch polysaccharides increases the abundance of Bacteroides–Prevotella–Porphyromonas in ileal microbial community of growing pigs. Animal 2014, 8, 1777–1787. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G.; Follador, R. Metabolism of Oligosaccharides and Starch in Lactobacilli: A Review. Front. Microbiol. 2012, 3, 340. [Google Scholar] [CrossRef] [Green Version]
- McCracken, B.A.; Spurlock, M.E.; Roos, M.A.; Zuckermann, F.A.; Gaskins, H.R. Weaning Anorexia May Contribute to Local Inflammation in the Piglet Small Intestine. J. Nutr. 1999, 129, 613–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pié, S.; Lallès, J.-P.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I.P. Weaning Is Associated with an Upregulation of Expression of Inflammatory Cytokines in the Intestine of Piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Ushida, K.; Kameue, C.; Tsukahara, T.; Fukuta, K.; Nakanishi, N. Decreasing Traits of Fecal Immunoglobulin A in Neonatal and Weaning Piglets. J. Vet. Med Sci. 2008, 70, 849–852. [Google Scholar] [CrossRef] [Green Version]
- Ponta, H.; Cato, A.; Herrlich, P. Interference of pathway specific transcription factors. Biochim. Biophys. Acta Gene Struct. Expr. 1992, 1129, 255–261. [Google Scholar] [CrossRef]
- Rao, R. Oxidative stress-induced disruption of epithelial and endothelial tight junctions. Front. Biosci. 2008, 13, 7210–7226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergauwen, H.; Tambuyzer, B.; Jennes, K.; DeGroote, J.; Wang, W.; de Smet, S.; Michiels, J.; Van Ginneken, C. Trolox and Ascorbic Acid Reduce Direct and Indirect Oxidative Stress in the IPEC-J2 Cells, an In Vitro Model for the Porcine Gastrointestinal Tract. PLoS ONE 2015, 10, e0120485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Dividich, J.L.; Sève, B. Effects of underfeeding during the weaning period on growth, metabolism, and hormonal adjustments in the piglet. Domest. Anim. Endocrinol. 2000, 19, 63–74. [Google Scholar] [CrossRef]
- Colson, V.; Orgeur, P.; Foury, A.; Mormède, P. Consequences of weaning piglets at 21 and 28 days on growth, behaviour and hormonal responses. Appl. Anim. Behav. Sci. 2006, 98, 70–88. [Google Scholar] [CrossRef]
- Heo, J.M.; Opapeju, F.O.; Pluske, J.R.; Kim, J.C.; Hampson, D.J.; Nyachoti, C.M. Gastrointestinal health and function in weaned pigs: A review of feeding strategies to control post-weaning diarrhea without using in-feed antimicrobial compounds. J. Anim. Physiol. Anim. Nutr. 2013, 97, 207–237. [Google Scholar] [CrossRef]
- Kats, L.J.; Tokach, M.D.; Goodband, R.D.; Nelssen, J.L. Influence of weaning weight and growth during the first week postweaning on subsequent pig performance. Kans. Agric. Exp. Stn. Res. Rep. 1992, 15–17. [Google Scholar] [CrossRef] [Green Version]
- Lallès, J.; Bosi, P.; Janczyk, P.; Koopmans, S.; Torrallardona, D. Impact of bioactive substances on the gastrointestinal tract and performance of weaned piglets: A review. Animal 2009, 3, 1625–1643. [Google Scholar] [CrossRef] [Green Version]
- Pluske, J.R. Feed and feed additives-related aspects of gut health and development in weanling pigs. J. Anim. Sci. Biotechnol. 2013, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Espinosa, C.D.; Abelilla, J.J.; Casas, G.A.; Lagos, L.V.; Lee, S.A.; Kwon, W.B.; Mathai, J.K.; Navarro, D.M.D.; Jaworski, N.W.; et al. Non-antibiotic feed additives in diets for pigs: A review. Anim. Nutr. 2018, 4, 113–125. [Google Scholar] [CrossRef]
- Ziegler, A.; Gonzalez, L.; Blikslager, A. Large Animal Models: The Key to Translational Discovery in Digestive Disease Research. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 716–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, X.; Hua, X.; Yang, Q.; Ding, D.; Che, C.; Cui, L.; Jia, W.; Bucheli, P.; Zhao, L. Inter-species transplantation of gut microbiota from human to pigs. ISME J. 2007, 1, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Hanczakowska, E.; Szewczyk, A.; Okoń, K. Effects of dietary caprylic and capric acids on piglet performance and mucosal epithelium structure of the ileum. J. Anim. Feed. Sci. 2011, 20, 556–565. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Hu, J.; Kim, I.H. Supplemental effects of coated omega-3 polyunsaturated fatty acids to basal diet on improving growth performance, nutrients digestibility, fecal lactobacillus count and fecal score in weaning pigs. Anim. Feed. Sci. Technol. 2020, 269, 114547. [Google Scholar] [CrossRef]
- Jang, K.B.; Kim, S.W. Supplemental effects of dietary nucleotides on intestinal health and growth performance of newly weaned pigs. J. Anim. Sci. 2019, 97, 4875–4882. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Xie, C.; Wang, Q.; Wan, D.; Zhang, Y.; Wu, X.; Yin, Y. Uridine/UMP metabolism and their function on the gut in segregated early weaned piglets. Food Funct. 2019, 10, 4081–4089. [Google Scholar] [CrossRef]
