1. Introduction
Selenium (Se) is an essential trace element that is widely thought to support oxidative stress tolerance in fish. Although the tolerance range of Se in biological systems is very narrow [
1], the Se requirements of different fish may vary greatly not only in a species-dependent manner but also depending on the molecular form of Se (e.g., organic, inorganic, and nano-forms) [
2,
3,
4]. Other factors that influence Se requirements include fish size, health status, dietary formulation, experimental conditions, and water quality [
5]. Previous studies have indicated that the presence of appropriate Se contents in feed can promote fish growth, whereas excessive Se concentrations may inhibit growth [
6,
7,
8]. Specifically, excessively high dietary Se concentrations lead to oxidative stress and an increased hemolysis rate, resulting in reproductive failure, tissue damage, and organ malformation [
9,
10,
11]. Lee et al. [
12] utilized Se methionine as a Se source and described that the growth performance, survival rate, and nonspecific immunity of Nile tilapia (
Oreochromis niloticus) were significantly reduced when the Se intake was more than 2.06 mg Se/kg diet.
Bamboo charcoal (i.e., an activated carbon material) is made from the thick stems of bamboo through dry distillation. Importantly, this material can adsorb a variety of molecules and contains a complex network of pores of different shapes and sizes [
13]. Due to the unique structure of the micropores in the bamboo stem, bamboo charcoal is thought to possess a higher adsorption capacity. For instance, this material has been used to adsorb ammonia nitrogen in water and decrease heavy metal intake [
14]. Given its activity as an insoluble carrier, bamboo charcoal can be used as a dietary supplement in fish feed to nonspecifically absorb potentially toxic agents. Activated carbon can be used as an indigestible carrier to prevent the intake of toxic substances through the digestive tract, and thus bamboo charcoal powder is often used as an oral antidote to treat poisoning [
15]. Therefore, bamboo charcoal and activated carbon have been widely used as adsorbents or detoxification agents in both animals and humans [
16,
17]. Previous studies have reported that dietary supplementation with bamboo charcoal has positive effects on animal growth, feed efficiency, specific growth rate, feed intake, nitrogen excretion, and digestion [
15,
18,
19,
20,
21].
Previous studies on the effects of dietary charcoal supplementation in animals have largely focused on mammals and some fish species, with a particular focus on animal growth [
22,
23,
24,
25]. However, to the best of our knowledge, the effects of dietary charcoal supplementation on trace element uptake and antioxidant performance in fish have not been previously characterized.
In previous studies, we determined that juvenile blunt snout bream (
Megalobrama amblycephala) require a Se dietary intake of 0.958–0.959 mg/kg (our assays were conducted using sodium selenite as a Se source) based on the quadratic fitting method. Nevertheless, the growth performance was significantly affected and the expression of anti-inflammatory factors was inhibited when the dietary Se intake was too high [
7]. Given its activity as an insoluble carrier, bamboo charcoal can be used as a dietary supplement in fish feed to absorb and slowly release excess nutrients. However, these hypotheses have not been evaluated in diets containing excessively high Se levels and are thus worth exploring. Therefore, our study sought to investigate the ability of bamboo charcoal to reduce the negative effects of high dietary selenium (Se) concentrations in
Megalobrama amblycephala.
