
No Evidence for Contagious Yawning in Juvenile Ravens (Corvus corax): An Observational Study

 ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
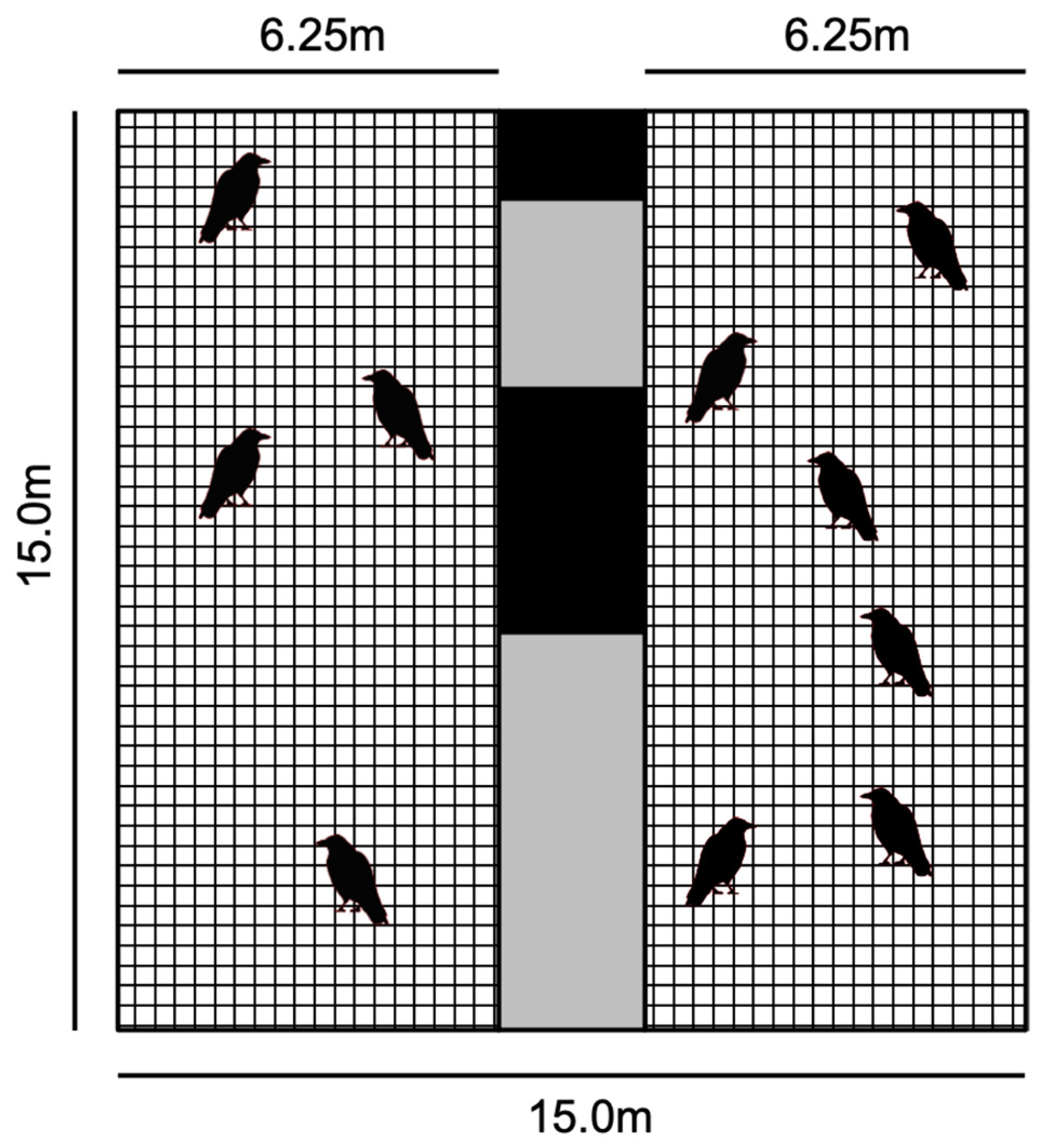
2.1. Subjects
2.2. Procedure
2.3. Behavioral Coding
2.4. Analysis
3. Results
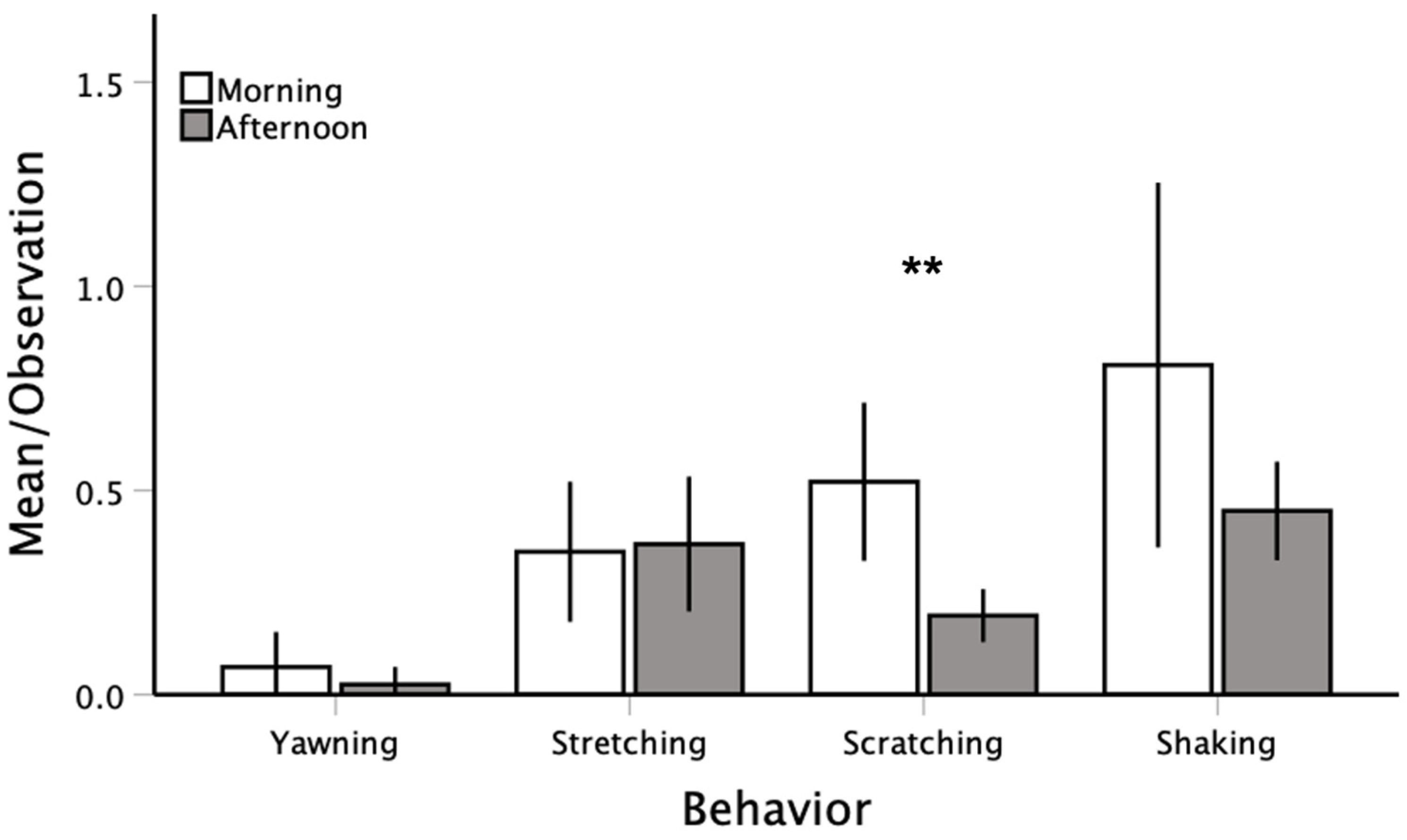
3.1. Descriptive Analyses and Circadian Effects
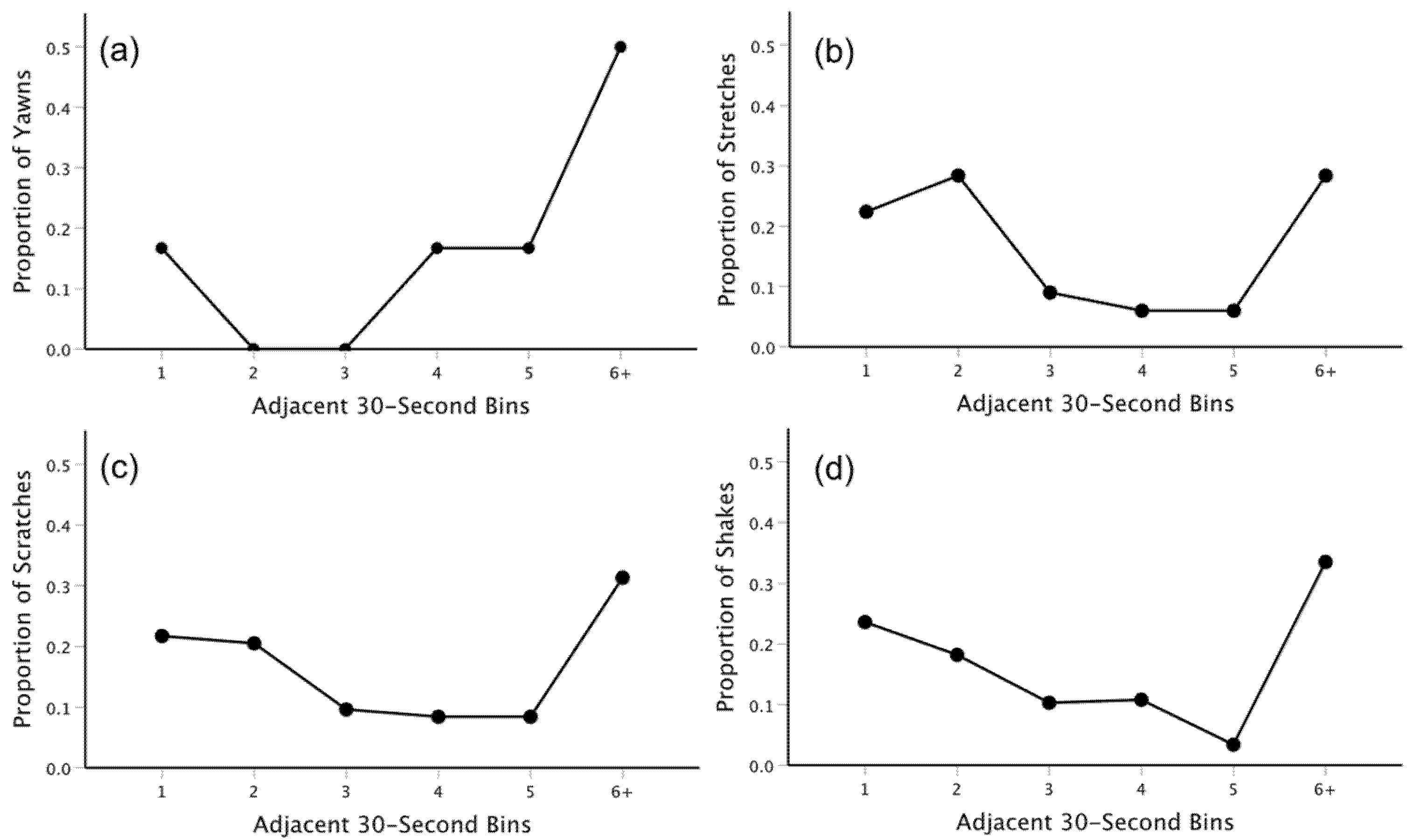
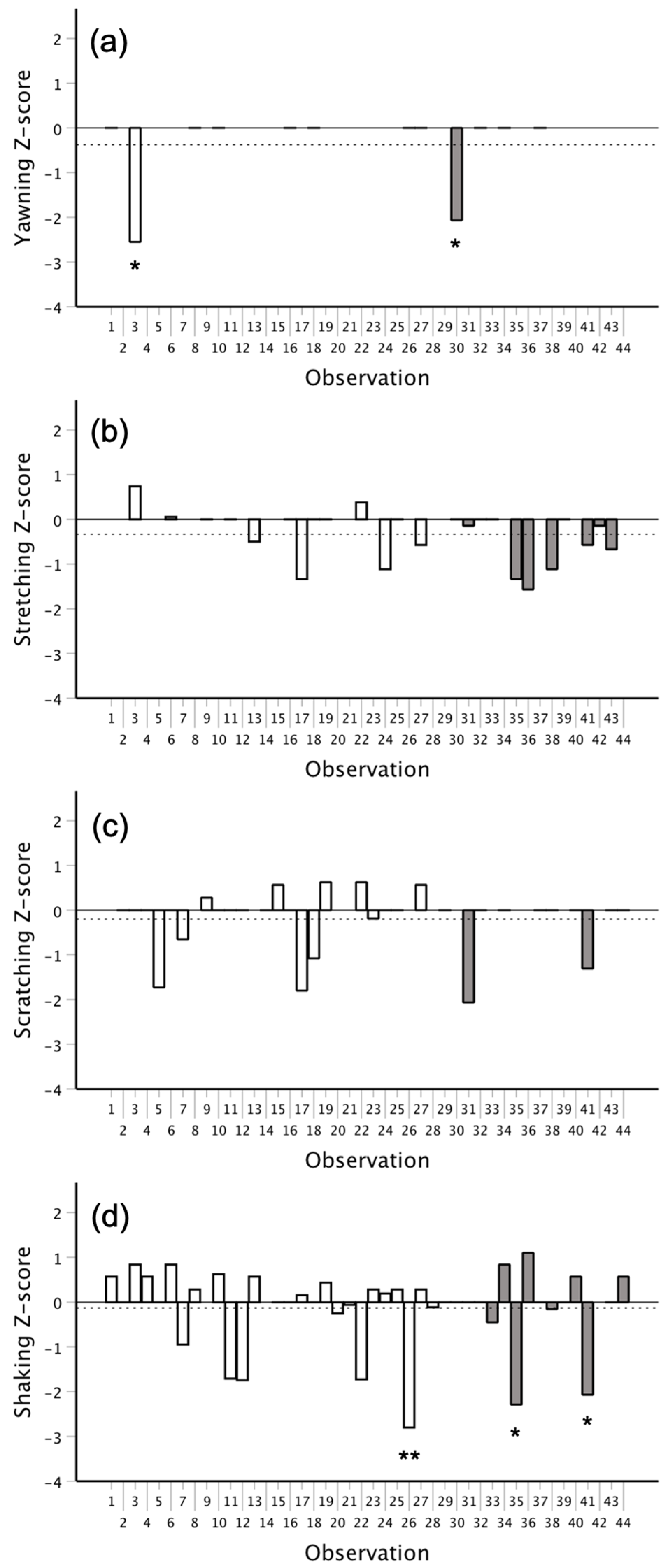
3.2. Temporal Distribution and Contagion Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zentall, T.R. Imitation by animals: How do they do it? Curr. Dir. Psychol. Sci. 2003, 12, 91–95. [Google Scholar] [CrossRef]
- Duranton, C.; Gaunet, F. Behavioural synchronization from an ethological perspective: Overview of its adaptive value. Adapt. Behav. 2016, 24, 181–191. [Google Scholar] [CrossRef]
- Casetta, G.; Nolfo, A.P.; Palagi, E. Yawn contagion promotes motor synchrony in wild lions, Panthera leo. Anim. Behav. 2021, 174, 149–159. [Google Scholar] [CrossRef]
- Gallup, A.C.; Meyers, K. Seeing others yawn selectively enhances vigilance: An eye-tracking study of snake detection. Anim. Cogn. 2021, 24, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Gallup, A.C. The causes and consequences of yawning in animal groups. Anim. Behav. 2022, 187, 209–219. [Google Scholar] [CrossRef]
- Anderson, J.R.; Myowa–Yamakoshi, M.; Matsuzawa, T. Contagious yawning in chimpanzees. Proc. Royal Soc. B 2004, 271 (Suppl. S6), S468–S470. [Google Scholar] [CrossRef] [Green Version]
- De Waal, F.B.; Preston, S.D. Mammalian empathy: Behavioural manifestations and neural basis. Nat. Rev. Neurosci. 2017, 18, 498–509. [Google Scholar] [CrossRef]
- Palagi, E.; Celeghin, A.; Tamietto, M.; Winkielman, P.; Norscia, I. The neuroethology of spontaneous mimicry and emotional contagion in human and non-human animals. Neurosci. Biobehav. Rev. 2020, 111, 149–165. [Google Scholar] [CrossRef]