- Perricone, V.; Comi, M.; Bontempo, V.; Lecchi, C.; Ceciliani, F.; Crestani, M.; Ferrari, A.; Savoini, G.; Agazzi, A. Effects of nucleotides administration on growth performance and immune response of post-weaning piglets. Ital. J. Anim. Sci. 2020, 19, 295–301. [Google Scholar] [CrossRef]
- Tsiloyiannis, V.; Kyriakis, S.; Vlemmas, J.; Sarris, K. The effect of organic acids on the control of porcine post-weaning diarrhoea. Res. Vet. Sci. 2001, 70, 287–293. [Google Scholar] [CrossRef]
- Luise, D.; Motta, V.; Salvarani, C.; Chiappelli, M.; Fusco, L.; Bertocchi, M.; Mazzoni, M.; Maiorano, G.; Costa, L.N.; van Milgen, J.; et al. Long-term administration of formic acid to weaners: Influence on intestinal microbiota, immunity parameters and growth performance. Anim. Feed. Sci. Technol. 2017, 232, 160–168. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Lee, K.Y.; Serpunja, S.; Song, T.H.; Kim, I.H. Growth performance, nutrient digestibility, fecal microbiota and fecal noxious gas emission in weaning pigs fed high and low density diet with and without protected organic acid blends. Anim. Feed. Sci. Technol. 2018, 239, 1–8. [Google Scholar] [CrossRef]
- Long, S.; Xu, Y.; Pan, L.; Wang, Q.; Wang, C.; Wu, J.; Wu, Y.; Han, Y.; Yun, C.; Piao, X. Mixed organic acids as antibiotic substitutes improve performance, serum immunity, intestinal morphology and microbiota for weaned piglets. Anim. Feed. Sci. Technol. 2018, 235, 23–32. [Google Scholar] [CrossRef]
- Yang, Y.; Lee, K.Y.; Kim, I. Effects of dietary protected organic acids on growth performance, nutrient digestibility, fecal microflora, diarrhea score, and fecal gas emission in weanling pigs. Can. J. Anim. Sci. 2019, 99, 514–520. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Chen, X.; Ji, C.; Ma, Q.; Hao, K. Study of the application of fructo-oligosaccharides in piglets. Asian-Australas. J. Anim. Sci. 2005, 18, 1011–1016. [Google Scholar] [CrossRef]
- Bhandari, S.K.; Nyachoti, C.M.; Krause, D.O. Raw potato starch in weaned pig diets and its influence on post-weaning scours and the molecular microbial ecology of the digestive tract. J. Anim. Sci. 2009, 87, 984–993. [Google Scholar] [CrossRef]
- Patterson, J.K.; Yasuda, K.; Welch, R.M.; Miller, D.D.; Lei, X.G. Supplemental Dietary Inulin of Variable Chain Lengths Alters Intestinal Bacterial Populations in Young Pigs. J. Nutr. 2010, 140, 2158–2161. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Ordaz, A.A.; González-Ortiz, G.; La Ragione, R.M.; Woodward, M.J.; Collins, J.W.; Pérez, J.F.; Martín-Orúe, S.M. Lactulose and Lactobacillus plantarum, a Potential Complementary Synbiotic To Control Postweaning Colibacillosis in Piglets. Appl. Environ. Microbiol. 2014, 80, 4879–4886. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.J.; Kim, Y.M.; Park, J.H.; Baek, D.H.; Nyachoti, C.M.; Kim, I.H. Effects of levan-type fructan on growth performance, nutrient digestibility, diarrhoea scores, faecal shedding of total lactic acid bacteria and coliform bacteria, and faecal gas emission in weaning pigs. J. Sci. Food Agric. 2017, 98, 1539–1544. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.-Y.; Li, K.-N.; Xu, Y.-Q.; Wu, Y.-Z.; Shi, L.-L.; Guo, S.-W.; Yan, S.-M.; Jin, X.; Shi, B.-L. Effects of galacto-oligosaccharide on growth performance, feacal microbiota, immune response and antioxidant capability in weaned piglets. J. Appl. Anim. Res. 2020, 48, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Van der Peet-Schwering, C.M.C.; Jansman, A.J.M.; Smidt, H.; Yoon, I. Effects of yeast culture on performance, gut integrity, and blood cell composition of weanling pigs 1,2. J. Anim. Sci. 2007, 85, 3099–3109. [Google Scholar] [CrossRef]
- Konstantinov, S.R.; Smidt, H.; Akkermans, A.D.L.; Casini, L.; Treisi, P.; Mazzoni, M.; Filippi, S.D.; Bosi, P.; De vos, W.M. Feeding of Lactobacillus sobrius reduces Escherichia coli F4 levels in the gut and promotes growth of infected piglets. FEMS Microbiol. Ecol. 2008, 66, 599–607. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.B.; Piao, X.S.; Kim, S.W.; Wang, L.; Liu, P.; Yoon, I.; Zhen, Y.G. Effects of yeast culture supplementation on growth performance, intestinal health, and immune response of nursery pigs1. J. Anim. Sci. 2009, 87, 2614–2624. [Google Scholar] [CrossRef]