4. Discussion
Many studies have reported that dietary charcoal supplementation promotes fish growth. Thu et al. [
23] fed
Paralichthys olivaceus with diets containing different amounts of bamboo charcoal (0%, 0.25%, 0.5%, 1%, 2%, and 4%) for 50 day. Interestingly, the 0.5% group exhibited the highest WG and SGR values. Similarly, Michael et al. [
22] fed red tilapia with diets containing different amounts of charcoal (0, 10, 20, 30, and 40 g/kg) for 60 day. The authors reported that the 30 g/kg and 40 g/kg groups exhibited significantly higher WG and SGR values, whereas the FCR of the 30 g/kg and 40 g/kg groups decreased significantly. This may have been because bamboo charcoal can absorb toxic substances in the intestine, thus improving intestinal function (i.e., nutrient absorption) and promoting growth [
15]. Previous studies have demonstrated not only that high supplementation levels of Se in feeds affect its utilization but also that excessive dietary Se is linked to fish growth inhibition (e.g., reduced WGR and SGR, increased mortality) [
7,
27]. In this study, both the 2 and 3 mg/kg bamboo charcoal concentrations in the Se-rich feed significantly improved the WGR and SGR values of juvenile blunt snout bream, suggesting that bamboo charcoal supplementation improves dietary Se utilization when administered in appropriate amounts. This was likely because bamboo charcoal absorbs excess nutrients in feed and improves the absorption and metabolic capacity of the digestive tract by slowing down the release of nutrients. However, dietary bamboo charcoal supplementation significantly reduces the average daily feed intake [
28]. Additionally, carbon supplementation has also been reported to inhibit growth when exposure concentrations are too high [
29]. These observations are likely attributed to two factors. First, one of the components of bamboo charcoal powder may have adverse effects on the digestive tract. Second, bamboo charcoal powder may induce bowel movements, thus shortening food retention and nutrient digestion and absorption. In turn, this reduces protein efficiency and increases feeding rates. Likewise, in our study, increasing bamboo charcoal concentrations led to increases in FCR values coupled with decreases in the PER.
Bamboo charcoal has a high absorption capacity and is thus considered an ideal insoluble carrier due to the unique structure of the micropores in the bamboo stem. For example, a study reported that the tolerance of Nile tilapia to waterborne heavy metals increased after feeding the fish with an activated carbon-supplemented feed for 8 weeks, thus reducing the accumulation of heavy metals in fish tissues [
30]. Likewise, the accumulation of heavy metals (Pb, Cd, and Cu) was minimized in
Huso huso muscle tissues after being fed with a diet supplemented with 15 g/kg of activated carbon for 60 days [
31]. In our study, selenium accumulation in the muscle tissue of juvenile blunt snout bream decreased with increasing dietary bamboo charcoal content. In contrast, another study reported that dietary supplementation with commercial charcoal had negligible effects on the protein, crude fat, and ash contents of red tilapia muscle tissue [
22]. Our study detected that the muscle protein content of blunt snout bream juveniles increased when the bamboo charcoal content in the feed was 2 g/kg and 3 g/kg.
Blood biochemical parameters such as ALB, ALT, AST, and ALP are common indicators of the physiological, liver health, and immune status of fish [
13,
32,
33]. Previous studies have reported that excess dietary Se (1.46 mg/kg) led to significantly lower ALB plasma levels in juvenile blunt snout bream, thus highlighting the negative effects of high Se dietary exposure on the immune response of these organisms [
7]. Our results indicate that the presence of bamboo charcoal (3 g/kg) in the Se-rich feed reduced ALP plasma levels and improved liver metabolism and detoxification capacity in juvenile blunt snout bream. Similarly, another study demonstrated that dietary supplementation with 25 g/kg of activated carbon in
Huso huso resulted in maximum ALB plasma levels and minimum ALT and AST plasma levels during a heavy metal (Pb, Cd, and Cu) stress challenge [
31]. This demonstrated that activated carbon can alleviate the negative effects of heavy metals on blood health. ALT, AST, and ALP plasma levels decreased drastically at bamboo charcoal levels of 0.5% and 1% in red tilapia after a 60 day feeding trial [
34]. In carp (
Cyprinus carpio L.), it was reported that the plasma levels of ALT, AST, and ALP were significantly lower when the bamboo charcoal content in the feed was 1–4% [
25]. Nevertheless, the optimal level of dietary bamboo charcoal or activated carbon may vary between studies. This can be attributed to species-specific differences, as well as carbon sources, and fish feeding habits.
