
Effects of Taurine on Sperm Quality during Room Temperature Storage in Hu Sheep

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Preparation of Semen Extender and Semen Collection
2.3. Dilution and Evaluation of Semen
2.3.1. Sperm Motility Parameters
2.3.2. Plasma Membrane Integrity
2.3.3. Acrosome Integrity
2.3.4. Seminal Oxidative Status Assessment
2.3.5. MMP
2.4. Statistical Analysis
3. Results
3.1. Effects of Tau Supplementation on Sperm Progressive Motility
3.2. Effects of Tau Supplementation on Sperm Kinetic Parameters
3.3. Effects of Tau Supplementation on Sperm Plasma Membrane Integrity
3.4. Effects of Tau Supplementation on Sperm Acrosome Integrity
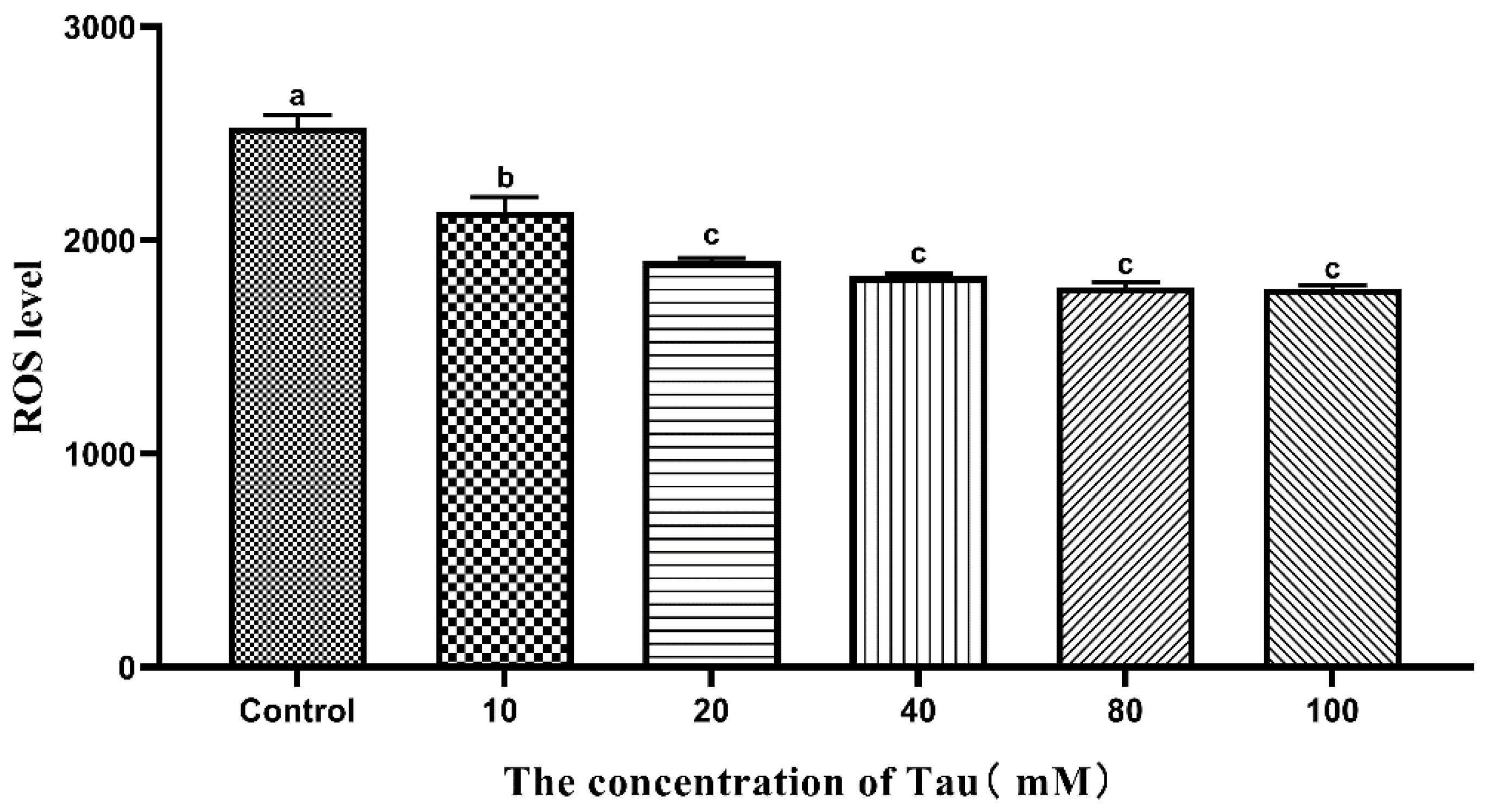
3.5. Effects of Tau Supplementation on Sperm ROS Content
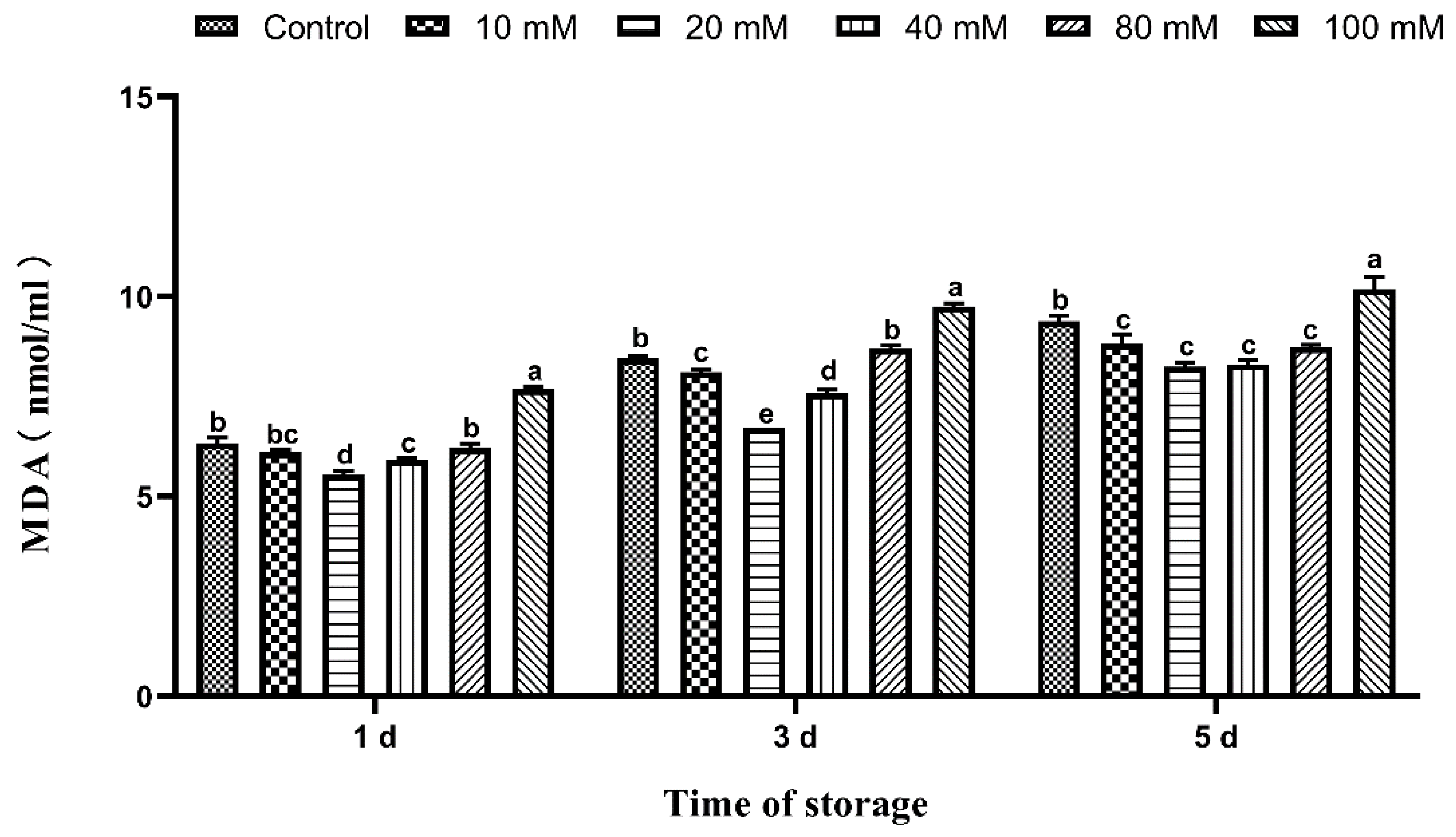
3.6. Effects of Tau Supplementation on Semen MDA Content
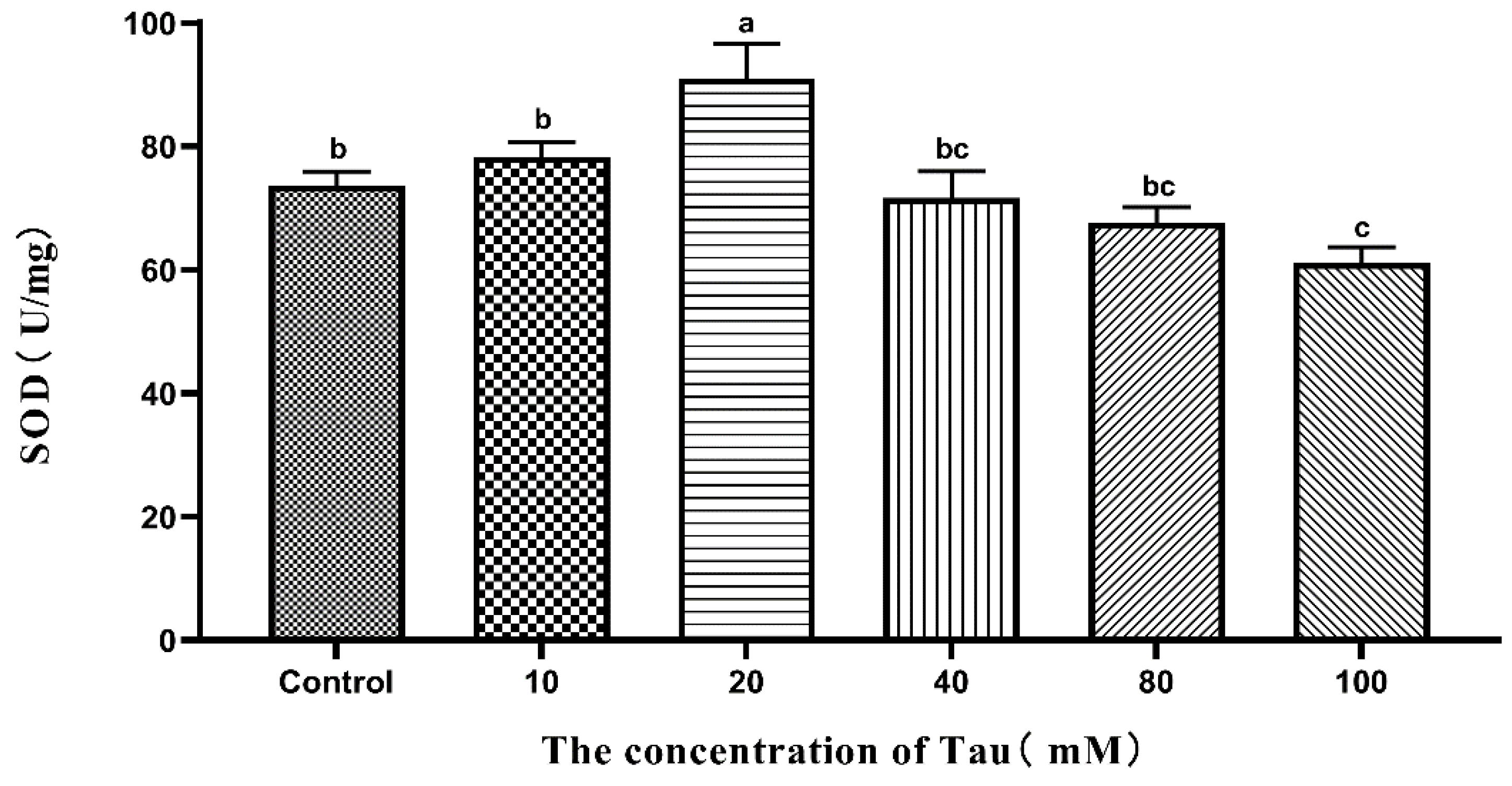
3.7. Effects of Tau Supplementation on Sperm SOD Activity
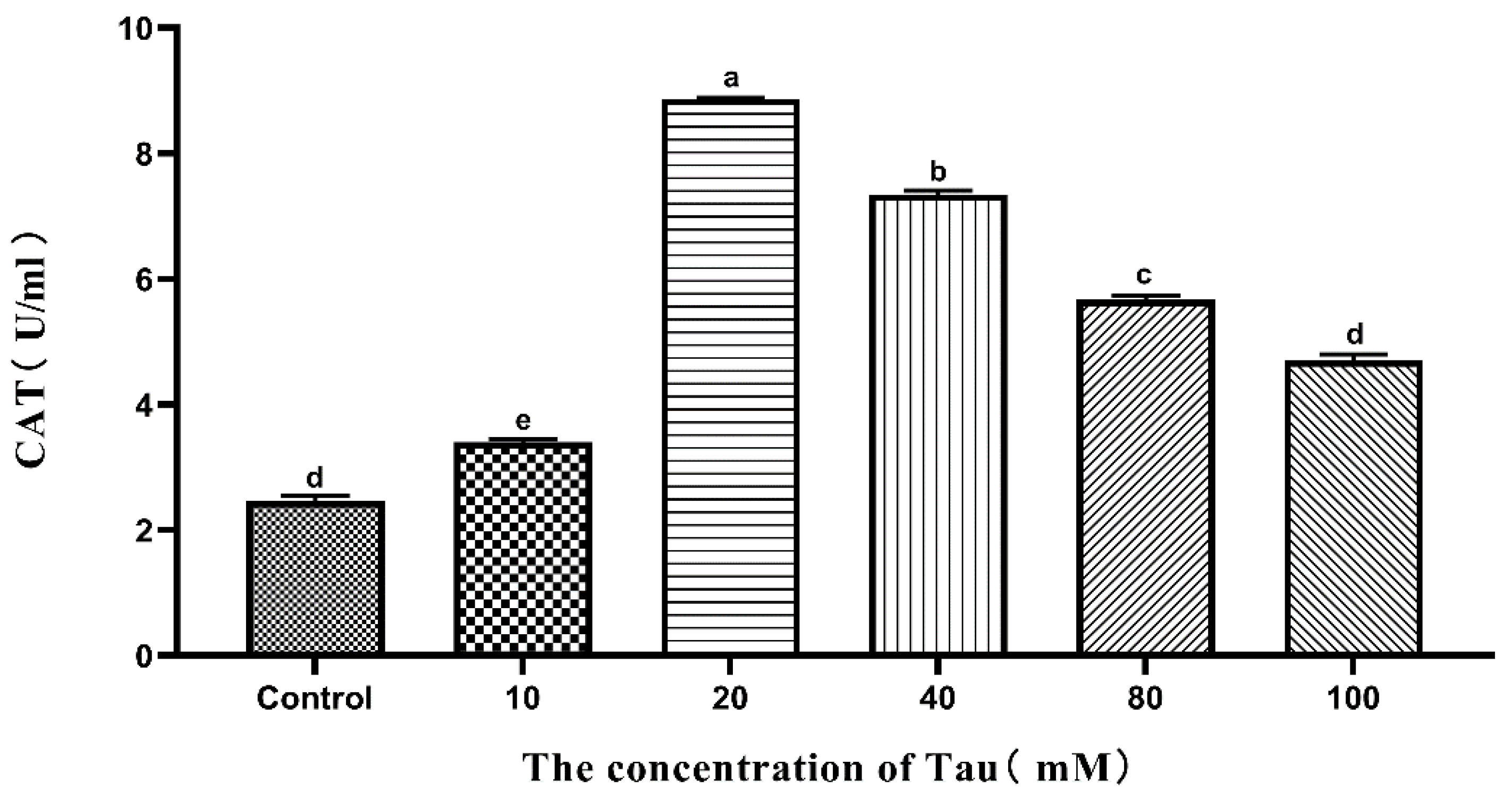
3.8. Effects of Tau Supplementation on Semen CAT Activity
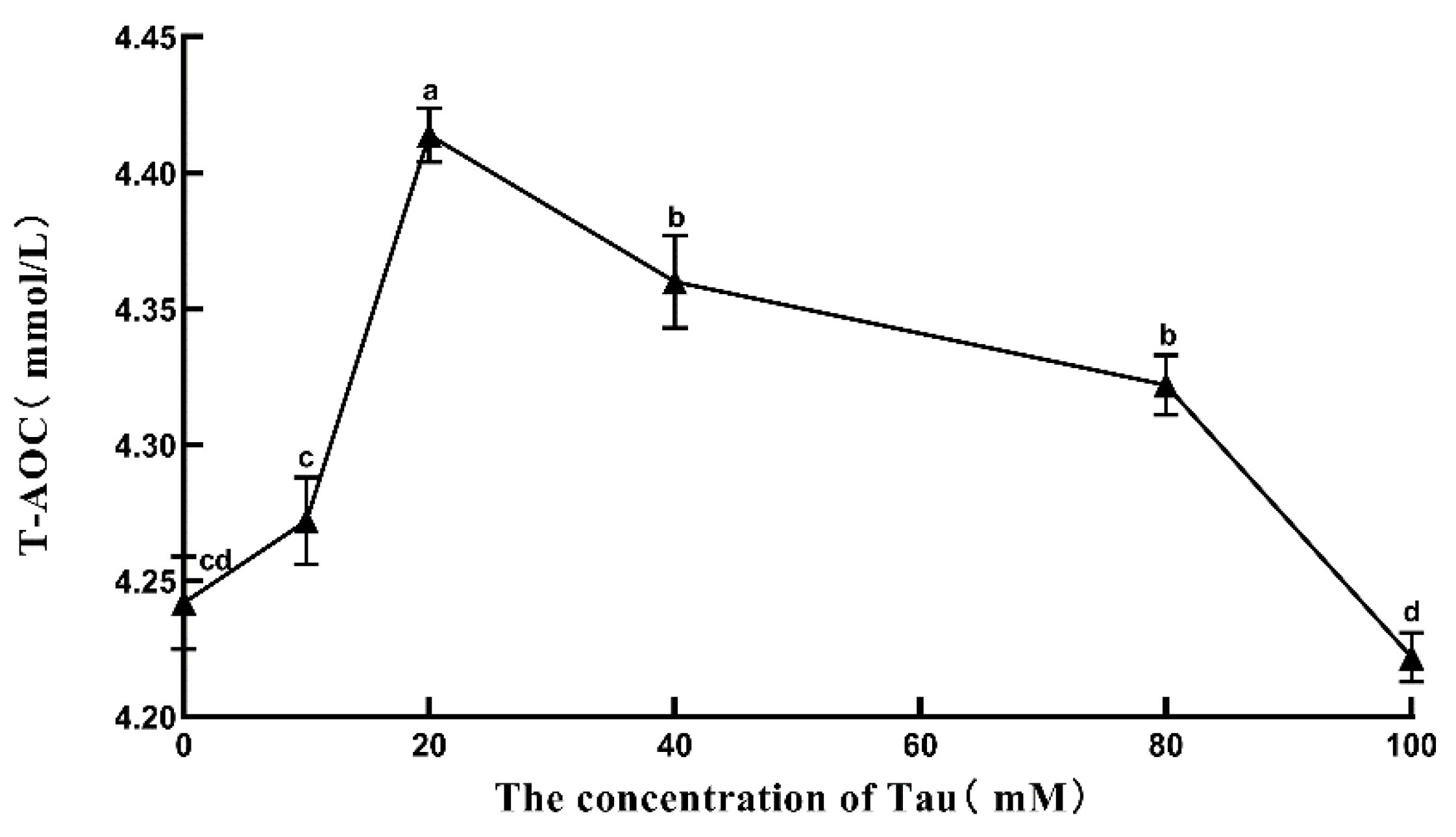
3.9. Effects of Tau Supplementation on Semen T-AOC
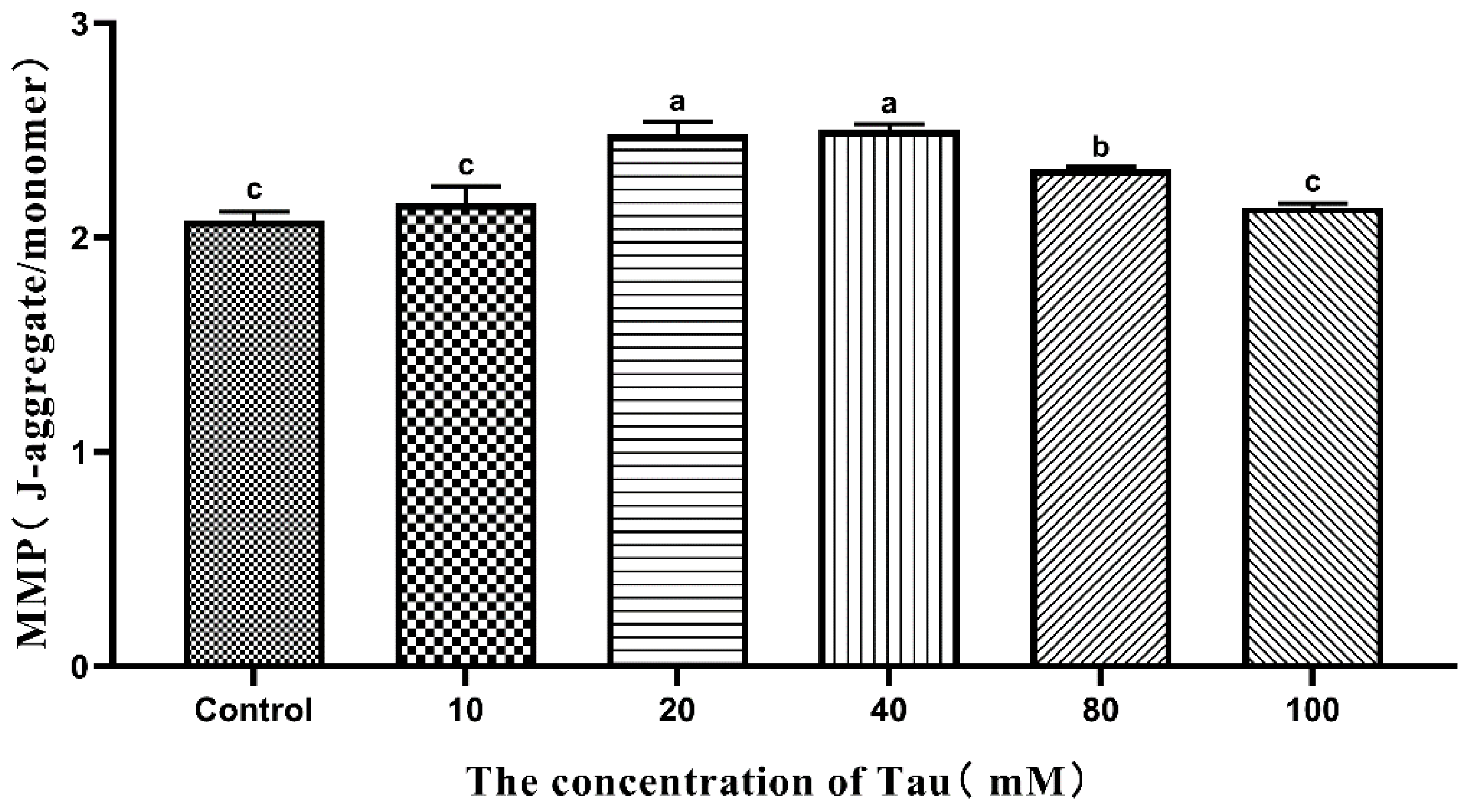
3.10. Effects of Tau Supplementation on Sperm MMP
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bucak, M.N.; Tekin, N. Protective effect of taurine, glutathione and trehalose on the liquid storage of ram semen. Small Rumin. Res. 2007, 73, 103–108. [Google Scholar] [CrossRef]
- Zhang, L.; Sohail, T.; Yongjun, L.I.; Yanhu, W.A.N.G.; Changjiang, C.H.U.; Yunkui, F.E.N.G.; Jinliang, M.A. Effect of Different Extenders on the Sperm Quality Parameters of Hu Ram Semen Preserved at 16 °C. Kafkas Univ. Vet. Fak. Derg. 2021, 27, 83–91. [Google Scholar]
- Goncalves, F.S.; Barretto, L.S.S.; Arruda, R.P.; Perri, S.H.V.; Mingoti, G.Z. Effect of Antioxidants During BovineIn VitroFertilization Procedures on Spermatozoa and Embryo Development. Reprod. Domest. Anim. 2010, 45, 129–135. [Google Scholar] [CrossRef]