- Chiang, M.-L.; Chen, H.-C.; Chen, K.-N.; Lin, Y.-C.; Lin, Y.-T.; Chen, M.-J. Optimizing Production of Two Potential Probiotic Lactobacilli Strains Isolated from Piglet Feces as Feed Additives for Weaned Piglets. Asian-Australas. J. Anim. Sci. 2015, 28, 1163–1170. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Dun, Y.; Li, S.; Zhang, D.; Peng, N.; Zhao, S.; Liang, Y. Dietary Enterococcus faecalis LAB31 Improves Growth Performance, Reduces Diarrhea, and Increases Fecal Lactobacillus Number of Weaned Piglets. PLoS ONE 2015, 10, e0116635. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Kim, K.P.; Kim, I.H. Influence of Bacillus subtilis GCB-13-001 on growth performance, nutrient digestibility, blood characteristics, faecal microbiota and faecal score in weanling pigs. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1919–1925. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.H.; Nyachoti, C.M.; Kim, I.H. Evaluation of effect of probiotics mixture supplementation on growth performance, nutrient digestibility, faecal bacterial enumeration, and noxious gas emission in weaning pigs. Ital. J. Anim. Sci. 2018, 18, 466–473. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Jinno, C.; Kim, K.; Wu, Z.; Tan, B.; Li, X.; Whelan, R.; Liu, Y. Dietary Bacillus spp. enhanced growth and disease resistance of weaned pigs by modulating intestinal microbiota and systemic immunity. J. Anim. Sci. Biotechnol. 2020, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Meng, Q.W.; Kim, I.H. Effect of an herb extract mixture on growth performance, nutrient digestibility, blood characteristics, and fecal microbial shedding in weanling pigs. Livest. Sci. 2012, 145, 189–195. [Google Scholar] [CrossRef]
- Bontempo, V.; Jiang, X.-R.; Cheli, F.; Verso, L.L.; Mantovani, G.; Vitari, F.; Domeneghini, C.; Agazzi, A. Administration of a novel plant extract product via drinking water to post-weaning piglets: Effects on performance and gut health. Animal 2014, 8, 721–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.I.; Kim, I.H. Eckol Alleviates Intestinal Dysfunction during Suckling-to-Weaning Transition via Modulation of PDX1 and HBEGF. Int. J. Mol. Sci. 2020, 21, 4755. [Google Scholar] [CrossRef]
- Cosgrove, M. Perinatal and infant nutrition. Nucleotides. Nutrition 1998, 14, 748–751. [Google Scholar] [CrossRef]
- Sauer, N.; Mosenthin, R.; Bauer, E. The role of dietary nucleotides in single-stomached animals. Nutr. Res. Rev. 2011, 24, 46–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, L.; Hu, L.; Liu, Y.; Yan, C.; Peng, X.; Xu, Q.; Wang, R.; Cheng, Y.; Chen, H.; Fang, Z.; et al. Dietary Nucleotides Supplementation Improves the Intestinal Development and Immune Function of Neonates with Intra-Uterine Growth Restriction in a Pig Model. PLoS ONE 2016, 11, e0157314. [Google Scholar] [CrossRef] [PubMed]
- Stein, H.H.; Mateo, C.D. Nucleotides may have a role in nutrition of young pigs. Feedstuffs 2004, 76, 11–14. [Google Scholar]
- Superchi, P.; Saleri, R.; Borghetti, P.; de Angelis, E.; Ferrari, L.; Cavalli, V.; Amicucci, P.; Ossiprandi, M.C.; Sabbioni, A. Effects of dietary nucleotide supplementation on growth performance and hormonal and immune responses of piglets. Animal 2012, 6, 902–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Zhao, P.; Lei, Y.; Li, T.; Kim, I. Response to an Escherichia coli K88 oral challenge and productivity of weanling pigs receiving a dietary nucleotides supplement. J. Anim. Sci. Biotechnol. 2015, 6, 49. [Google Scholar] [CrossRef]
- Waititu, S.M.; Yin, F.; Patterson, R.; Yitbarek, A.; Rodriguez-Lecompte, J.C.; Nyachoti, C.M. Dietary supplementation with a nucleotide-rich yeast extract modulates gut immune response and microflora in weaned pigs in response to a sanitary challenge. Animal 2017, 11, 2156–2164. [Google Scholar] [CrossRef]
- Kulkarni, A.D.; Rudolph, F.B.; van Buren, C.T. The Role of Dietary Sources of Nucleotides in Immune Function: A Review. J. Nutr. 1994, 124, 1442S–1446S. [Google Scholar] [CrossRef]
- Mateo, C.D.; Peters, D.N.; Stein, H.H. Nucleotides in sow colostrum and milk at different stages of lactation 1,2,3. J. Anim. Sci. 2004, 82, 1339–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.I.; Kim, I.H. Nucleotide-mediated SPDEF modulates TFF3-mediated wound healing and intestinal barrier function during the weaning process. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Sauer, N.; Eklund, M.; Bauer, E.; Gänzle, M.; Field, C.; Zijlstra, R.T.; Mosenthin, R. The effects of pure nucleotides on performance, humoral immunity, gut structure and numbers of intestinal bacteria of newly weaned pigs1. J. Anim. Sci. 2012, 90, 3126–3134. [Google Scholar] [CrossRef] [PubMed]
- Upadhaya, S.D.; Kim, I.H. Efficacy of phytogenic feed additive on performance, production and health status of monogastric animals—a review. Ann. Anim. Sci. 2017, 17, 929–948. [Google Scholar] [CrossRef] [Green Version]
