1. Introduction
Research into the developmental origins of health and disease (DOHaD) has revealed the far-reaching effects that both maternal under- and overnutrition can have on the epigenome of the developing embryo and fetus, with the potential to influence susceptibility to lifestyle-related diseases (e.g., hypertension and type 2 diabetes) into adulthood [
1]. Overfeeding is an increasingly common issue in horses and is currently a topic of interest primarily because of its association with an increased risk of metabolic syndrome [
2] in the individual overfed animal. Equine metabolic syndrome is the collective term for a group of associated clinicopathological changes that includes obesity, insulin dysregulation (ID) and other related endocrine disturbances, and which together predispose to the development of endocrinopathic laminitis [
2]. A less well-investigated area of study is the possibility for transgenerational effects and, in particular, whether maternal overfeeding or obesity in the periconception period affects the epigenome of the horse embryo and may predispose the offspring to the development of metabolic or orthopedic disease later in life [
3].
Epigenetics refers to changes in gene expression that occur without any change in the DNA sequence itself, for example inhibition of gene transcription by methylation of the DNA or acetylation of the binding histones [
4]. During gametogenesis and preimplantation development, epigenetic reprogramming must occur to ensure that the zygote/early embryo is capable of giving rise to all the cell types required to produce a viable embryo, fetus and neonate. During the initial period of reprogramming, nearly all of the epigenetic marks accumulated by the parental genomes are erased, e.g., by global DNA demethylation, with the exception of the parentally imprinted marks, such that the early blastomeres become totipotent [
5]. The process of re-acquiring epigenetic marks, also known as DNA methylation reprogramming (DMR), then starts at or directly before blastocyst formation (day 5–7 after ovulation in the horse) [
6], when cells begin to choose lineage fates, differentiate and lose potency [
5,
7]. The way in which embryonic demethylation and subsequent re-methylation occurs is influenced by the environment in which it occurs. In particular, in other species, maternal nutrition has been shown to influence epigenetic reprogramming, with the potential for lifelong effects on gene expression in the offspring, including increased susceptibility to complex, lifestyle related diseases in adulthood [
8].
DMR also plays an important role in regulating the expression of imprinted genes [
4]. Genomic imprinting is an epigenetic modification that involves inactivation of one of the two alleles of a gene in a parent-of-origin specific manner [
9]. In general, imprinted genes are important for fetoplacental development and help regulate the nutrient supply to the fetus by influencing the size, architecture and nutrient transfer capacity of the placenta [
10]. In this respect, it has been proposed that imprinted genes play a critical role in maternal-fetal resource allocation to balance maternal needs against fetal demands [
9]. Based on this theory of a conflict for resources, maternally expressed genes, such as
IGF2R and
GRB10 [
11,
12,
13], are expected to limit conceptus growth, whereas paternally expressed genes, such as
PEG10 [
14], should enhance fetoplacental growth [
9].
IGF2 and
IGF2R have further been proposed to be key regulators of early placental formation, which in turn determines the capacity for fetal and placental growth during the remainder of gestation [
13]. Murine studies have revealed profound consequences of maternal overnutrition and obesity on the expression of imprinted genes [
15,
16], suggesting, for example, that pre-conception exposure to a high-fat (HF) diet may influence re-programming of imprinted marks during early embryonic development. Besides the potential effects on embryonic epigenetic reprogramming, maternal diet clearly has the capacity to influence embryonic and fetal nutrient provision throughout gestation. In the horse, definitive placental formation, characterized by interdigitation of the chorioallantois with the endometrium, only starts at around day 40 of gestation, indicating that the capacity for uptake of nutrients during the first part of gestation is limited [
17]. The equine conceptus is entirely dependent on histotrophic nutrition during the first 40 days of development [
18]. Histotroph is synthesized by the endometrial glands and luminal epithelial cells and secreted into the uterine cavity; this “uterine milk” contains nutrients such as amino acids, carbohydrates, and lipids, together with growth factors, cytokines, and hormones [
19]. The composition of the histotroph is determined by factors including the prevailing maternal endocrine milieu (e.g., circulating progesterone and estrogen concentrations) [
20], endometrial nutrient transporters and, presumably, maternal circulating nutrient concentrations. The conceptus membranes also express specific transporters to facilitate the uptake of nutrients present in the histotroph into the embryo [
18,
21], and the expression of certain nutrient transporters may, in turn, be adjusted in response to the abundance of nutrients in the uterine cavity. For example, rodent and mice studies show increased expression of various amino acid and glucose transporters in the trophectoderm or placenta (respectively, day 18.5 and day 21) of mice fed an HF [
22] and rats fed a HSHF [
23] diet, when compared to control chow-fed mice or rats. Placental capacity to transfer glucose and amino acids to the fetus has also been shown to be enhanced in response to a maternal diet high in simple sugars and saturated fats [
16].
The aim of this study was to determine the effect of maternal overfeeding on early epigenetic reprogramming and pathways for conceptus nutrient uptake. Mares were fed either a high-energy (HE) diet equating to 200% of net energy requirements, which resulted in gradual onset of obesity and ID [
24], or a control maintenance diet. The effect of maternal overfeeding on pre-implantation conceptus development was investigated by transferring day-seven embryos (i.e., immediately after blastocyst formation) between HE and control mares and assessing the expression of genes coding for various imprinted genes and DNMTs, glucose and amino acid transporters in the conceptus membranes at day 28 of gestation (i.e., late in the pre-implantation period). To further assess the effects of maternal overfeeding on embryonic nutrient handling, the lipid droplet content was compared between day-seven embryos originating from control versus HE donor mares, and glucose uptake was examined in the extra-embryonic membranes of day-28 conceptuses, resulting from transfer of embryos between mares from the same or the alternate feeding group.