- Agarwal, A.; Makker, K.; Sharma, R. Clinical Relevance of Oxidative Stress in Male Factor Infertility: An Update. Am. J. Reprod. Immunol. 2007, 59, 2–11. [Google Scholar] [CrossRef]
- Wathes, D.C.; Abayasekara, D.R.E.; Aitken, R.J. Polyunsaturated Fatty Acids in Male and Female Reproduction1. Biol. Reprod. 2007, 77, 190–201. [Google Scholar] [CrossRef]
- Tremellen, K. Oxidative stress and male infertility—A clinical perspective. Hum. Reprod. Updat. 2008, 14, 243–258. [Google Scholar] [CrossRef]
- Sawyer, D.E.; Mercer, B.G.; Wiklendt, A.M.; Aitken, R. Quantitative analysis of gene-specific DNA damage in human spermatozoa. Mutat. Res. Mol. Mech. Mutagen. 2003, 529, 21–34. [Google Scholar] [CrossRef]
- Beer-Ljubić, B.; Aladrović, J.; Marenjak, T.; Laškaj, R.; Majić-Balić, I.; Milinković-Tur, S. Cholesterol concentration in seminal plasma as a predictive tool for quality semen evaluation. Theriogenology 2009, 72, 1132–1140. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.T.; Wang, J.R.; Sun, L.Z.; Jin, X.H.; Shi, X.Y.; Lin, J.Y.; Yue, S.L.; Zhou, J.B. Effects of astaxanthin on plasma membrane function and fertility of boar sperm during cryopreservation. Theriogenology 2021, 164, 58–64. [Google Scholar] [CrossRef]
- Jong, C.J.; Azuma, J.; Schaffer, S. Mechanism underlying the antioxidant activity of taurine: Prevention of mitochondrial oxidant production. Amino Acids 2011, 42, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Lambert, I.H.; Jensen, J.V.; Pedersen, P.A. mTOR ensures increased release and reduced uptake of the organic osmolyte taurine under hypoosmotic conditions in mouse fibroblasts. Am. J. Physiol. Physiol. 2014, 306, C1028–C1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Puerta, C.; Arrieta, F.J.; Balsa, J.A.; Botella-Carretero, J.I.; Zamarrón, I.; Vázquez, C. Taurine and glucose metabolism: A review. Nutr. Hosp. 2010, 25, 910–919. [Google Scholar] [PubMed]