- Prochazkova, E.; Kret, M.E. Connecting minds and sharing emotions through mimicry: A neurocognitive model of emotional contagion. Neurosci. Biobehav. Rev. 2017, 80, 99–114. [Google Scholar] [CrossRef]
- Ostner, J.; Wilken, J.; Schülke, O. Social contagion of affiliation in female macaques. R. Soc. Open Sci. 2021, 8, 201538. [Google Scholar] [CrossRef]
- Southern, W.E. Copulatory wing-flagging: A synchronizing stimulus for nesting ring-billed gulls. Bird-Band. 1974, 45, 210–216. [Google Scholar] [CrossRef]
- Stevens, E.F. Flamingo breeding: The role of group displays. Zoo Biol. 1991, 10, 53–63. [Google Scholar] [CrossRef]
- Palestis, B.G.; Burger, J. Evidence for social facilitation of preening in the common tern. Anim. Behav. 1988, 56, 1107–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppitt, W.; Blackburn, L.; Laland, K.N. Response facilitation in the domestic fowl. Anim. Behav. 2007, 73, 229–238. [Google Scholar] [CrossRef]
- Beauchamp, G. Collective waves of sleep in gulls (Larus spp.). Ethology 2011, 117, 326–331. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Díaz, M.; Grim, T.; Ibáñez-Álamoz, J.D.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.S.; Tatte, K.; Marko, G.; Jiang, Y.; et al. Contagious fear: Escape behavior increases with flock size in European gregarious birds. Ecol. Evol. 2019, 9, 6096–6104. [Google Scholar] [CrossRef]
- Provine, R.R. Yawning as a stereotyped action pattern and releasing stimulus. Ethology 1986, 72, 109–122. [Google Scholar] [CrossRef]
- Provine, R.R. Yawning: The yawn is primal, unstoppable and contagious, revealing the evolutionary and neural basis of empathy and unconscious behavior. Am. Sci. 2005, 93, 532–539. [Google Scholar] [CrossRef]
- Massen, J.J.M.; Church, A.M.; Gallup, A.C. Auditory contagious yawning in humans: An investigation into affiliation and status effects. Front. Psychol. 2015, 6, 1735. [Google Scholar] [CrossRef] [Green Version]