4. Discussion
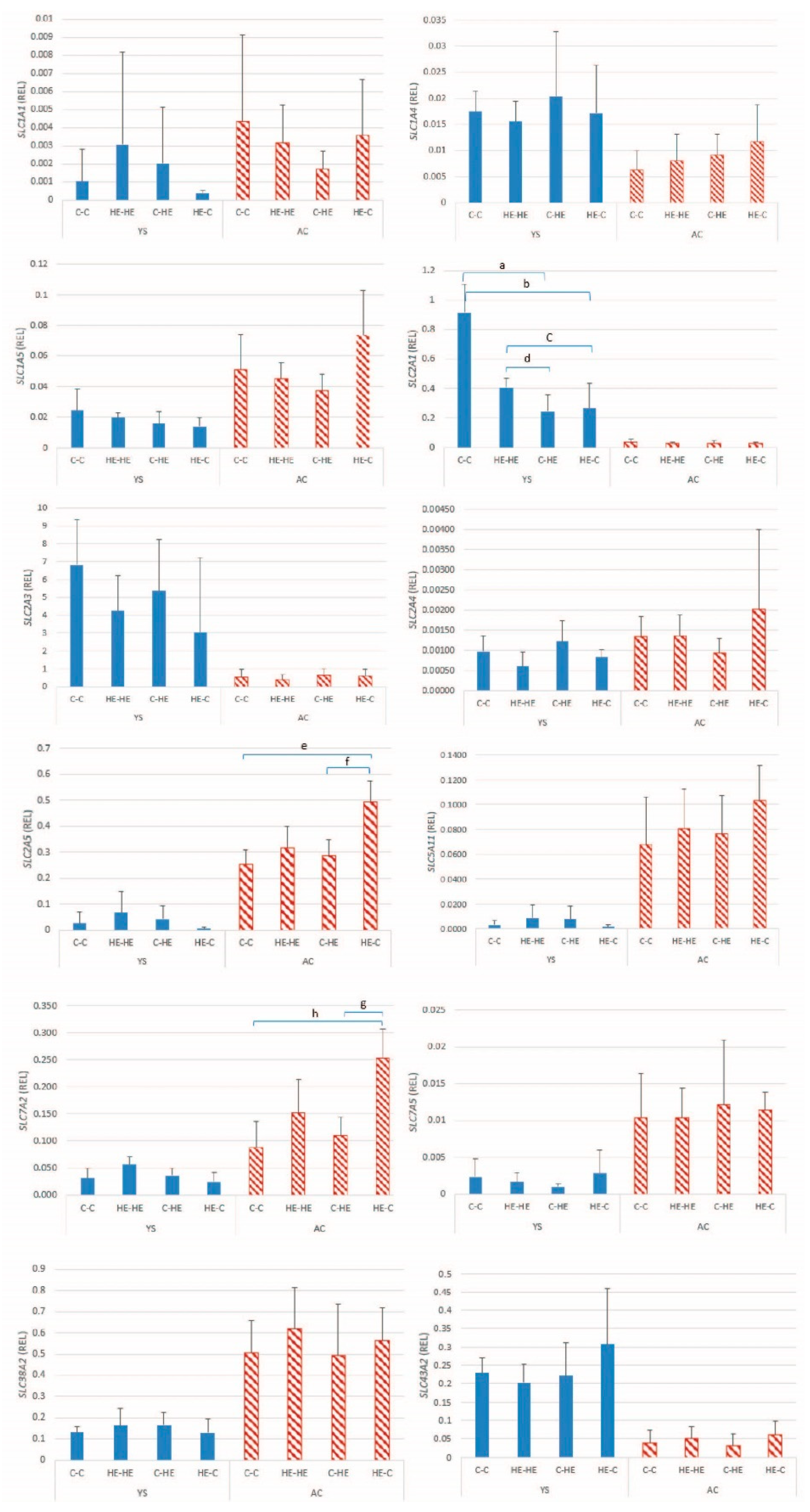
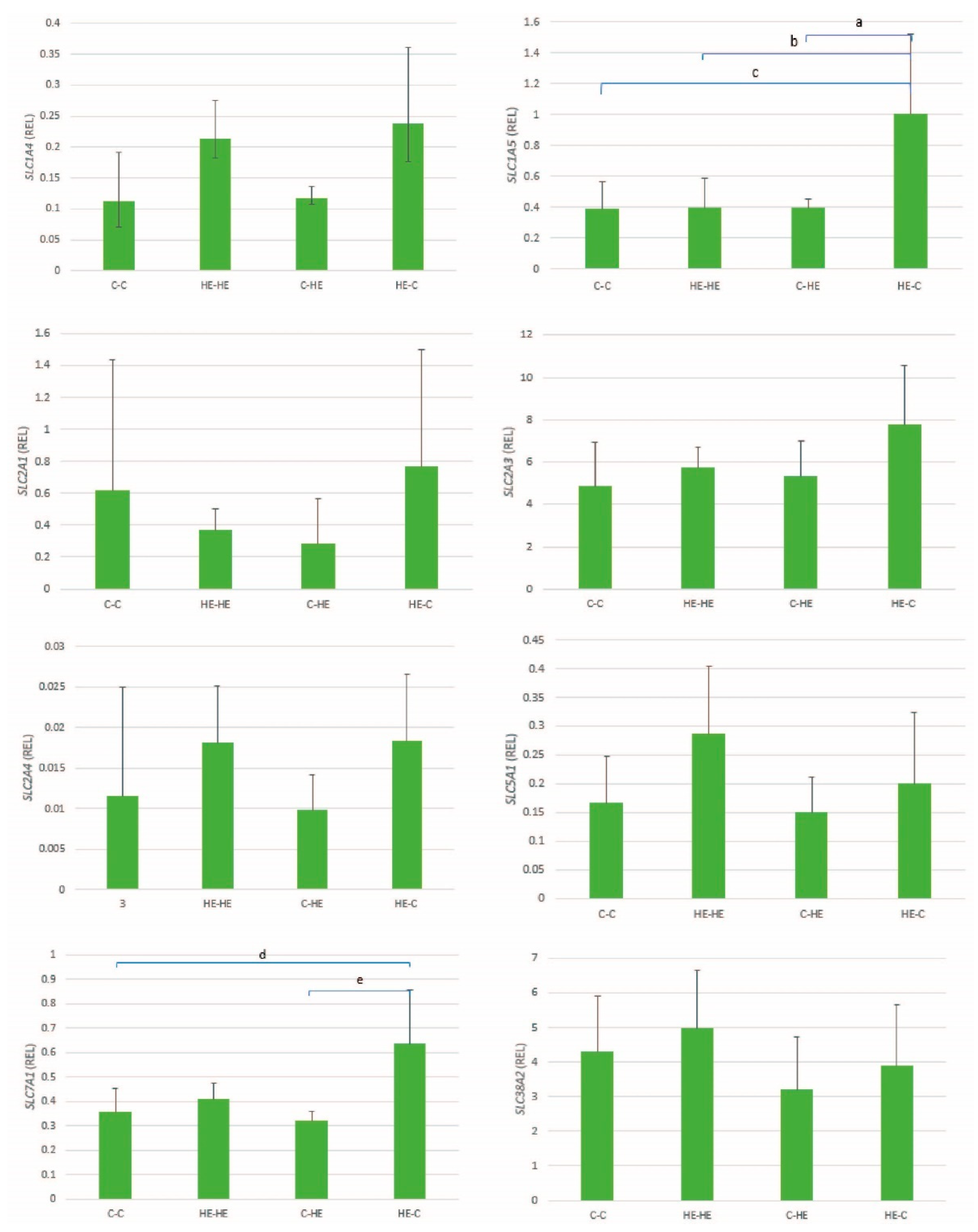
This study aimed to examine the effect of maternal overfeeding during the periconception period on nutrient handling and imprinted gene expression in Shetland pony preimplantation conceptuses. To this end, we examined the expression of a range of glucose and amino acid transporters and genes involved in DNA methylation and imprinted genes, in which the latter are thought to be particularly sensitive to epigenetic perturbation during the periconception period. In addition, we examined the actual glucose uptake into the extra-embryonic membranes and lipid accumulation in blastocyst stage embryos and the expression of glucose and amino acid transporters in the endometrium of the recipient mare. Maternal overfeeding was found to affect the expression of three maternally and four paternally expressed genes, two DNMTs, two glucose transporters and one amino acid transporter in the extra-embryonic membranes of day-28 conceptuses and the expression of two different amino acid transporters in the endometrium of recipient mares; in almost all cases, this effect took the form of significantly higher expression when embryos were transferred from an HE donor to a control recipient mare. Conversely, there was no significant effect of maternal overnutrition on embryonic content of lipid droplets on day 7 of gestation or on glucose uptake by the extra-embryonic membranes on day 28 of gestation.