- Huxtable, R.J. Physiological actions of taurine. Physiol. Rev. 1992, 72, 101–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarıözkan, S.; Bucak, M.N.; Tuncer, P.B.; Ulutaş, P.A.; Bilgen, A. The influence of cysteine and taurine on microscopic–oxidative stress parameters and fertilizing ability of bull semen following cryopreservation. Cryobiology 2009, 58, 134–138. [Google Scholar] [CrossRef]
- Schaffer, S.W.; Jong, C.J.; Kc, R.; Azuma, J. Physiological roles of taurine in heart and muscle. J. Biomed. Sci. 2010, 17, S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seify, M.; Zarabadipour, M.; Ghaleno, L.R.; Alizadeh, A.; Valojerdi, M.R. The anti-oxidant roles of Taurine and Hypotaurine on acrosome integrity, HBA and HSPA2 of the human sperm during vitrification and post warming in two different temperature. Cryobiology 2019, 90, 89–95. [Google Scholar] [CrossRef]
- Banday, M.N.; Lone, F.A.; Rasool, F.; Rashid, M.; Shikari, A. Use of antioxidants reduce lipid peroxidation and improve quality of crossbred ram sperm during its cryopreservation. Cryobiology 2017, 74, 25–30. [Google Scholar] [CrossRef]
- Dorado, J.; Acha, D.; Ortiz, I.; Gálvez, M.; Carrasco, J.; Gómez-Arrones, V.; Calero-Carretero, R.; Hidalgo, M. Effect of extender and amino acid supplementation on sperm quality of cooled-preserved Andalusian donkey (Equus asinus) spermatozoa. Anim. Reprod. Sci. 2014, 146, 79–88. [Google Scholar] [CrossRef]
- Chhillar, S.; Singh, V.K.; Kumar, R.; Atreja, S.K. Effects of Taurine or Trehalose supplementation on functional competence of cryopreserved Karan Fries semen. Anim. Reprod. Sci. 2012, 135, 1–7. [Google Scholar] [CrossRef]
- Aly, H.A.; Khafagy, R.M. Taurine reverses endosulfan-induced oxidative stress and apoptosis in adult rat testis. Food Chem. Toxicol. 2014, 64, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, J.F.; Gadella, B.M. In situ detection and localization of lipid peroxidation in individual bovine sperm cells. Free Radic. Biol. Med. 2003, 35, 1382–1391. [Google Scholar] [CrossRef]