The presence of 2% bamboo charcoal in diets with high deoxynivalenol concentrations significantly increased the activity of T-AOC and reduced the MDA content [
35]. Likewise, dietary supplementation with activated carbon can enhance the tolerance of tilapia to oxidative stress and reduce heavy metal uptake in these organisms [
36]. Our previous study detected that the activities of enzymes related to the antioxidant system were inhibited when the Se content in the feed was 1.46 mg/kg diet, suggesting that excessive dietary exposure to Se may chronically impair the functions of the antioxidant system [
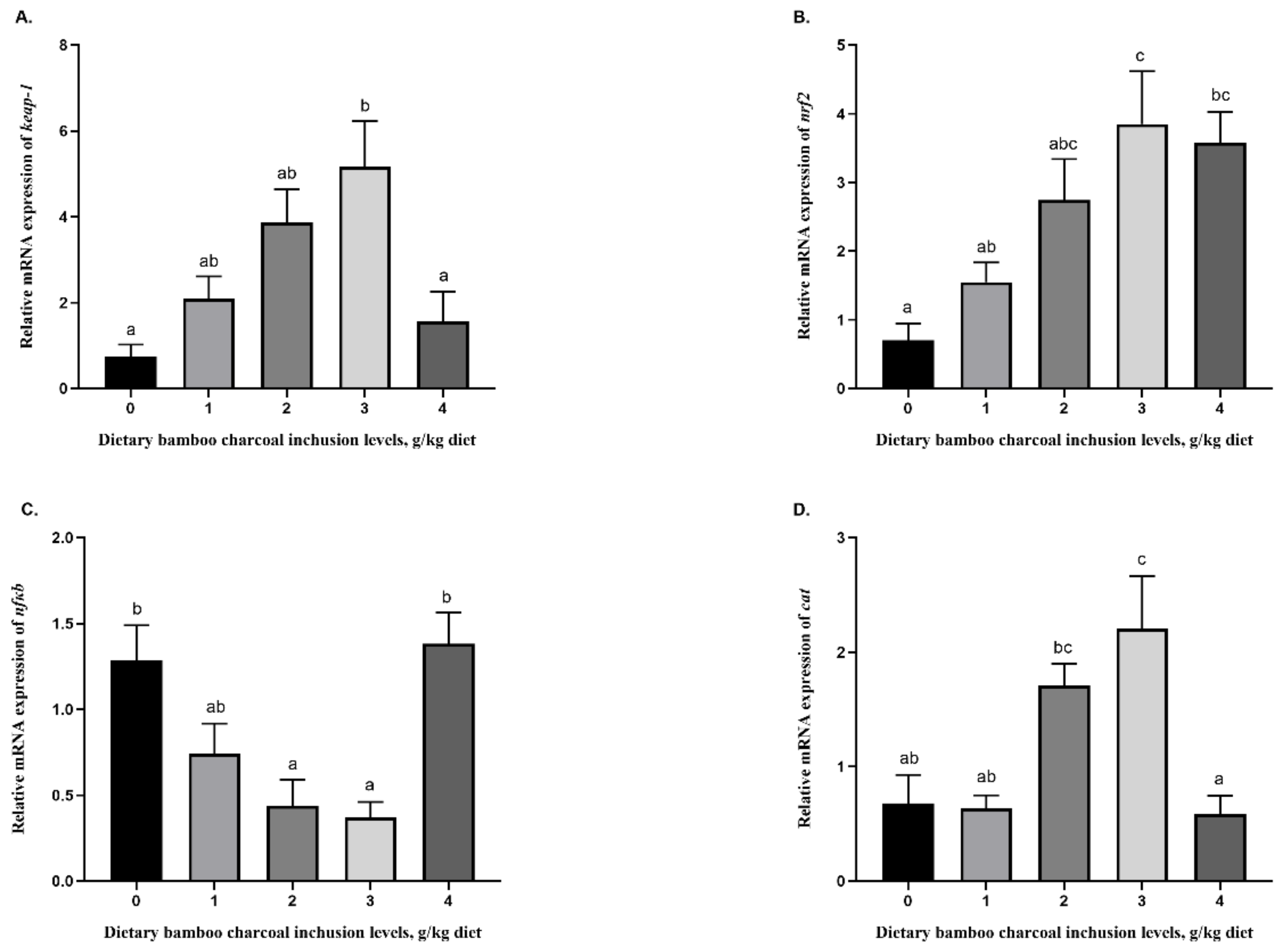