- Lillehoj, H.; Liu, Y.; Calsamiglia, S.; Fernandez-Miyakawa, M.E.; Chi, F.; Cravens, R.L.; Oh, S.; Gay, C.G. Phytochemicals as potential antibiotic alternatives to promote growth and enhance host health: A report from the second international symposium on alternatives to antibiotics. Vet. Res. 2018, 46, 76–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Song, M.; Che, T.M.; Almeida, J.A.S.; Lee, J.J.; Bravo, D.; Maddox, C.W.; Pettigrew, J.E. Dietary plant extracts alleviate diarrhea and alter immune responses of weaned pigs experimentally infected with a pathogenic Escherichia coli1. J. Anim. Sci. 2013, 91, 5294–5306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Xiang, Q.; Wang, J.; Peng, J.; Wei, H. Oregano Essential Oil Improves Intestinal Morphology and Expression of Tight Junction Proteins Associated with Modulation of Selected Intestinal Bacteria and Immune Status in a Pig Model. Bio. Med. Res. Int. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yuan, D.; Hussain, T.; Tan, B.; Liu, Y.; Ji, P.; Yin, Y. The Evaluation of Antioxidant and Anti-Inflammatory Effects ofEucommia ulmoidesFlavones Using Diquat-Challenged Piglet Models. Oxidative Med. Cell. Longev. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Hosseindoust, A.; Goel, A.; Lee, S.; Jha, P.K.; Kwon, I.K.; Chae, B.-J. Effects of Ecklonia cava as fucoidan-rich algae on growth performance, nutrient digestibility, intestinal morphology and caecal microflora in weanling pigs. Asian-Australas. J. Anim. Sci. 2016, 30, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boirivant, M.; Strober, W. The mechanism of action of probiotics. Curr. Opin. Gastroenterol. 2007, 23, 679–692. [Google Scholar] [CrossRef]
- De Vrese, M.; Schrezenmeir, J. Probiotics, prebiotics, synbiotics. Adv. Biochem. Eng. Biotechnol. 2008, 111, 1–66. [Google Scholar]
- Upadhaya, S.D.; Shanmugam, S.K.; Kang, D.K.; Kim, I.H. Preliminary assessment on potentials of probiotic B. subtilis RX7 and B. methylotrophicus C14 strains as an immune modulator in Salmonella-challenged weaned pigs. Trop. Anim. Health Prod. 2017, 49, 1065–1070. [Google Scholar] [CrossRef]
- Ohland, C.L.; Macnaughton, W.K. Probiotic bacteria and intestinal epithelial barrier function. Am. J. Physiol. Liver Physiol. 2010, 298, G807–G819. [Google Scholar] [CrossRef] [Green Version]
- Rose, E.; Odle, J.; Blikslager, A.; Ziegler, A. Probiotics, Prebiotics and Epithelial Tight Junctions: A Promising Approach to Modulate Intestinal Barrier Function. Int. J. Mol. Sci. 2021, 22, 6729. [Google Scholar] [CrossRef] [PubMed]
- Huan-Long, Q.; Shen, T.Y.; Gao, Z.G.; Fan, X.B.; Hang, X.M.; Jiang, Y.Q.; Zhang, H.Z. Effect of lactobacillus on the gut microflora and barrier function of the rats with abdominal infection. World J. Gastroenterol. 2005, 11, 2591. [Google Scholar]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; Park, Z.; McCann, M.J.; Kelly, W.J.; Roy, N.C. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 2010, 10, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.-J.M.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwood, B.P.; Yuan, C.Y.; Wood, D.R.; Nicolas, J.D.; Grothaus, J.S.; Hunter, C.J. Probiotic Lactobacillus Species Strengthen Intestinal Barrier Function and Tight Junction Integrity in Experimental Necrotizing Enterocolitis. J. Probiotics Health 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, K.R.; Liu, S.X.; Tian, R.; Kushnir, A.; Turner, J.R.; Li, H.-L.; Chou, P.M.; Weber, C.; De Plaen, I.G. Bifidobacteria Stabilize Claudins at Tight Junctions and Prevent Intestinal Barrier Dysfunction in Mouse Necrotizing Enterocolitis. Am. J. Pathol. 2013, 182, 1595–1606. [Google Scholar] [CrossRef] [Green Version]
- Mattar, A.F.; Teitelbaum, D.H.; Drongowski, R.A.; Yongyi, F.; Harmon, C.M.; Coran, A.G. Probiotics up-regulate MUC-2 mucin gene expression in a Caco-2 cell-culture model. Pediatr. Surg. Int. 2002, 18, 586–590. [Google Scholar]
- Mack, D.R.; Ahrne, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extracellular MUC3 mucin secretion follows adherence of Lactobacillus strains to intestinal epithelial cells in vitro. Gut 2003, 52, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Shanahan, F. A commentary on the safety of probiotics. Gastroenterol. Clin. North Am. 2012, 41, 869–876. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Chen, Q.; Ren, Y.; Lu, J.; Bartlett, M.; Chen, L.; Zhang, Y.; Guo, X.; Liu, C. A Novel Prebiotic Blend Product Prevents Irritable Bowel Syndrome in Mice by Improving Gut Microbiota and Modulating Immune Response. Nutrients 2017, 9, 1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Tian, S.; Yu, H.; Wang, J.; Zhu, W. Response of colonic mucosa-associated microbiota composition, mucosal immune homeostasis, and barrier function to early life galacto-oligosaccharides intervention in suckling piglets. J. Agric. Food Chem. 2019, 67, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Sun, W.; Pei, X.; Jin, Y.; Wang, H.; Tao, W.; Xiao, Z.; Liu, L.; Wang, M. Galactooligosaccharide pretreatment alleviates damage of the intestinal barrier and inflammatory responses in LPS-challenged mice. Food Funct. 2021, 12, 1569–1579. [Google Scholar] [CrossRef] [PubMed]