- Rai, N.K.; Ashok, A.; Rai, A.; Tripathi, S.; Nagar, G.K.; Mitra, K.; Bandyopadhyay, S. Exposure to As, Cd and Pb-mixture impairs myelin and axon development in rat brain, optic nerve and retina. Toxicol. Appl. Pharmacol. 2013, 273, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Prabakaran, S.; Allamaneni, S. What an andrologist/urologist should know about free radicals and why. Urology 2006, 67, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.; Ryan, A.L.; Baker, M.A.; McLaughlin, E.A. Redox activity associated with the maturation and capacitation of mammalian spermatozoa. Free. Radic. Biol. Med. 2004, 36, 994–1010. [Google Scholar] [CrossRef] [PubMed]
- Michael, A.; Alexopoulos, C.; Pontiki, E.; Hadjipavlou-Litina, D.; Saratsis, P.; Boscos, C. Effect of antioxidant supplementation on semen quality and reactive oxygen species of frozen-thawed canine spermatozoa. Theriogenology 2007, 68, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Foote, R.; Chen, Y.; Brockett, C.; Kaproth, M. Fertility of Bull Spermatozoa Frozen in Whole Milk Extender with Trehalose, Taurine, or Blood Serum. J. Dairy Sci. 1993, 76, 1908–1913. [Google Scholar] [CrossRef]
- Bucak, M.N.; Tuncer, P.B.; Sarıözkan, S.; Ulutaş, P.A.; Çoyan, K.; Başpınar, N.; Özkalp, B. Effects of hypotaurine, cysteamine and aminoacids solution on post-thaw microscopic and oxidative stress parameters of Angora goat semen. Res. Vet. Sci. 2009, 87, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Bucak, M.N.; Ateşşahin, A.; Varışlı, Ö.; Yüce, A.; Tekin, N.; Akçay, A. The influence of trehalose, taurine, cysteamine and hyaluronan on ram semen: Microscopic and oxidative stress parameters after freeze–thawing process. Theriogenology 2007, 67, 1060–1067. [Google Scholar] [CrossRef]
- Ijaz, A.; Ducharme, R. Effect of various extenders and taurine on survival of stallion sperm cooled to 5 degrees C. Theriogenology 1995, 44, 1039–1050. [Google Scholar] [CrossRef]