- Massen, J.J.M.; Vermunt, D.A.; Sterck, E.H. Male yawning is more contagious than female yawning among chimpanzees (Pan troglodytes). PLoS ONE 2012, 7, e40697. [Google Scholar] [CrossRef] [Green Version]
- Campbell, M.W.; Cox, C.R. Observational data reveal evidence and parameters of contagious yawning in the behavioral repertoire of captive-reared chimpanzees (Pan troglodytes). Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.; Ariely, D.; Hare, B. Bonobos respond prosocially toward members of other groups. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Berlo, E.; Díaz-Loyo, A.P.; Juárez-Mora, O.E.; Kret, M.E.; Massen, J.J.M. Experimental evidence for yawn contagion in orangutans (Pongo pygmaeus). Sci. Rep. 2020, 10, 1–11. [Google Scholar]
- Moyaho, A.; Rivas-Zamudio, X.; Ugarte, A.; Eguibar, J.R.; Valencia, J. Smell facilitates auditory contagious yawning in stranger rats. Anim. Cogn. 2015, 18, 279–290. [Google Scholar] [CrossRef]
- Joly-Mascheroni, R.M.; Senju, A.; Shepherd, A.J. Dogs catch human yawns. Biol. Lett. 2008, 4, 446–448. [Google Scholar] [CrossRef] [Green Version]
- Harr, A.L.; Gilbert, V.R.; Phillips, K.A. Do dogs (Canis familiaris) show contagious yawning? Anim. Cogn. 2009, 12, 833–837. [Google Scholar] [CrossRef]
- Palagi, E.; Leone, A.; Mancini, G.; Ferrari, P.F. Contagious yawning in gelada baboons as a possible expression of empathy. Proc. Natl. Acad. Sci. USA 2009, 106, 19262–19267. [Google Scholar] [CrossRef] [Green Version]
- Romero, T.; Ito, M.; Saito, A.; Hasegawa, T. Social modulation of contagious yawning in wolves. PLoS ONE 2014, 9, e105963. [Google Scholar] [CrossRef] [Green Version]
- Norscia, I.; Coco, E.; Robino, C.; Chierto, E.; Cordoni, G. Yawn contagion in domestic pigs (Sus scrofa). Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Rossman, Z.T.; Padfield, C.; Young, D.; Hart, B.L.; Hart, L.A. Contagious yawning in African elephants (Loxodonta africana): Responses to other elephants and familiar humans. Front. Vet. Sci. 2020, 7, 252. [Google Scholar] [CrossRef]