- Uerlings, J.; Schroyen, M.; Willems, E.; Tanghe, S.; Bruggeman, G.; Bindelle, J.; Everaert, N. Differential effects of inulin or its fermentation metabolites on gut barrier and immune function of porcine intestinal epithelial cells. J. Funct. Foods 2020, 67, 103855. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Wongkrasant, P.; Pongkorpsakol, P.; Ariyadamrongkwan, J.; Meesomboon, R.; Satitsri, S.; Pichyangkura, R.; Barrett, K.E.; Muanprasat, C. A prebiotic fructo-oligosaccharide promotes tight junction assembly in intestinal epithelial cells via an AMPK-dependent pathway. Biomed. Pharmacother. 2020, 129, 110415. [Google Scholar] [CrossRef]
- Muanprasat, C.; Wongkrasant, P.; Satitsri, S.; Moonwiriyakit, A.; Pongkorpsakol, P.; Mattaveewong, T.; Pichyangkura, R.; Chatsudthipong, V. Activation of AMPK by chitosan oligosaccharide in intestinal epithelial cells: Mechanism of action and potential applications in intestinal disorders. Biochem. Pharmacol. 2015, 96, 225–236. [Google Scholar] [CrossRef]
- Biagi, G.; Piva, A.; Moschini, M.; Vezzali, E.; Roth, F.X. Performance, intestinal microflora, and wall morphology of weanling pigs fed sodium butyrate1. J. Anim. Sci. 2007, 85, 1184–1191. [Google Scholar] [CrossRef] [Green Version]
- Mazzoni, M.; Gall, M.L.; de Filippi, S.; Minieri, L.; Trevisi, P.; Woliński, J.; Lalatta-Costerbosa, G.; Lallès, J.-P.; Guilloteau, P.; Bosi, P. Supplemental Sodium Butyrate Stimulates Different Gastric Cells in Weaned Pigs. J. Nutr. 2008, 138, 1426–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gall, M.L.; Gallois, M.; Sève, B.; Louveau, I.; Holst, J.J.; Oswald, I.; Lallès, J.-P.; Guilloteau, P. Comparative effect of orally administered sodium butyrate before or after weaning on growth and several indices of gastrointestinal biology of piglets. Br. J. Nutr. 2009, 102, 1285–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhaya, S.D.; Jiao, Y.; Kim, Y.M.; Lee, K.Y.; Kim, I.H.; Yang, J. Coated sodium butyrate supplementation to a reduced nutrient diet enhanced the performance and positively impacted villus height and faecal and digesta bacterial composition in weaner pigs. Anim. Feed. Sci. Technol. 2020, 265, 114534. [Google Scholar] [CrossRef]
- Deschepper, K.; Gantois, I.; Maertens, L.; van Meenen, E. A balanced mixture of medium chain fatty acids improves zootechnical performances and slaughter results of broilers. In Proceedings of the International Poultry Scientific Forum, Atlanta, GA, USA, 28–29 January 2013; p. 22. [Google Scholar]
- Jackman, J.A.; Boyd, R.D.; Elrod, C.C. Medium-chain fatty acids and monoglycerides as feed additives for pig production: Towards gut health improvement and feed pathogen mitigation. J. Anim. Sci. Biotechnol. 2020, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.-J. Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [Green Version]
- Skřivanová, E.; Marounek, M.; Benda, V.; Březina, P. Susceptibility of Escherichia coli, Salmonella sp and Clostridium perfringens to organic acids and monolaurin. Vet. Med. 2006, 51, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.M.; Hwang, J.H.; Kim, I.H. Effect of Medium-chain Triglyceride (MCT) on Growth Performance, Nutrient Digestibility, Blood Characteristics in Weanling Pigs. Asian-Australas. J. Anim. Sci. 2012, 25, 1003–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, H.; Yang, L.; Zhang, L.; Wang, T. Effect of medium-chain triglycerides on growth performance, nutrient digestibility, plasma metabolites and antioxidant capacity in weanling pigs. Anim. Nutr. 2015, 1, 12–18. [Google Scholar] [CrossRef]
- Lee, S.I.; Kang, K.S. Function of capric acid in cyclophosphamide-induced intestinal inflammation, oxidative stress, and barrier function in pigs. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Wang, J.; Huang, N.; Xiong, J.; Wei, H.; Jiang, S.; Peng, J. Caprylic acid and nonanoic acid upregulate endogenous host defense peptides to enhance intestinal epithelial immunological barrier function via histone deacetylase inhibition. Int. Immunopharmacol. 2018, 65, 303–311. [Google Scholar] [CrossRef]
- Zhou, Z.; Huang, J.; Hao, H.; Wei, H.; Zhou, Y.; Peng, J. Applications of new functions for inducing host defense peptides and synergy sterilization of medium chain fatty acids in substituting in-feed antibiotics. J. Funct. Foods 2018, 52, 348–359. [Google Scholar] [CrossRef]
- Desbois, A.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2009, 85, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- Desbois, A.P. Potential Applications of Antimicrobial Fatty Acids in Medicine, Agriculture and Other Industries. Recent Pat. Anti-Infect. Drug Discov. 2012, 7, 111–122. [Google Scholar] [CrossRef]