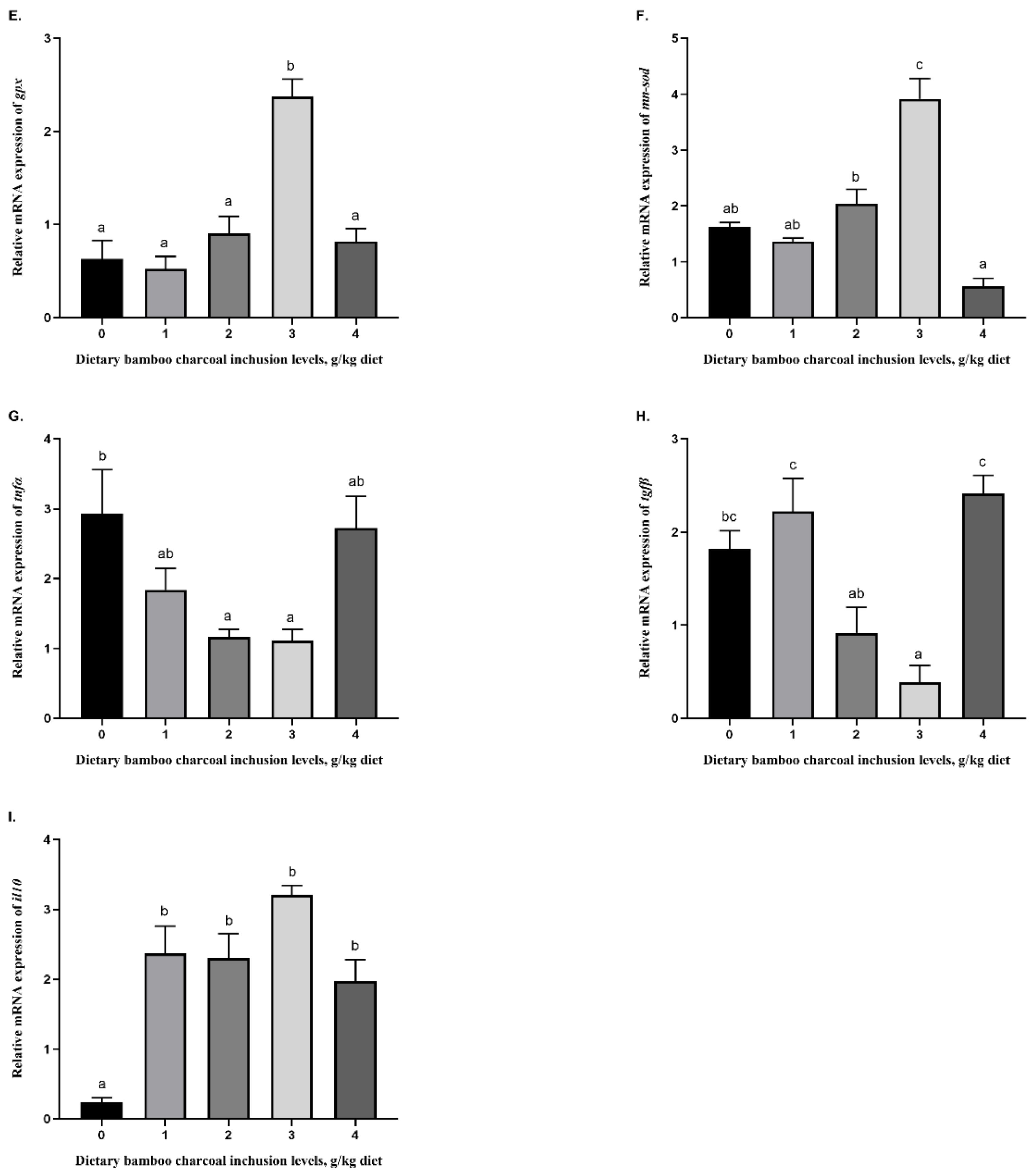
7]. While improvements in the MDA content were observed at 1 g/kg inclusion levels, upon close inspection, the data suggest that when considering an inclusion level that can produce the maximum beneficial effects on these hepatic antioxidant enzyme activities and MDA levels, and thus on the overall health of fish, an improvement in most parameters (CAT, T-SOD, T-AOC, GSH-Px, and GSH) can be observed at the 3 g/kg inclusion level compared to the control group. Therefore, BC supplementation at 3 g/kg inclusion can be applied in Se-rich diets of blunt snout bream to enhance the hepatic activities.