- Wojczulanis-Jakubas, K.; Plenzler, J.; Jakubas, D. Indications of contagious behaviours in the southern elephant seal: An observational study. Behaviour 2019, 156, 59–77. [Google Scholar] [CrossRef]
- Yonezawa, T.; Sato, K.; Uchida, M.; Matsuki, N.; Yamazaki, A. Presence of contagious yawning in sheep. Anim. Sci. J. 2017, 88, 195–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massen, J.J.M.; Šlipogor, V.; Gallup, A.C. An observational investigation of behavioral contagion in common marmosets (Callithrix jacchus): Indications for contagious scent-marking. Front. Psychol. 2016, 7, 1190. [Google Scholar] [CrossRef] [Green Version]
- Reddy, R.B.; Krupenye, C.; MacLean, E.L.; Hare, B. No evidence for contagious yawning in lemurs. Anim. Cogn. 2016, 19, 889–898. [Google Scholar] [CrossRef] [Green Version]
- Palagi, E.; Norscia, I.; Cordoni, G. Lowland gorillas (Gorilla gorilla gorilla) failed to respond to others’ yawn: Experimental and naturalistic evidence. J. Comp. Psychol. 2019, 133, 406. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.L.; Gallup, A.C.; Vogel, A.R.; Vicario, S.M.; Clark, A.B. Evidence for contagious behaviors in budgerigars (Melopsittacus undulatus): An observational study of yawning and stretching. Behav. Processes 2012, 89, 264–270. [Google Scholar] [CrossRef]
- Gallup, A.C.; Swartwood, L.; Militello, J.; Sackett, S. Experimental evidence of contagious yawning in budgerigars (Melopsittacus undulatus). Anim. Cogn. 2015, 18, 1051–1058. [Google Scholar] [CrossRef]
- Bugnyar, T. Social cognition in ravens. Comp Cogn. Behav. Rev. 2013, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Boucherie, P.H.; Loretto, M.C.; Massen, J.J.M.; Bugnyar, T. What constitutes “social complexity” and “social intelligence” in birds? Lessons from ravens. Behav. Ecol. Sociobiol. 