- Desbois, A.P.; Lawlor, K.C. Antibacterial Activity of Long-Chain Polyunsaturated Fatty Acids against Propionibacterium acnes and Staphylococcus aureus. Mar. Drugs 2013, 11, 4544–4557. [Google Scholar] [CrossRef] [Green Version]
- Wieckowski, M.; Wojtczak, L. Fatty acid-induced uncoupling of oxidative phosphorylation is partly due to opening of the mitochondrial permeability transition pore. FEBS Lett. 1998, 423, 339–342. [Google Scholar] [CrossRef]
- Shaikh, S.R.; Jolly, C.A.; Chapkin, R. n−3 Polyunsaturated fatty acids exert immunomodulatory effects on lymphocytes by targeting plasma membrane molecular organization. Mol. Asp. Med. 2012, 33, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Holman, D.B.; Baurhoo, B.; Chénier, M.R. Temporal analysis of the effect of extruded flaxseed on the swine gut microbiota. Can. J. Microbiol. 2014, 60, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Upadhaya, S.D.; Kim, I.H. Supplemental effects of fish oil and powdered/coated docosahexaenoic acid on the growth performance, nutrient digestibility, blood profile and fecal coliform and lactic acid bacteria counts in weaner pigs. Anim. Feed. Sci. Technol. 2021, 275, 114885. [Google Scholar] [CrossRef]
- Thaela, M.-J.; Jensen, M.S.; Pierzynowski, S.G.; Jakob, S. Effect of lactic acid supplementation on pancreatic secretion in pigs after weaning. J. Anim. Feed. Sci. 1998, 7, 181–183. [Google Scholar] [CrossRef] [Green Version]
- Risley, C.R.; Kornegay, E.T.; Lindemann, M.D.; Wood, C.M.; Eigel, W.N. Effect of feeding organic acids on gastrointestinal digesta measurements at various times postweaning in pigs challenged with enterotoxigenic Escherichia coli. Can. J. Anim. Sci. 1993, 73, 931–940. [Google Scholar] [CrossRef]
- Risley, C.; Kornegay, E.; Lindemann, M.; Weakland, S. Effects of organic acids with and without a microbial culture on performance and gastrointestinal tract measurements of weanling pigs. Anim. Feed. Sci. Technol. 1991, 35, 259–270. [Google Scholar] [CrossRef]
- Zentek, J.; Ferrara, F.; Pieper, R.; Tedin, L.; Meyer, W.; Vahjen, W. Effects of dietary combinations of organic acids and medium chain fatty acids on the gastrointestinal microbial ecology and bacterial metabolites in the digestive tract of weaning piglets. J. Anim. Sci. 2013, 91, 3200–3210. [Google Scholar] [CrossRef]
- Ferrara, F.; Tedin, L.; Pieper, R.; Meyer, W.; Zentek, J. Influence of medium-chain fatty acids and short-chain organic acids on jejunal morphology and intra-epithelial immune cells in weaned piglets. J. Anim. Physiol. Anim. Nutr. 2016, 101, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.T.; Hwang, J.A.; Hoon, J.; Mun, H.S.; Yang, C.J. Comparison of single and blend of acidifiers as an alternative to antibiotics on growth performance, fecal microflora and humoral immunity in weaned piglets. Asian-Australas. J. Anim. Sci. 2014, 27, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Popiolek, L.; Biernasiuk, A. Design, synthesis, and in vitro antimicrobial activity of hydrazide-hydrazones of 2-substituted acetic acid. Chem. Biol. Drug Des. 2016, 88, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Warnecke, T.; Gill, R.T. Organic acid toxicity, tolerance, and production in Escherichia coli biorefining applications. Microb. Cell Factories 2005, 4, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, H.; Leonard, F.; Walia, K.; Lawlor, P.; Duffy, G.; Fanning, S.; Markey, B.; Brady, C.; Gardiner, G.; Argüello, H. Investigation of in-feed organic acids as a low cost strategy to combat Salmonella in grower pigs. Prev. Vet. Med. 2017, 139, 50–57. [Google Scholar] [CrossRef]
- Walia, K.; Argüello, H.; Lynch, H.; Leonard, F.C.; Grant, J.; Yearsley, D.; Kelly, S.; Duffy, G.; Gardiner, G.E.; Lawlor, P.G. Effect of strategic administration of an encapsulated blend of formic acid, citric acid, and essential oils on Salmonella carriage, seroprevalence, and growth of finishing pigs. Prev. Vet. Med. 2017, 137, 28–35. [Google Scholar] [CrossRef]
- Koyuncu, S.; Andersson, M.G.; Löfström, C.; Skandamis, P.N.; Gounadaki, A.; Zentek, J.; Häggblom, P. Organic acids for control of Salmonella in different feed materials. BMC Vet. Res. 2013, 9, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Additive Type | Composition | Age of Piglets/Experiment Duration | Dose | Intestinal Structure, Gut/Health Microbiota | Performance/Other Observations | References |
|---|---|---|---|---|---|---|
| Fatty acid (FA) | ||||||
| Medium-chain fatty acid | Caprylic or capric acid | 32 days old/49-d trial | 2 g/kg | Reduced Clostridium perfringens levels;improved mucosal epithelium structure of ileum | Improved overall ADG. FCR in pigs receiving diet supplemented with caprylic acid was better; increased digestibility and reduced mortality | [53] |