- Lone, F.A.; Naikoo, M.; Shah, S.M.; Darzi, S.A.; Farooq, J. Effect of Idebenone, Resveratrol and Taurine on the Sperm Quality and Lipid Peroxidation of Cryopreserved Crossbred Ram Semen. Cryoletters 2021, 42, 146–153. [Google Scholar]
- Rostami, B.; Ebrahimi, D.; Sadeghipanah, H.; Masoumi, R.; Shahir, M.H. Effects of supplementation of tris-egg yolk extender with different sugars and antioxidants on freezability of ram semen. Cryobiology 2020, 92, 62–66. [Google Scholar] [CrossRef]
- Rather, H.A.; Islam, R.; Malik, A.A.; Lone, F. Addition of antioxidants improves quality of ram spermatozoa during preservation at 4 °C. Small Rumin. Res. 2016, 141, 24–28. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.-G.; Fang, Q.; Liu, Q.; Du, R.-R.; Yang, G.-S.; Wang, L.-Q.; Hu, J.-H. Supplemental effect of different levels of taurine in Modena on boar semen quality during liquid preservation at 17 °C. Anim. Sci. J. 2017, 88, 1692–1699. [Google Scholar] [CrossRef] [PubMed]
- Stephens, T.D.; Brooks, R.M.; Carrington, J.L.; Cheng, L.; Carrington, A.C.; Porr, C.A.; Splan, R.K. Effects of Pentoxifylline, Caffeine, and Taurine on Post-Thaw Motility and Longevity of Equine Frozen Semen. J. Equine Vet. Sci. 2013, 33, 615–621. [Google Scholar] [CrossRef]
- McLaughlin, R.; Bowler, D.; Kelly, C.J.; Kay, E.; Bouchier-Hayes, D. Taurine protects against early and late skeletal muscle dysfunction secondary to ischaemia reperfusion injury. Eur. J. Surg. 2000, 166, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Awda, B.J.; Mackenzie-Bell, M.; Buhr, M.M. Reactive Oxygen Species and Boar Sperm Function1. Biol. Reprod. 2009, 81, 553–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (deltapsi(m)) in apoptosis; an update. Apoptosis 2003, 8, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Bilgeri, Y.R.; Winckelmann, A.; Berzin, M.; Lyons, G.; Reinach, S.G. Adenosine Triphosphate Levels in Human Spermatozoa. Arch. Androl. 1987, 18, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Partyka, A.; Rodak, O.; Bajzert, J.; Kochan, J.; Niżański, W. The Effect of L-Carnitine, Hypotaurine, and Taurine Supplementation on the Quality of Cryopreserved Chicken Semen. BioMed Res. Int. 2017, 2017, 7279341. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time of Storage (d) | Progressive Motility (%) | |||||