2019, 73, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Massen, J.J.M.; Pašukonis, A.; Schmidt, J.; Bugnyar, T. Ravens notice dominance reversals among conspecifics within and outside their social group. Nat. Comm. 2014, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, B.; Kaye, D.; Knight, T.; Schaumburg, K. Dispersal and association among common ravens. Condor 1994, 545–551. [Google Scholar] [CrossRef]
- Heinrich, B. Winter foraging at carcasses by three sympatric corvids, with emphasis on recruitment by the raven, Corvus corax. Behav. Ecol. Sociobiol. 1988, 23, 141–156. [Google Scholar] [CrossRef]
- Massen, J.J.M.; Ritter, C.; Bugnyar, T. Tolerance and reward equity predict cooperation in ravens (Corvus corax). Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Massen, J.J.M.; Schaake, W.A.A.; Bugnyar, T. Chimpanzees in Context; Hopper, L., Ross, S., Eds.; University of Chicago Press: Chicago, MI, USA, 2020; pp. 368–390. [Google Scholar]
- Osvath, M.; Sima, M. Sub-adult ravens synchronize their play: A case of emotional contagion. Anim. Behav. Cogn. 2014, 2, 197. [Google Scholar] [CrossRef]
- Wenig, K.; Pacher, L.; Bugnyar, T. Testing the contagious nature of allopreening: Bystander ravens are affected by conspecifics’ affiliative interactions. Anim. Behav. 2022, 184, 71–80. [Google Scholar] [CrossRef]
- Massen, J.J.M.; Gallup, A.C. Why contagious yawning does not (yet) equate to empathy. Neurosci. Biobehav. Rev. 2017, 80, 573–585. [Google Scholar] [CrossRef]
- Gallup, A.C. On the link between emotional contagion and contagious yawning. Neurosci. Biobehav. Rev. 2021, 121, 18–19. [Google Scholar] [CrossRef]
- Adriaense, J.E.; Martin, J.S.; Schiestl, M.; Lamm, C.; Bugnyar, T. Negative emotional contagion and cognitive bias in common ravens (Corvus corax). Proc. Natl. Acad. Sci. USA 2019, 116, 11547–11552. [Google Scholar] [CrossRef] [Green Version]
- Wenig, K.; Boucherie, P.H.; Bugnyar, T. Early evidence for emotional play contagion in juvenile ravens. Anim. Cogn. 2021, 24, 717–729. [Google Scholar] [CrossRef]