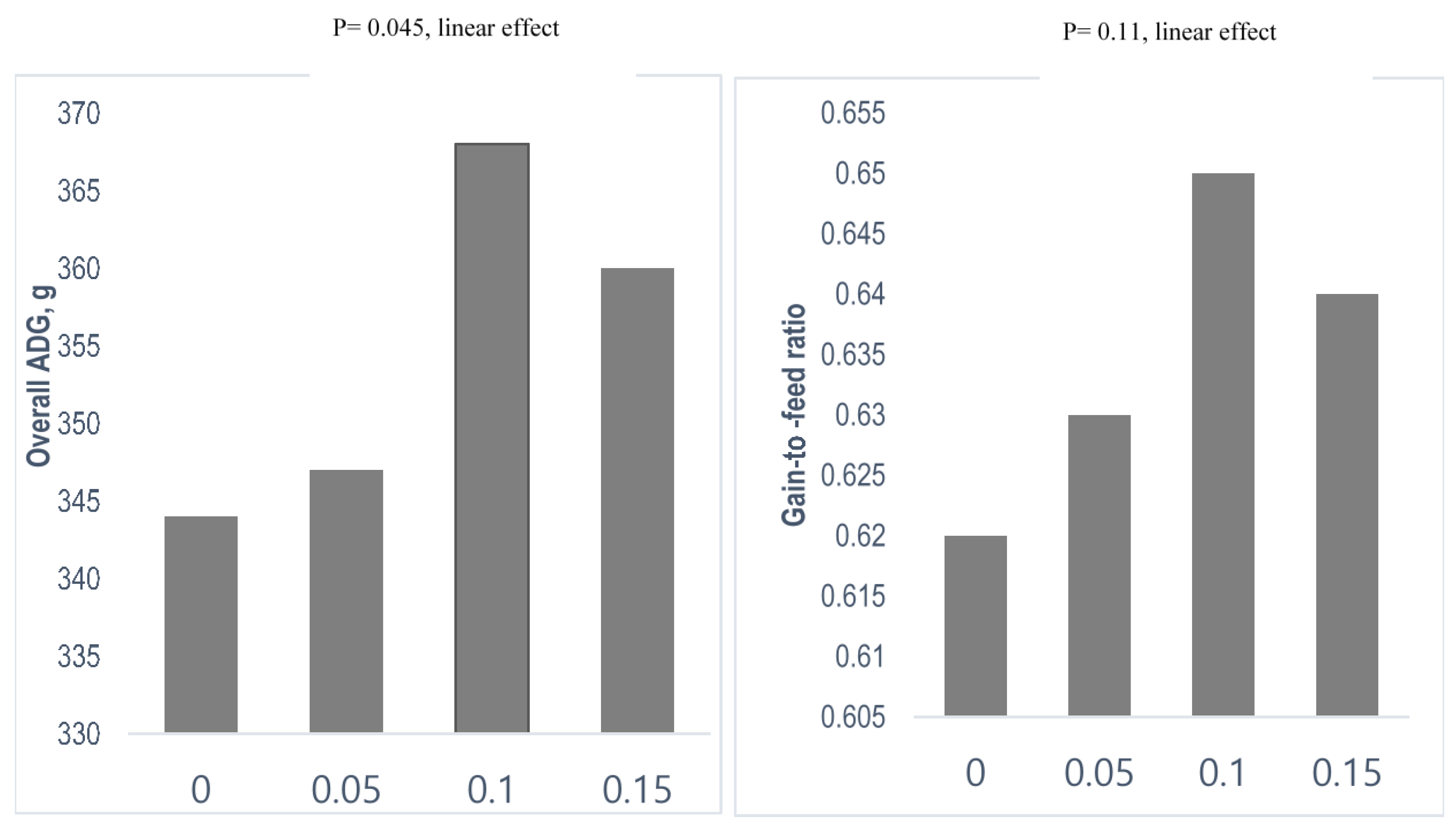
| Poly unsaturated fatty acids | Coated n-3 FA | 28 days old/42-d trial | 5, 10 and 15 g/kg. | Trends in linear increment in fecal Lactobacillus counts at weeks 3 and 6 | Linear increase in ADG during week 1, 3 and overall, G:F linearly increased during overall; linear increase in DM and N digestibility at the end of experiment | [54] |
| Nucleotides | ||||||
| UMP, GMP, AMP, CMP, and IMP | 19 days old/21-d trial | 0, 50, 150, 250, and 500 mg/kg | Quadratic response on villus height–crypt ratio, linear reduction in crypt cell proliferation in jejunum, reduced jejunal IL-6 when nucleotide increased from 50 to 150 mg/kg | Increased ADG during the first 11 days when pigs received diet supplemented with 50–150 mg/kg nucleotide | [55] | |
| UMP/UR | 12 days old/10-d trail | 476 mg UP, 348 mg UR(orally) | Decreased the diarrhea rate, UR increased the jejunum villus length/crypt depth ratio, Claudin-3, and E-cadherin expression, and the pyrimidine nucleotide metabolic enzymes in the duodenal mucosa, UMP and UR decreased the expression of CAD and RRM2 at the jejunal mucosa | UMP and UR supplements improved the ADG of piglets | [56] | |
| UMP, GMP, AMP, CMP, and IMP | 28 days old/28-d trial | 0.8 g/head/day | No differences in gene expression levels of inflammatory cytokines (IL1α, IL1β, IL6, IL10, TNFα, TLR2, TLR4, and PPAR) at ileal Peyer’s patches level; no effect on IgA and IgG content in blood | Increased BW and ADG but not G:F | [57] | |
| Organic acid (OA) | ||||||
| Pure OA (propionic acid, lactic acid, formic acid, malic acid, citric acid, or fumaric acid) | 25 days old/28-d trial | 10, 16, 12, 12, 15, and 15 g/kg, respectively | Reduced incidence and severity of diarrhea | Heavier BW, and increased ADG and FI especially with lactic acid supplementation | [58] | |
| Formic acid | 35 days old/42-d trial | 0, 1.4 g/kg (low formic acid; LFA), or 6.4 g/kg (high formic acid; HFA) | Increased microbiota diversity in high FA group | Increased ADG, ADFI and improved feed efficiency during the first three weeks in both high and low OA group | [59] | |
| 17% fumaric acid, 13% citric acid, 10% malic acid, and 1.2% medium-chain fatty acid (protected OA) | 28 days old/42-d trial | 1 and 2 g/kg | Reduction in E. coli counts at week 3 and increase in Lactobacillus counts in week 6 with 2 g/kg organic acid | Increased overall ADG and ADFI | [60] | |
| Formic acid, acetic acid, and propionic acid combined with medium chain fatty acids | 28-d trial | 3 g/kg | Reduced the incidence of diarrhea and fecal E. coli counts, the ratio of villus height-to-crypt depth in the jejunum and ileum was higher | Digestibility of dry matter, total carbohydrates, NDF, and ADF was increased during days 14–28 | [61] | |
| 17% fumaric acid, 13% citric acid, 10% malic acid, and 1.2% medium-chain fatty acid | 28 days old/42-d trial | 0, 1, and 2 g/kg | Linear increase in fecal Lactobacillus counts and linear reduction in E. coli and Salmonella counts | Increase in overall ADG and DM digestibility | [62] | |
| Prebiotics | ||||||
| Fructo-oligosaccharide | 33 days old/21-d trial | 4 g/kg | Increased villus height, reduced diarrhea | Improved ADG, increased the concentrations of isobutyric and butyric acid and total VFAs in the caecum, and acetic acid, isovaleric acid, and total VFAs in feces | [63] | |
| Resistant starch | 17 days old/21-d trial | 70 and 140 g/kg | Enhanced microbial diversity in colon and reduced diarrhea with 7% resistant starch inclusion in the diet | No effect on growth performance | [64] | |
| Inulin | 42 day old/35-d trial | 40 mg/kg | Increased Lactobacilli and Bifidobacteria and reduced Enterobacteriaceae and Clostridium spp. in the lumen and mucosa of gut | Higher blood hemoglobin | [65] | |
| Lactulose | 25 days old/18-d trail | 10 g/kg | Increased Lactobacilli and the percentage of butyric acid in the colon; an increase in the ileum villous height | Improved the ADG; reduction of the pig major acute-phase protein in serum | [66] | |
| Levan-type fructan | 28 days old/42-d trial | 0, 0.1, 0.5, and 1.0 g/kg | Linear increase in fecal lactic acid bacteria counts with the increase in the dose of levan | Increased ADG and ADFI linearly during days 0–21 and overall; linear increase in the digestibility of DM, CP, and GE | [67] | |
| Galacto oligosaccharide (GOS) | 28 days old/28-d trial | 0, 500, 1000, 1500, and 2000 mg/kg | Increased the number of Lactobacillus and Bifidobacterium, and decreased the number of E. coli in a linear or quadratic dose-dependent manner; decreased serum concentration of pro-inflammatory cytokines but increased anti-inflammatory cytokines in a linear or quadratic dose-dependent manner | Promoted the growth and activities of antioxidant enzyme in a linear or quadratic dose-dependent manner | [68] | |