|---|---|---|---|---|---|---|
| Control | 10 mM | 20 mM | 40 mM | 80 mM | 100 mM | |
| 0 | 86.09 ± 1.24 | 85.93 ± 1.24 | 86.26 ± 0.75 | 85.94 ± 1.18 | 86.17 ± 1.28 | 86.13 ± 1.12 |
| 1 | 80.92 ± 1.39 | 80.04 ± 1.23 | 81.70 ± 1.16 | 81.28 ± 1.86 | 81.31 ± 0.65 | 76.72 ± 2.16 |
| 3 | 67.79 ± 0.79 d | 69.31 ± 0.90 cd | 74.41 ± 0.92 a | 72.00 ± 0.68 ab | 70.48 ± 0.39 bc | 63.35 ± 1.06 e |
| 5 | 52.93 ± 0.71 de | 54.64 ± 0.23 d | 65.49 ± 0.67 a | 60.75 ± 0.41 b | 58.11 ± 0.67 c | 51.64 ± 0.57 e |
| 7 | 24.09 ± 0.43 cd | 25.59 ± 0.25 c | 34.79 ± 0.74 a | 30.80 ± 0.35 b | 29.68 ± 0.48 b | 23.67 ± 0.61 d |
| Index | Time of Storage (d) | Control | 10 mM | 20 mM | 40 mM | 80 mM | 100 mM |
|---|---|---|---|---|---|---|---|
| VSL (μm/s) | 0 d | 38.47 ± 1.48 | 38.24 ± 0.52 | 35.92 ± 0.65 | 36.06 ± 0.74 | 36.59 ± 0.10 | 36.85 ± 0.33 |
| 1 d | 35.14 ± 0.83 | 33.22 ± 0.87 | 33.88 ± 0.19 | 34.63 ± 1.02 | 33.72 ± 0.74 | 34.62 ± 0.33 | |
| 3 d | 32.40 ± 0.56 a | 30.86 ± 0.56 ab | 32.25 ± 0.70 a | 31.65 ± 0.81 ab | 31.91 ± 0.69 ab | 29.25 ± 1.49 b | |
| 5 d | 28.92 ± 1.63 | 30.45 ± 0.89 | 30.75 ± 0.49 | 30.85 ± 0.51 | 27.48 ± 0.63 | 28.51 ± 2.53 | |
| 7 d | 23.11 ± 0.73 | 20.60 ± 0.52 | 22.14 ± 0.31 | 22.20 ± 1.19 | 21.78 ± 0.41 | 21.92 ± 1.05 | |
| VCL (μm/s) | 0 d | 68.04 ± 4.13 | 65.85 ± 1.64 | 64.09 ± 2.58 | 65.07 ± 1.00 | 63.53 ± 2.77 | 65.85 ± 0.92 |
| 1 d | 63.51 ± 0.89 | 60.16 ± 1.88 | 60.99 ± 1.15 | 62.02 ± 0.83 | 61.29 ± 1.45 | 62.17 ± 1.39 | |
| 3 d | 63.16 ± 1.01 a | 60.89 ± 0.64 ab | 64.41 ± 0.63 a | 63.23 ± 1.11 a | 63.41 ± 1.41 a | 56.94 ± 2.75 b | |
| 5 d | 57.15 ± 2.65 | 61.94 ± 2.10 | 62.83 ± 0.68 | 62.77 ± 0.24 | 52.75 ± 1.99 | 55.61 ± 7.30 | |
| 7 d | 39.09 ± 1.35 ab | 35.62 ± 1.22 b | 39.89 ± 2.07 ab | 40.14 ± 1.47 ab | 40.42 ± 0.84 ab | 41.89 ± 2.00 a | |