Nrf2 plays a central role in cell protection against a wide variety of stressors. Many studies have characterized the molecular sequences and regulatory functions of nrf2 in teleosts including grass carp (
Ctenopharyngodon idellus) [
37], zebrafish (
Danio rerio) [
38],
Pseudosciaena crocea [
39],
Cyprinus carpio var. Jian [
40], and
Coilia nasus [
41]. The Nrf2 signaling pathway is activated as a stress compensation mechanism under environmental or nutritional stress. For instance, nrf2 expression was elevated in isoniazid-induced liver injury in zebrafish larvae [
42]. Our previous study identified a link between nrf2 upregulation caused by oxidative stress and hyperglycemia in juvenile blunt snout bream (
Megalobrama amblycephala) [
43]. The Nrf2-Keap1 system is an evolutionarily conserved intracellular defense mechanism to counteract oxidative stress and to preserve cellular homeostasis [
44,
45,
46]. In stressful conditions, Nrf2 is activated, detaches from Keap1, and translocates to the nucleus to induce the expression of antioxidant and metabolic genes. Additionally, the Nrf2-Nfκb pathways regulate the physiological homeostasis of the cellular redox status, as well as stress and inflammation responses. Compounds that decrease the inflammatory response by suppressing NF-κB signaling are known to activate the Nrf2 pathway. Consistently, our study detected that 0 to 3 k/kg bamboo charcoal supplementation in Se-rich diets upregulated the mRNA expression of
keap1 and
nrf2, whereas
nfkb expression exhibited the opposite trend. It is widely accepted that the activities of antioxidant enzymes could partly result from the induction of their encoding genes to decrease the inflammatory response, which has been reported to be regulated by the Nrf2/Keap1/Nfκb signaling pathway in fish [
47,
48,
49]. In addition to the transcriptional changes of the aforementioned Nrf2/Keap1/Nfκb signaling genes, the gene expressions of the antioxidant enzymes
cat,
gpx, and
mn-sod were consistently enhanced in the group fed with the diet with 3 k/kg bamboo charcoal supplementation. Inflammation is generally a pervasive response to disturbances in tissue homeostasis due to a variety of stimuli, which involves the activation of innate and adaptive immunity. For instance, pro-inflammatory cytokines (e.g., tnfα, tgfβ, IL-1β, and IL-6) and anti-inflammatory cytokines (e.g., il10) have been described to play a critical role in the inflammatory process. Excessive dietary nutrients always lead to adverse effects on fish. For example, pro-inflammatory cytokines were upregulated and anti-inflammatory cytokines were downregulated in grass carp fed with excessive amounts of dietary histidine [
50]. Nevertheless, some feed additives could alleviate these adverse effects and even improve immunity. Song et al. [
51] described that dietary emodin supplementation upregulated the expression of the antioxidant gene
cat while inhibiting
Il-1β,
tnfα, and
tgfβ by modulating the Nrf2-Keap1 signaling pathway. In the early life stages of the zebrafish, the antioxidant enzymes SOD, CAT, and GPx were activated due to Nrf2 overexpression, thus preventing perfluorooctanesulfonic acid from inducing oxidative stress [
52]. Consistent with these results, the pro-inflammatory cytokines
tnfα and
tgfβ were inhibited in the 2–3 g/kg bamboo charcoal supplementation groups in our study, whereas the expression of the anti-inflammatory cytokine
il10 was upregulated in the bamboo charcoal supplementation groups compared with the Se-rich diet control group.







{kind=link}
{kind=link}