| Probiotics | ||||||
| Saccharomyces cerevisiae | 27 days old/35-d trial | 1.25 g/kg | Villus length and crypt depth not affected by probiotic but were greater at 5 weeks vs. 2 weeks after weaning; CD4 and CD8 cells were lower at 5 week after weaning | Improved ADG and G:F | [69] | |
| Lactobacillus sobrius | 21 days old (challenged with 1.5 mL suspension of 1010 CFU ETEC F4) | 1010 CFU probiotic in 1 mL skimmed milk/day | Reduced ETEC levels in the ileum | Improved daily weight gain | [70] | |
| Saccharomyces cerevisiae | 21 days old/21-d trial | 5, 10, 20 g/kg probiotic | Increase in jejunal villus height and villus height: crypt depth ratio was also increased; gut IFN-gamma concentration increased on day 21 but plasma IFN-gamma reduced on day 7 and CD4 reduced on day 14 | Feed intake was enhanced with the inclusion of 5 or 10 g/kg; enhanced digestibility of DM, CP, GE with 5 g/kg yeast supplement | [71] | |
| Lactobacillus johnsonii and Lactobacillus mucosae single or combined | 21 days old/21-d trial | 5 × 107 or 108 CFU/g/piglet/day of each strain | Increase in fecal Lactobacillus and reduction in E. coli counts | Both single or combined supplementation increased overall FI and BWG | [72] | |
| Enterococcus faecalis | 24–26 days old/28-d trial | 0.5 × 109, 1 × 109, or 2.5 × 109 CFU/kg of feed | Lower incidence of diarrhea and increase in fecal Lactobacillus counts | Higher ADG and feed efficiency in pigs receiving the highest probiotic dose. | [73] | |
| Bacillus subtilis GCB-13-001 (1 × 109 CFU/kg) | 28 days old/42-d trial | 1 g/kg | Fecal Lactobacillus counts were improved, and E. coli counts were reduced | The BW and ADG improved during all phases, F:G improved during the overall experiment period | [74] | |
| B. coagulans, B. licheniformis, B. subtilis, and C. butyricum mixed dried spores(1 × 1012, 5 × 1011, 1 × 1012 and 1 × 1011 CFU/kg respectively). | 28 days old/42-d trial | 0, 1, 2, and 3 g/kg | Linear increase in fecal Lactobacillus counts and decreased Escherichia coli counts and ammonia (NH3) emission | Linearly increased ADG and ADFI during d 0–7, increased ADG and G:F during d 8–21 | [75] | |
| Bacillus subtilis or Bacillus pumilus 1 × 109 CFU/kg | 21 days old (challenged with ETEC/28-d trail) | 500 mg/kg | Bacillus subtilis supplement alleviated diarrhea severity, enhanced gut health, and reduced systemic inflammation of weaned pigs infected with ETEC F18 | Bacillus subtilis supplementation improved growth performance | [76] | |
| Phytogenics | ||||||
| Mixture of herb extract | Buckwheat, thyme, curcuma, black pepper, and ginger | 21 days old/42-d trial | 250 mg/kg | Reduced fecal E. coli counts | Improved energy digestibility but no effect on growth performance | [77] |
| Plant extract(PE) | Green tea leaves (Camellia sinensis) and pomegranate fruit (Punica granatum) | 24 days old/35-d trial | 8 μL/kg per day PE in drinking water | Reduced fecal E. coli counts in challenged pigs on days 14 and 35 and reduced Enterobacteriaceae on day 35 | Increased ADG from days 28–35 and increase G:F ratio from days 7 to 14 | [78] |
| Brown algae (ethanol extract from Ecklonia sp.) | Eckol | 28 days old/42-d trial | 0.5 and 1 g/kg | Improved intestinal barrier function | Improved growth performance, and reduced the levels of stress hormones (cortisol, epinephrine, and norepinephrine) and antioxidants (superoxide dismutase and glutathione peroxide) | [79] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Upadhaya, S.-D.; Kim, I.-H. The Impact of Weaning Stress on Gut Health and the Mechanistic Aspects of Several Feed Additives Contributing to Improved Gut Health Function in Weanling Piglets—A Review. Animals 2021, 11, 2418. https://doi.org/10.3390/ani11082418
Upadhaya S-D, Kim I-H. The Impact of Weaning Stress on Gut Health and the Mechanistic Aspects of Several Feed Additives Contributing to Improved Gut Health Function in Weanling Piglets—A Review. Animals. 2021; 11(8):2418. https://doi.org/10.3390/ani11082418
Chicago/Turabian StyleUpadhaya, Santi-Devi, and In-Ho Kim. 2021. "The Impact of Weaning Stress on Gut Health and the Mechanistic Aspects of Several Feed Additives Contributing to Improved Gut Health Function in Weanling Piglets—A Review" Animals 11, no. 8: 2418. https://doi.org/10.3390/ani11082418
APA StyleUpadhaya, S.-D., & Kim, I.-H. (2021). The Impact of Weaning Stress on Gut Health and the Mechanistic Aspects of Several Feed Additives Contributing to Improved Gut Health Function in Weanling Piglets—A Review. Animals, 11(8), 2418. https://doi.org/10.3390/ani11082418