| VAP (μm/s) | 0 d | 48.11 ± 2.92 | 46.56 ± 1.16 | 45.32 ± 1.83 | 46.01 ± 0.71 | 44.92 ± 1.95 | 46.56 ± 0.65 |
| 1 d | 44.91 ± 0.63 | 42.54 ± 1.33 | 43.13 ± 0.82 | 43.85 ± 0.58 | 43.33 ± 1.02 | 43.96 ± 0.98 | |
| 3 d | 44.66 ± 0.71 a | 43.06 ± 0.46 ab | 45.55 ± 0.44 a | 44.71 ± 0.78 a | 44.84 ± 1.00 a | 40.26 ± 1.94 b | |
| 5 d | 40.42 ± 1.87 | 43.80 ± 1.48 | 44.43 ± 0.49 | 44.39 ± 0.17 | 37.30 ± 1.41 | 39.32 ± 5.16 | |
| 7 d | 27.64 ± 0.96 ab | 25.18 ± 0.87 b | 28.21 ± 1.47 ab | 28.38 ± 1.04 ab | 28.58 ± 0.59 ab | 29.62 ± 1.42 a | |
| ALH (μm) | 0 d | 19.93 ± 1.21 | 19.29 ± 0.48 | 18.77 ± 0.76 | 19.06 ± 0.29 | 18.61 ± 0.81 | 19.29 ± 0.27 |
| 1 d | 18.60 ± 0.26 | 17.62 ± 0.55 | 17.86 ± 0.34 | 18.16 ± 0.24 | 17.95 ± 0.42 | 18.21 ± 0.41 | |
| 3 d | 18.50 ± 0.29 a | 17.83 ± 0.19 ab | 18.87 ± 0.18 a | 18.52 ± 0.32 a | 18.57 ± 0.41 a | 16.68 ± 0.81 b | |
| 5 d | 16.74 ± 0.77 | 18.14 ± 0.61 | 18.40 ± 0.20 | 18.39 ± 0.07 | 15.45 ± 0.58 | 16.29 ± 2.14 | |
| 7 d | 11.45 ± 0.40 ab | 10.43 ± 0.36 b | 11.68 ± 0.61 ab | 11.76 ± 0.43 ab | 11.84 ± 0.25 ab | 12.27 ± 0.59 a | |
| MAD (°/s) | 0 d | 113.99 ± 2.79 | 104.76 ± 9.97 | 100.01 ± 6.39 | 110.36 ± 3.32 | 97.39 ± 15.22 | 113.28 ± 10.41 |
| 1 d | 64.92 ± 0.62 | 67.73 ± 4.37 | 69.02 ± 2.80 | 75.19 ± 10.97 | 78.67 ± 1.08 | 71.13 ± 2.95 | |
| 3 d | 57.78 ± 2.89 b | 66.97 ± 2.17 ab | 71.98 ± 5.74 a | 67.02 ± 2.59 ab | 68.03 ± 3.21 ab | 58.33 ± 1.44 b | |
| 5 d | 49.78 ± 2.28 cd | 53.41 ± 1.08 bc | 63.79 ± 4.16 a | 58.37 ± 2.35 ab | 55.78 ± 1.19 bc | 42.19 ± 2.41 d | |
| 7 d | 22.69 ± 2.36 c | 25.86 ± 1.54 bc | 27.62 ± 0.93 b | 32.83 ± 0.98 a | 32.46 ± 1.10 a | 24.43 ± 1.22 bc |
| Time of Storage (d) | Plasma Membrane Integrity (%) | |||||
|---|---|---|---|---|---|---|
| Control | 10 mM | 20 mM | 40 mM | 80 mM | 100 mM | |
| 0 | 73.38 ± 1.04 | 75.14 ± 0.58 | 74.39 ± 0.35 | 72.53 ± 0.61 | 73.41 ± 0.28 | 72.71 ± 0.20 |
| 1 | 65.58 ± 0.89 a | 65.14 ± 0.39 a | 65.37 ± 0.18 a | 63.07 ± 0.91 ab | 62.20 ± 0.72 b | 50.61 ± 1.18 c |