- Massen, J.J.M.; Szipl, G.; Spreafico, M.; Bugnyar, T. Ravens intervene in others’ bonding attempts. Curr. Biol. 2014, 24, 2733–2736. [Google Scholar] [CrossRef] [Green Version]
- Bugnyar, T.; Massen, J.J.M. Avian Cognition; Healy, S., ten Cate, C.J., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 314–336. [Google Scholar]
- Fraser, O.N.; Bugnyar, T. Do ravens show consolation? Responses to distressed others. PLoS ONE 2010, 5, e10605. [Google Scholar] [CrossRef] [PubMed]
- De Waal, F.B.M. Putting the altruism back into altruism: The evolution of empathy. Annu. Rev. Psychol. 2008, 59, 279–300. [Google Scholar] [CrossRef] [PubMed]
- Provine, R.R.; Hamernik, H.B.; Curchack, B.C. Yawning: Relation to sleeping and stretching in humans. Ethology 1987, 76, 152–160. [Google Scholar] [CrossRef]
- Gallup, A.C.; Miller, R.; Clark, A.B. Changes in ambient temperature trugger yawning but not stretching in rats. Ethology 2011, 117, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.L.; Gallup, A.C.; Vogel, A.R.; Clark, A.B. Auditory disturbances promote temporal clustering of yawning and stretching in small groups of budgerigars (Melopsittacus undulatus). J. Comp. Psychol. 2012, 126, 324. [Google Scholar] [CrossRef]
- Gallup, A.C.; Militello, J.; Swartwood, L.; Sackett, S. Experimental evidence of contagious stretching and ingroup bias in budgerigars (Melopsittacus undulatus). J. Comp. Psychol. 2017, 131, 69. [Google Scholar] [CrossRef]
- Schut, C.; Grossman, S.; Gieler, U.; Kupfer, J.; Yosipovitch, G. Contagious itch: What we know and what we would like to know. Front. Hum. Neurosci. 2015, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, K. Observing conspecifics scratching induces a contagion of scratching in Japanese monkeys (Macaca fuscata). J. Comp. Psychol. 2004, 118, 20. [Google Scholar] [CrossRef]