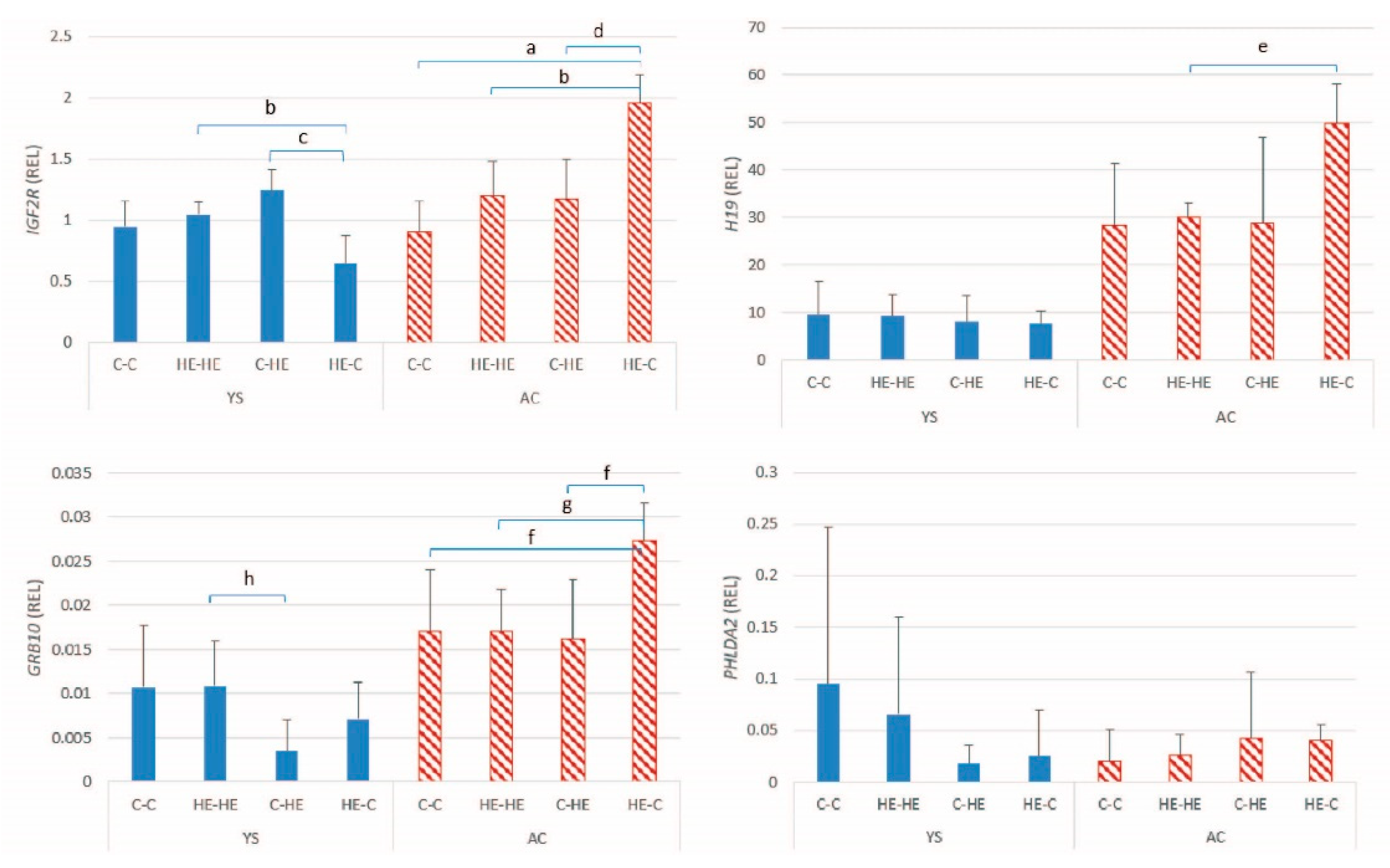
An association was found between maternal overfeeding and the expression of seven imprinted genes in conceptus membranes at day 28 of gestation; these included three maternally expressed (
IGF2R, H19 and
GRB10) and four paternally expressed genes (
PEG10, NDN, IGF2 and
SNRPN). The overall pattern was in agreement with previous studies that reported an effect of maternal overfeeding on placental gene expression in mice [
15,
16]. For example, exposure to a high-sugar, high-fat (HSHF) diet during murine gestation resulted in altered placental expression of a range of imprinted genes and associated changes in metabolic signaling pathways involved in placental growth, metabolism and nutrient transport at day 16 of gestation; this included upregulation of
GRB10,
IGF2, and the maternally expressed
H19 gene that is linked to IGF2 expression [
16]. Similarly, for murine embryos transferred at the two-cell stage between mice fed either a control or high-fat (HF) diet, both a pre-gestational and gestational HF diet resulted in fetal growth restriction and abnormal placentation [
15] and, in the group in which embryos were transferred from HF to control dams, several imprinted genes, including
IGF2 and
IGF2R, which were differentially expressed in the placenta compared to when embryos were transferred between control dams [
15]. For most of the imprinted genes in the present study, the relative expression was higher in the conceptus membranes of embryos transferred from an HE donor to a control recipient mare, irrespective of whether the genes were maternally or paternally expressed; the exception was
NDN, which showed a drop in expression. That the expression of these genes was higher in the conceptus membranes of embryos originating from an HE donor and transferred to a control maternal environment on day 7 of gestation suggests that maternal overnutrition or obesity has an effect on the oocyte or on the early embryo, as previously proposed in both mice [
15] and horses [
34,
35]. However, there was no increase in expression of imprinted genes in the HE-HE group, although these embryos also originated from an HE donor mare; this is in agreement with the modest differences reported by Sessions-Bresnahan et al. [
35] in expression of a range of genes between day 16 conceptuses from obese versus normal mares. Our findings may therefore indicate that the transfer from an excessively nutrient-rich to a “normal” maternal environment results in a mismatch between embryonic expectations and maternal provision and that it is this change that leads to dysregulation of a range of imprinted genes. In this case, early exposure to an HE diet will still need to be responsible for underlying changes that only become apparent when the uterine environment does not meet the altered embryonic requirements. As reported in a previous study, there were no significant between-embryo transfer group differences in crown-rump length or weight of the embryo at day 28 of gestation [