| 3 | 56.38 ± 0.82 c | 61.66 ± 1.68 ab | 64.91 ± 0.57 a | 61.86 ± 0.73 ab | 60.37 ± 0.53 b | 50.18 ± 1.50 d |
| 5 | 47.29 ± 1.90 d | 49.79 ± 0.43 cd | 57.08 ± 0.48 a | 54.03 ± 0.45 b | 50.42 ± 0.12 c | 42.64 ± 0.86 e |
| 7 | 42.59 ± 0.93 c | 44.21 ± 0.73 c | 53.88 ± 0.08 a | 47.76 ± 0.71 b | 43.15 ± 0.35 c | 36.88 ± 0.83 d |
| Time of Storage (d) | Acrosomal Integrity (%) | |||||
|---|---|---|---|---|---|---|
| Control | 10 mM | 20 mM | 40 mM | 80 mM | 100 mM | |
| 0 | 95.17 ± 1.35 | 95.72 ± 1.01 | 93.49 ± 0.26 | 93.91 ± 0.25 | 93.60 ± 0.83 | 94.20 ± 0.62 |
| 1 | 92.60 ± 0.32 a | 91.05 ± 0.33 bc | 93.10 ± 0.23 a | 90.57 ± 0.17 c | 92.09 ± 0.69 ab | 87.39 ± 0.65 d |
| 3 | 85.76 ± 0.85 d | 87.27 ± 0.33 c | 91.74 ± 0.43 a | 90.12 ± 0.46 b | 87.95 ± 0.08 c | 83.14 ± 0.42 e |
| 5 | 83.93 ± 0.08 d | 85.25 ± 0.31 bc | 88.93 ± 0.36 a | 85.89 ± 0.23 b | 84.23 ± 0.22 cd | 81.97 ± 0.59 e |
| 7 | 79.66 ± 0.93 cd | 81.62 ± 0.80 bc | 85.88 ± 0.32 a | 83.28 ± 0.42 b | 81.06 ± 1.09 bcd | 78.92 ± 0.27 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Wang, Y.; Sohail, T.; Kang, Y.; Niu, H.; Sun, X.; Ji, D.; Li, Y. Effects of Taurine on Sperm Quality during Room Temperature Storage in Hu Sheep. Animals 2021, 11, 2725. https://doi.org/10.3390/ani11092725
Zhang L, Wang Y, Sohail T, Kang Y, Niu H, Sun X, Ji D, Li Y. Effects of Taurine on Sperm Quality during Room Temperature Storage in Hu Sheep. Animals. 2021; 11(9):2725. https://doi.org/10.3390/ani11092725
Chicago/Turabian StyleZhang, Liuming, Yanhu Wang, Tariq Sohail, Yan Kang, Haoyuan Niu, Xiaomei Sun, Dejun Ji, and Yongjun Li. 2021. "Effects of Taurine on Sperm Quality during Room Temperature Storage in Hu Sheep" Animals 11, no. 9: 2725. https://doi.org/10.3390/ani11092725
APA StyleZhang, L., Wang, Y., Sohail, T., Kang, Y., Niu, H., Sun, X., Ji, D., & Li, Y. (2021). Effects of Taurine on Sperm Quality during Room Temperature Storage in Hu Sheep. Animals, 11(9), 2725. https://doi.org/10.3390/ani11092725