- Yu, Y.Q.; Barry, D.M.; Hao, Y.; Liu, X.T.; Chen, Z.F. Molecular and neural basis of contagious itch behavior in mice. Science 2017, 355, 1072–1076. [Google Scholar] [CrossRef] [Green Version]
- Helt, M.S.; Sorensen, T.M.; Scheub, R.J.; Nakhle, M.B.; Luddy, A.C. Patterns of contagious yawning and itching differ amongst adults with autistic traits vs. psychopathic traits. Front. Psychol. 2021, 12, 645310. [Google Scholar] [CrossRef]
- Haker, H.; Kawohl, W.; Herwig, U.; Rössler, W. Mirror neuron activity during contagious yawning—An fMRI study. Brain Imaging Behav. 2013, 7, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Holle, H.; Warne, K.; Seth, A.K.; Critchley, H.D.; Ward, J. Neural basis of contagious itch and why some people are more prone to it. Proc. Natl. Acad. Sci. USA 2012, 109, 19816–19821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munteanu, A.M.; Stocker, M.; Stöwe, M.; Massen, J.J.M.; Bugnyar, T. Behavioural and hormonal stress responses to social separation in ravens, Corvus corax. Ethology 2017, 123, 123–135. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: https://www.R-project.org/ (accessed on 25 April 2022).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Shoup-Knox, M.L.; Gallup, A.C.; Gallup, G.; McNay, E.C. Yawning and stretching predict brain temperature changes in rats: Support for the thermoregulatory hypothesis. Front. Evol. Neurosci. 2010, 2, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baenninger, R. Some comparative aspects of yawning in Betta splendens, Homo sapiens, Panthera leo, and Papio sphinx. J. Comp. Psychol. 1987, 101, 349. [Google Scholar] [CrossRef]
- Sokal, R.; Rohlf, F. Biometry: The Principles and Practice of Statistics in Bio-Logical Research, 2nd ed.; WH Freeman: New York, NY, USA, 1995. [Google Scholar]
- Wyndham, E. Diurnal cycle, behaviour and social organization of the budgerigar. Melopsittacus Undulatus. Emu 1980, 80, 25–33. [Google Scholar] [CrossRef]