36], which presumably indicates that the relative upregulation of the imprinted genes resulted in a better match between the maternal environment and embryonic needs, rather than leading to excessive or restricted embryonic growth. The exact nature of the relationship between maternal overnutrition and the changes in imprinted gene expression can, however, not be determined based purely on our results. Moreover, whether these changes persist during later gestation and into post-natal life has not been established.
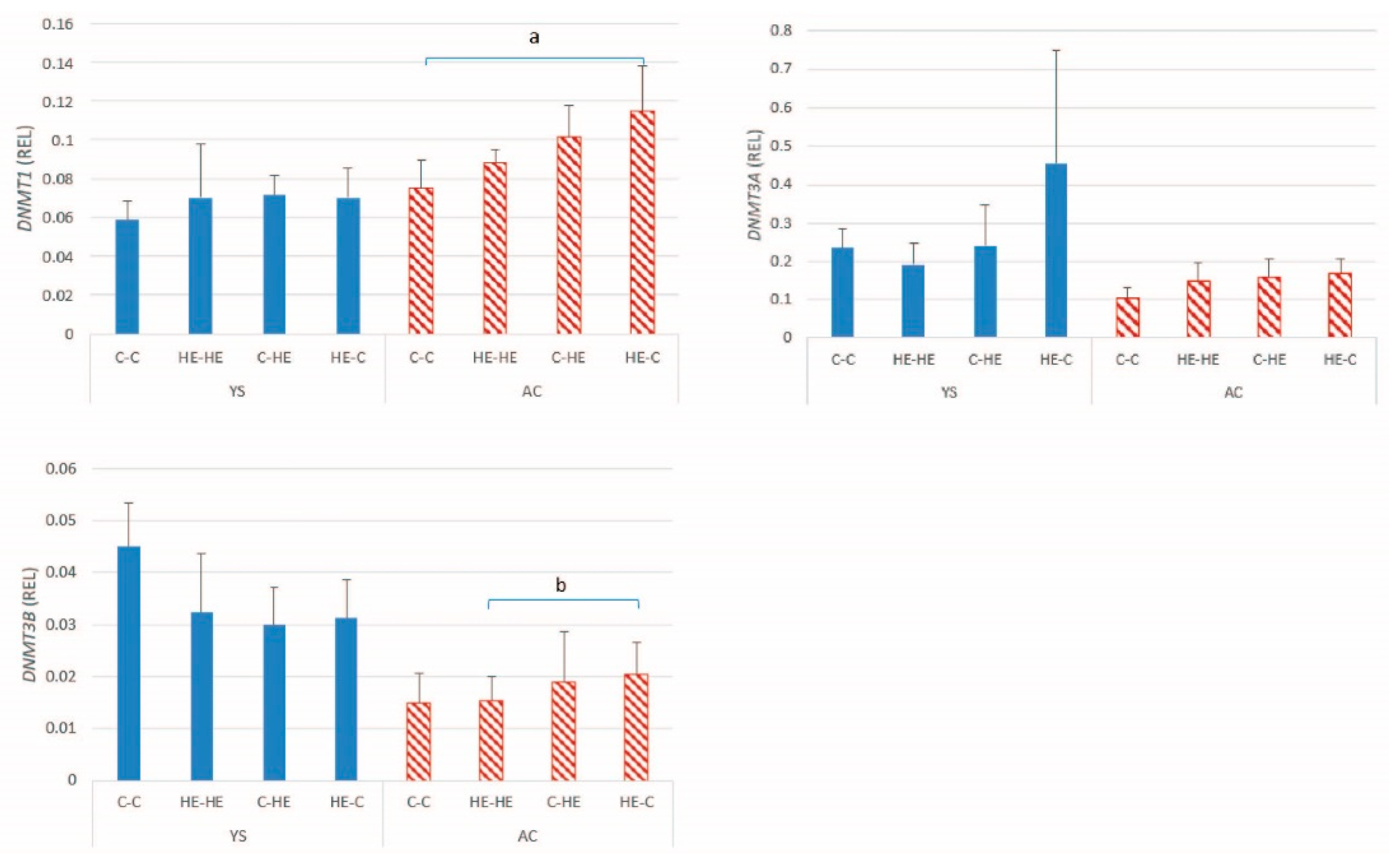
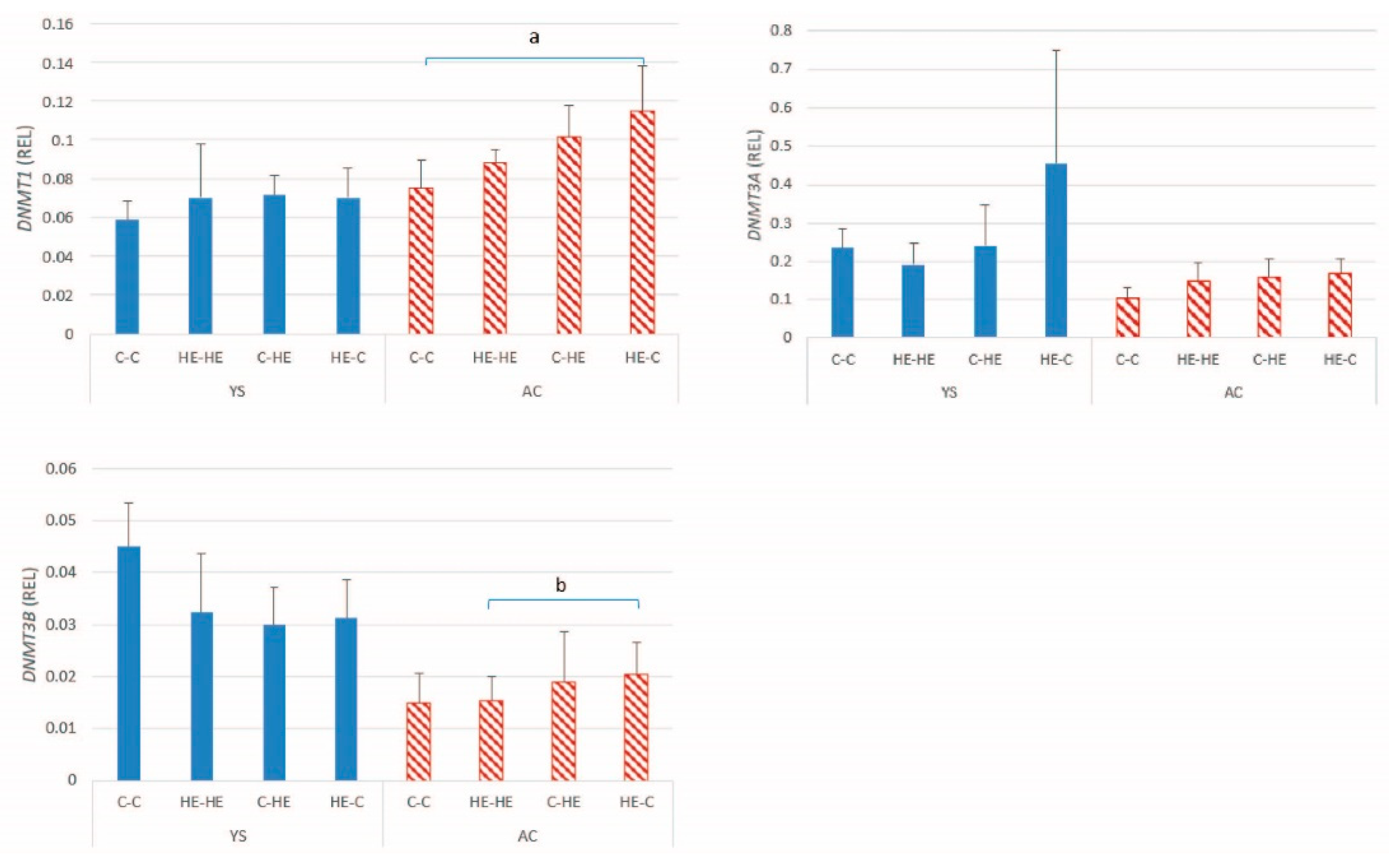
Since changes in DNA methylation are likely to play an important role in regulation of gene expression and maintenance or re-establishment of imprinting status, we also examined expression of the DNA methyltransferases. A significant effect of maternal overnutrition was evident for
DNMT1 and
DNMT3B expression in the AC of day-28 conceptuses. Previous studies in other species have described effects of maternal nutrition on the DNA methylation status of developing conceptuses. For example, in the placenta of obese human mothers, global methylation levels are higher than in the placenta of normal weight mothers at birth [
37]. In the present study, a similar pattern for the change in expression of
DNMTs 1 and
3B was found to be that described for the imprinted genes, namely higher expression in the AC of conceptuses resulting from transfer of an embryo from an HE donor to a control recipient mare. This finding may be reflected by an alteration in the DNA methylation status of embryos transferred from an overfed mare to a neutral maternal environment; however, DNA methylation status was not examined. Conversely, Sessions-Bresnahan et al. [
35] compared global DNA methylation status of day-16 equine conceptuses recovered from normal versus obese mares and reported a tendency to higher levels of hydroxy-methylation in embryos from the normal mares, suggesting that the changes we observed in DNMT expression most likely result from the effects of maternal overnutrition (or associated increase in body condition) on the oocyte or early embryo. However, similar to the discussion for imprinted genes, no significant increase in DNMT expression was evident in the HE-HE group, suggesting that any effects at the oocyte level or before day 7 of embryo development only become apparent when the embryo is exposed to an environment that does not meet its expectations or after a longer period of exposure to maternal overnutrition or obesity.
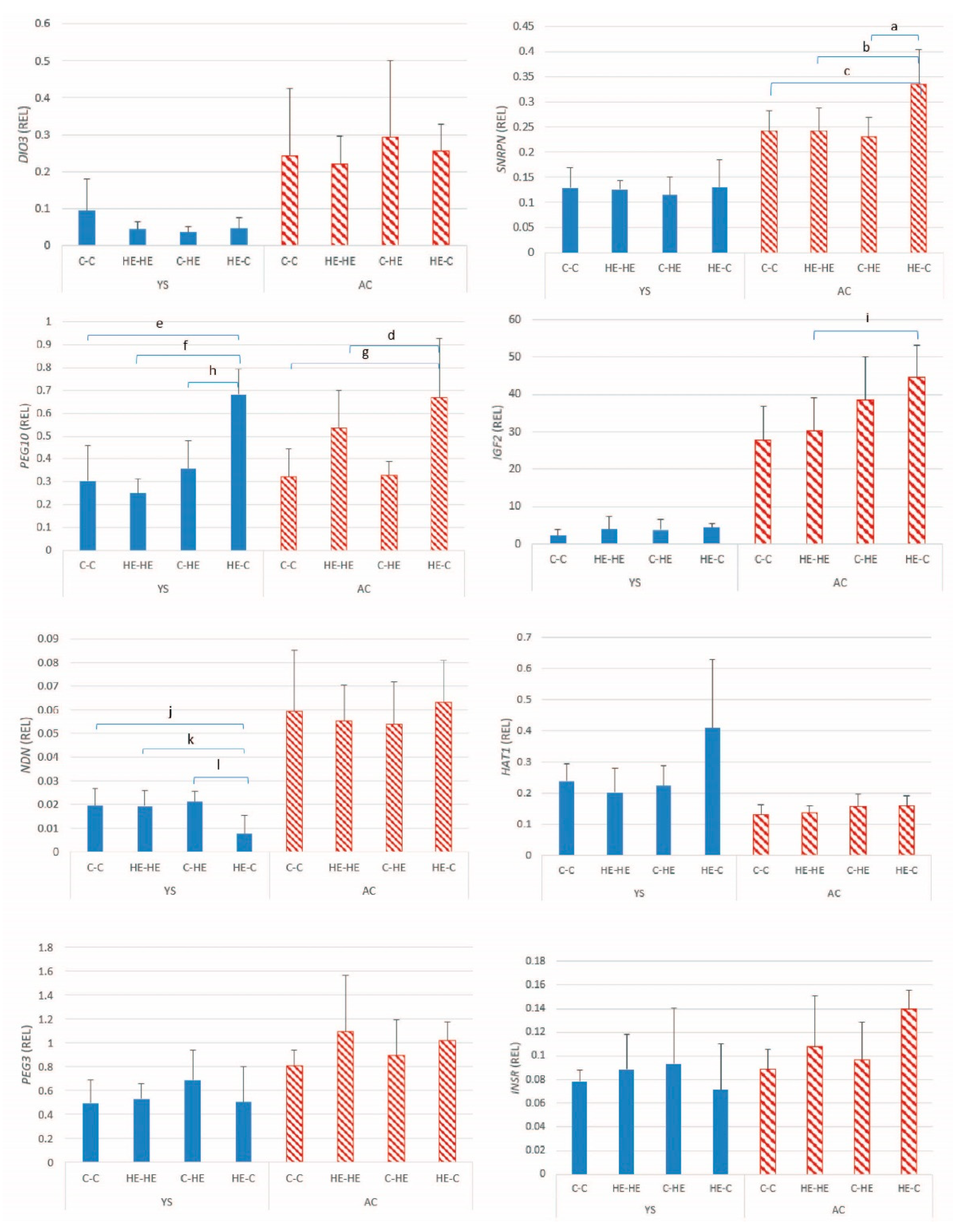
Following its arrival in the uterus at the blastocyst stage of development, the primary energy substrate for the developing embryo is glucose. Moreover, the trophectoderm and extra-embryonic membranes of the pre-implantation equine conceptus express a number of glucose transporters, which help ensure adequate glucose provision to the embryo [
18]. In the present study, a significant effect of maternal overfeeding was found on the expression of
SLC2A1 (GLUT-1) in the YS membrane, and on
SLC2A5 (GLUT-5) in AC at day 28 of gestation. As for certain imprinted genes and DNMTs, relative AC
SLC2A5 expression was higher after transfer of embryos from an HE donor to a control recipient mare. In contrast,
SLC2A1 expression was higher in the YS membrane of conceptuses, resulting from transfer of an embryo from a control donor to a control recipient mare, which presumably reflects a down-regulation in the other groups as a response to overfeeding of either the donor or the recipient mare. In the horse,
SLC2A1 has been proposed to play a role in transporting glucose into the yolk sac via facilitated glucose diffusion [
18], which explains the relatively high expression detected in the YS membranes in the present study. In the case of maternal overnutrition, downregulation of
SLC2A1 in the YS membrane presumably helps prevent the embryo from being oversupplied with glucose. A previous study showed that the expression of GLUT-1 protein significantly decreased in day-9–10 rat embryos and yolk-sac tissue cultured in medium containing high concentrations of glucose (33.3 and 66.6 mmol/l glucose) for 48 h [
38]. The expression of GLUT-1 mRNA was, however, not downregulated in that study. GLUT-1 is a fructose transporter [
39], and upregulation of expression in the AC of the HE-C group may reflect an imbalance between the fructose needs of an embryo that initially developed in an HE donor mare and presumed lower supply of fructose in the uterine environment of a control recipient mare.
For the amino acid transporters, only
SLC7A2 in the AC of embryos transferred from an HE donor to a control recipient mare showed an altered pattern of expression, namely upregulation.
SLC7A2 has previously been shown to be upregulated in equine conceptus membranes as early pregnancy proceeds and has been suggested to help delivery of amino acids from the uterine lumen to the trophectoderm [
21]. The relatively high expression of
SLC7A2 may reflect an imbalance between amino acid demands of an embryo that initially developed in an HE environment and supply in the control uterine environment.
The general findings for nutrient transporter expression are in agreement with other studies, namely an increase in expression of certain amino acid and glucose transporters in the trophoblast membranes and early placentae of mice fed an HF [
22] or HSHF [
23] diet. However, Sasson et al. [
15] reported that the expression of the upregulated transporters normalized during later pregnancy and concluded that early changes in nutrient transporter expression are neither permanent nor responsible for differences in the metabolic phenotype. Moreover, they noted that there was no proof that upregulation of transporter expression necessarily resulted in a change in actual nutrient transport [
15]. Similarly, Sferruzzi-Perri et al. [
16] showed that while a maternal HSHF diet resulted in growth restriction of mouse fetuses on day 16 of gestation, this growth restriction was no longer evident by day 19; moreover, the expression of the nutrient transporters was unaffected, again, suggesting that temporal differences in the embryonic and fetal expression of genes encoding for nutrient transporters induced by maternal periconception overnutrition do not become permanent alterations. In the present study, we investigated glucose uptake by day-28 conceptus membranes and found no significant effect of maternal nutrition status on the uptake of the fluorescent glucose analog 2-NBDG, which is thought to be transported by both sodium-glucose linked transporters (SGLTs) and glucose transporters (GLUTs) [
33]. However, fructose uptake was not measured, and it cannot be determined whether maternal overfeeding affected fructose uptake via GLUT-5. The glucose analogue used in our study could have been taken up into the day-28 membranes via GLUT-5, but based on findings in other species, it cannot be confirmed or ruled out, and we suggest that GLUT-5 did not play a significant role in glucose analogue absorption [
39]. If the increase in gene expression for the fructose and amino acid transporters did not lead to increased uptake of glucose, fructose and amino acids, it may also help explain why maternal nutritional status did not affect embryo size or weight at day 28 of gestation [
36]. If few amino acid transporters are affected (only one of the seven we examined), the change is unlikely to have significant effects on the whole system since there is a large degree of overlap and redundancy among the amino acid transporters, and up- or down-regulation of the majority within a type (neutral, anionic, cationic) will be required to significantly impact overall amino acid transport [
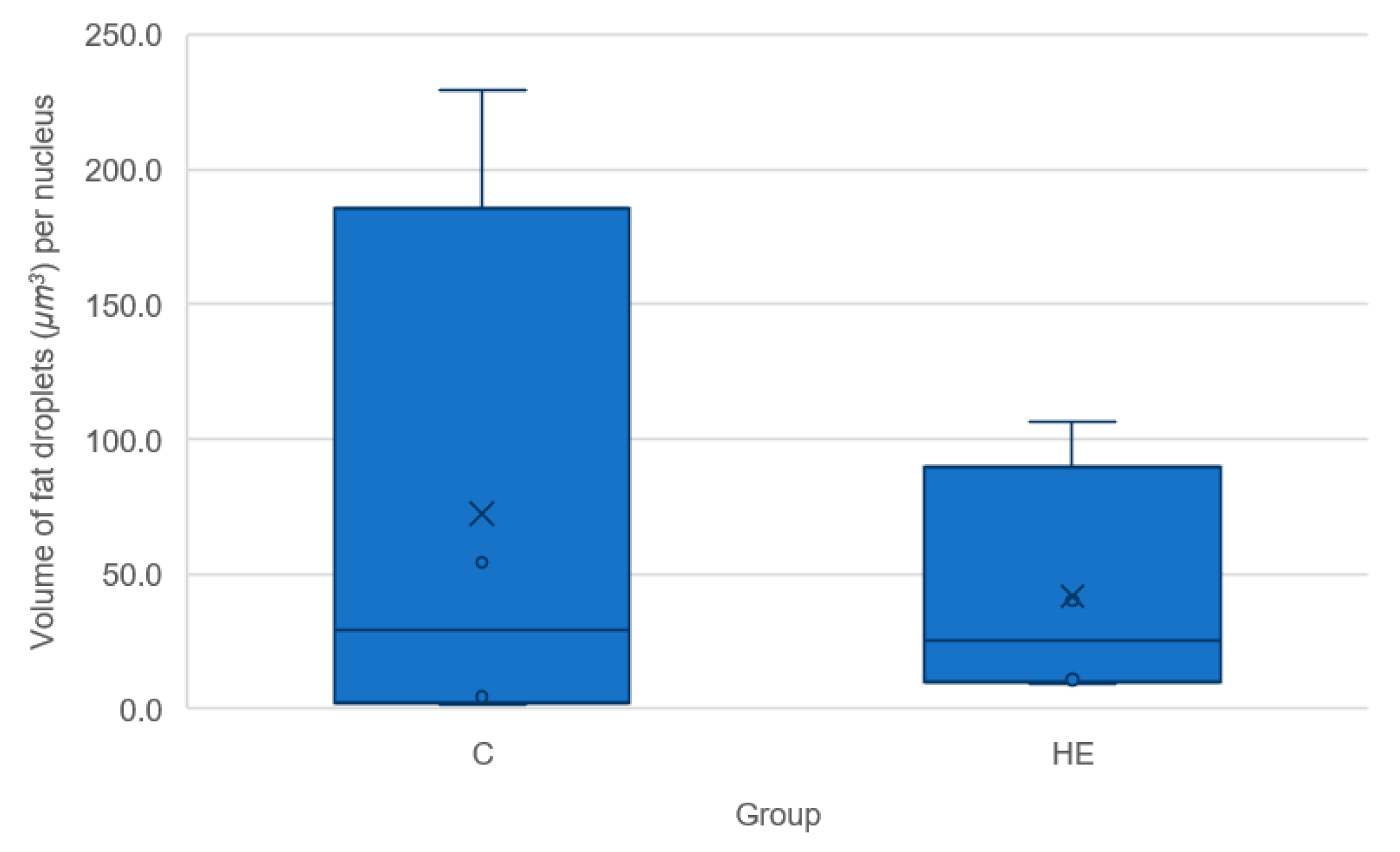
40]. In a similar vein, the lipid droplet content did not differ between blastocysts originating from control or HE donor mares, indicating that maternal overfeeding had no gross effect on lipid uptake and storage prior to day 7 of gestation.
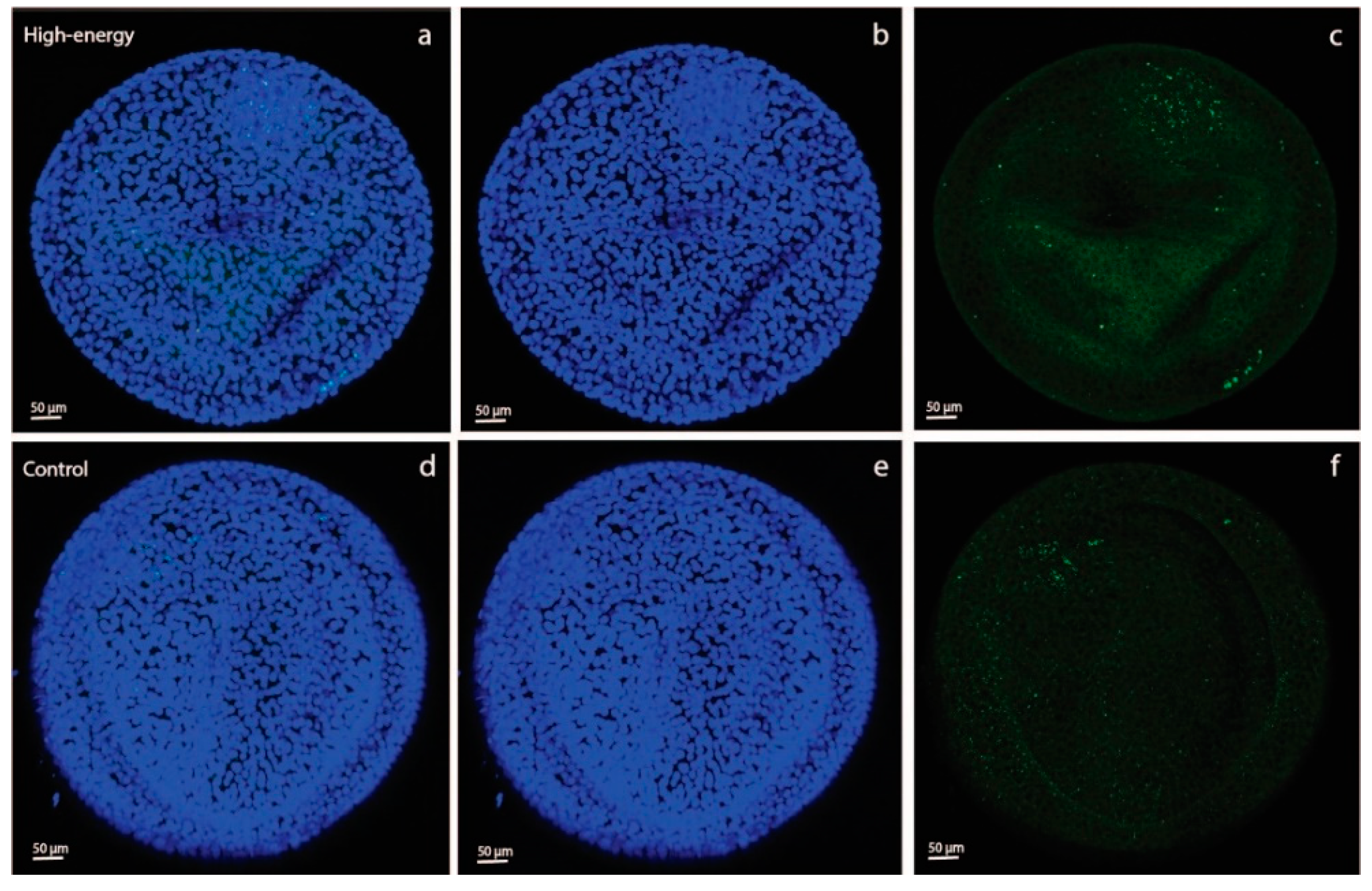
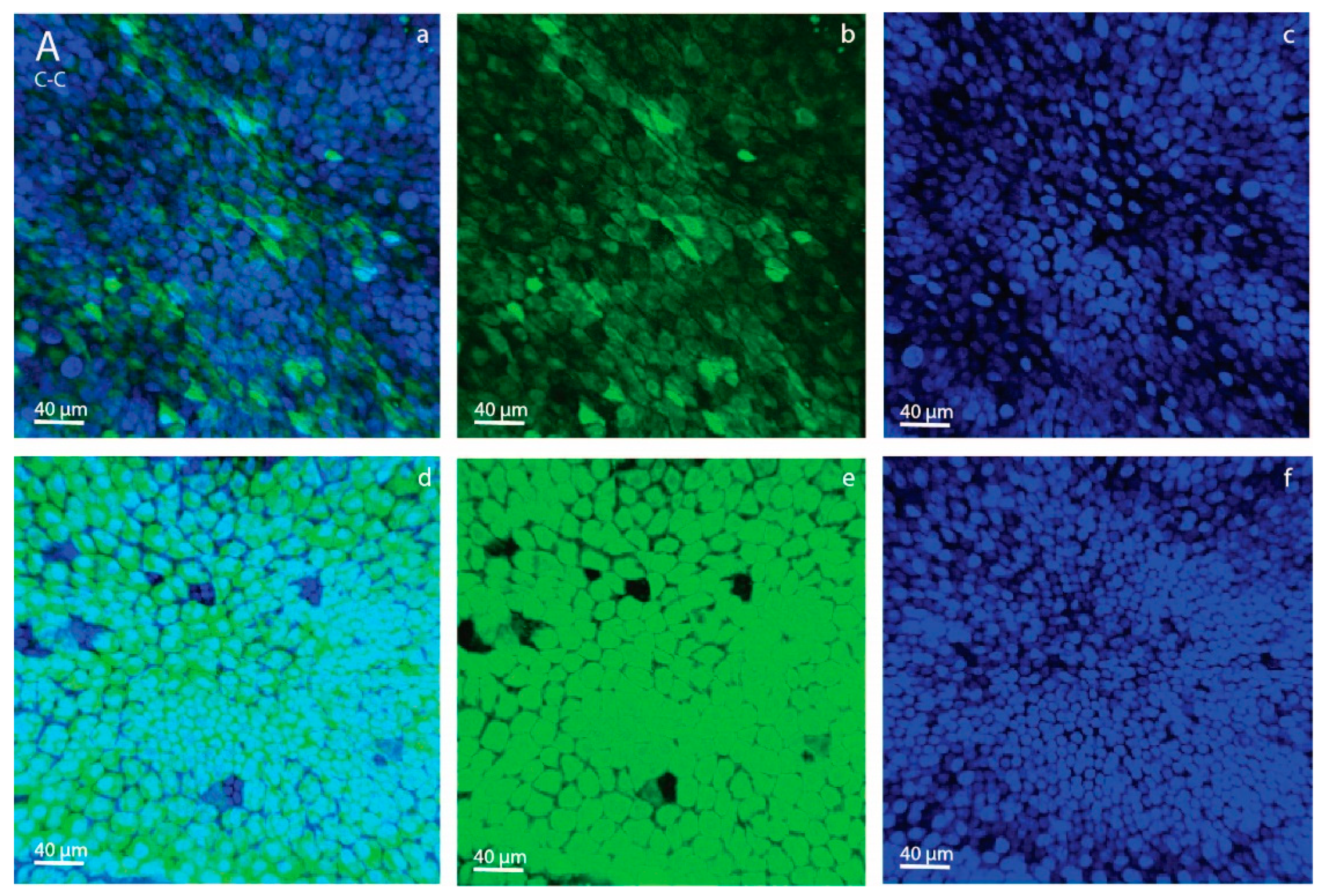
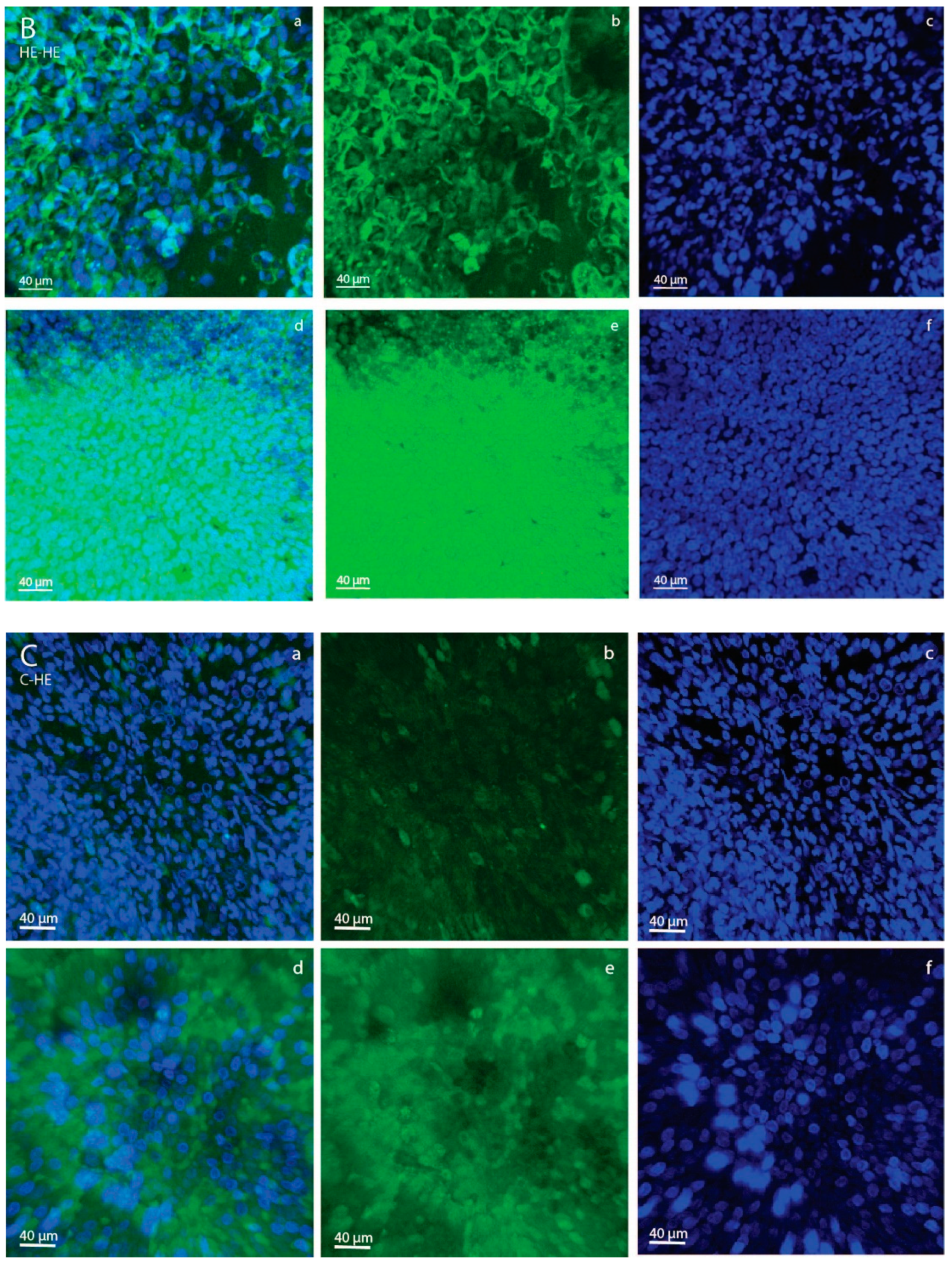
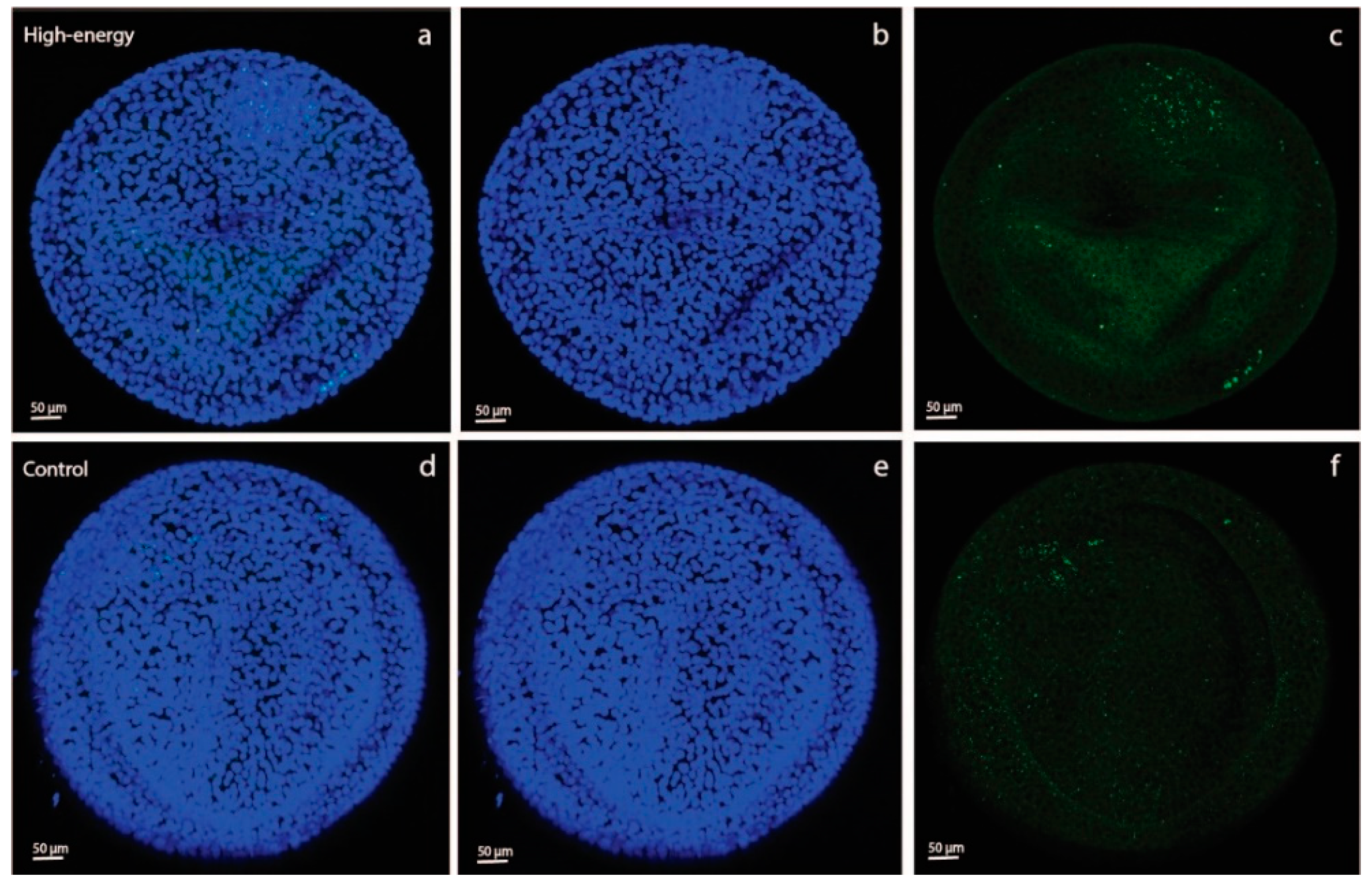
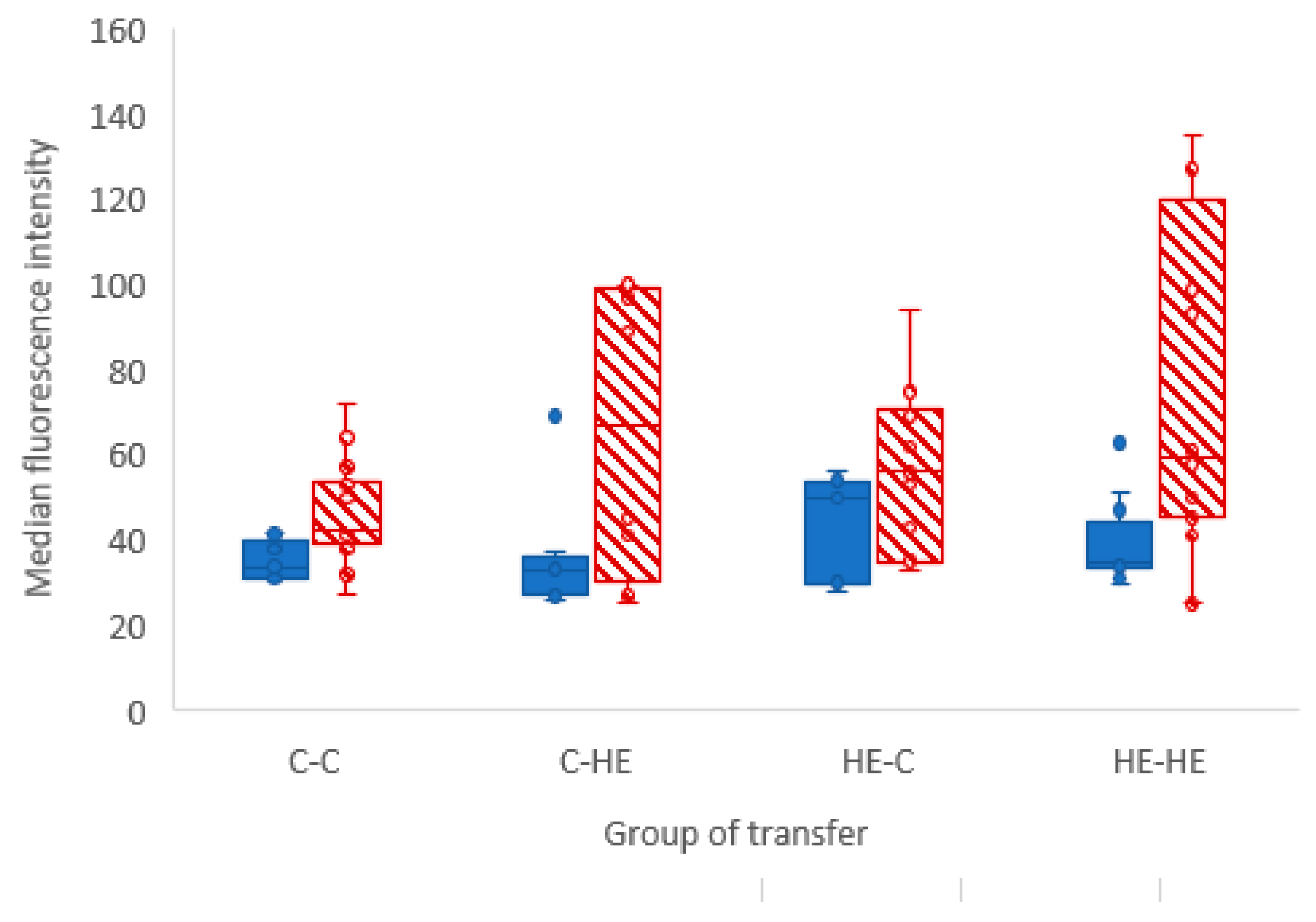
As an aside, we saw a difference in 2-NBDG fluorescence intensity between YS and AC in both the C-C and HE-HE groups, with a higher intensity in YS than AC (
Figure 2). This suggests that, in ponies, the day-28 YS membrane had a greater capacity to take up glucose than the AC. This presumably suggests that the YS is still the primary route of nutrient uptake, whereas the AC is not yet fully developed, e.g., has yet to sufficiently attach to and interact with the endometrium to take up its role as the major route for nutrient supply.
Another important potential contributor to the abundance of nutrients in the uterine lumen is the expression of nutrient transporters in the endometrium, and the expression of amino acid transporters in the mare’s endometrium has been shown to change during early pregnancy, presumably as a result of the presence of a conceptus and factors secreted by that conceptus [
21]. We found an upregulation of endometrial expression of the amino acid transporters
SLC1A5 and
SLC7A1 in control mares that received an embryo from an HE donor mare. Previously, the expression of
SLC1A5 was reported to be upregulated in equine endometrium between days 14 and 21 of gestation [
21], and the transporter is thought to play a crucial role in delivering amino acids to the uterine lumen [
21].
SLC7A1 has previously been reported to be downregulated during early pregnancy [
21]. Upregulation of both these amino acid transporters in the endometrium of control mares that received an embryo from an HE donor may reflect an embryo-directed effect to help meet demands for amino acids.
Due to the small number of donor and recipient mares included in this study, several were used twice within a single embryo transfer group and several mares were used during more than one study year, whereas others were used for one year only. Although statistical methods were selected to consider individual mare effects, it is possible that reuse of ponies within groups and over multiple years could have biased the results. Finally, embryos were not divided by sex (male versus female), whereas it is possible that embryos of different genders respond differently to maternal overfeeding; in this respect, Sessions-Bresnahan et al. [
35] reported a tendency for a higher level of global methylation in female but not male embryos recovered from normal weight, compared to obese mares.
In conclusion, transferring embryos from overfed Shetland pony mares to recipient mares on a maintenance diet resulted in increased expression of several imprinted genes (both maternally and paternally expressed genes), DNMTs 1 and 3B, and a number of glucose and amino acid transporters in the conceptus membranes on day 28 of gestation. Transfer of an embryo from an overfed donor to a control recipient mare also led to upregulated expression of two different amino acid transporters in the endometrium of the recipient mare. Although transferring horse embryos from overfed to control mares seems to disturb gene expression, it did not affect glucose uptake by the conceptus membranes on day 28 of gestation, and it remains to be seen whether the alterations in gene expression are maintained throughout gestation and into postnatal life.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}