- Marzluff, J.M.; Heinrich, B.; Marzluff, C.S. Raven roosts are mobile information centres. Anim. Behav. 1996, 51, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Loretto, M.C.; Schuster, R.; Itty, C.; Marchand, P.; Genero, F.; Bugnyar, T. Fission-fusion dynamics over large distances in raven non-breeders. Sci. Rep. 2017, 7, 1–9. [Google Scholar]
- Smuts, B.B.; Cheney, D.L.; Seyfarth, R.M.; Wrangham, R.W.; Struhsaker, T.T. Primate Societies; University of Chicago Press: Chicago, MI, USA, 1987. [Google Scholar]
- Gallup, A.C.; Kret, M.E.; Eldakar, O.T.; Folz, J.; Massen, J.J. People that score high on psychopathic traits are less likely to yawn contagiously. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Ikkatai, Y.; Okanoya, K.; Seki, Y. Observing real-time social interaction via telecommunication methods in budgerigars (Melopsittacus undulatus). Behav. Processes 2016, 128, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Gallup, A.C.; Miller, M.L.; Clark, A.B. Yawning and thermoregulation in budgerigars, Melopsittacus undulatus. Anim. Behav. 2009, 77, 109–113. [Google Scholar] [CrossRef]
- Gallup, A.C.; Miller, M.L.; Clark, A.B. The direction and range of ambient temperature change influences yawning in budgerigars (Melopsittacus undulatus). J. Comp. Psychol. 2010, 124, 133–138. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yawning: a wide opening of the beak combined with a slight closing of the eye. |
| Stretching: a lifting of one or both wings with extension above the legs and towards the anterior, or a lifting of both wings upwards and backwards. |
| Scratching: a raising of a leg towards the side and above the wing to brush against the body. |
| Shaking: a shuddering of the feathers followed by a brief pause in which the feathers are redirected towards their natural positioning. |
| Individual (Sex) | Yawns | Stretches | Scratches | Shakes |
|---|---|---|---|---|
| Group 1 | ||||
| Adele (F) | 0 | 29 | 26 | 23 |
| Paul (M) | 4 | 20 | 34 | 45 |
| Rufus (M) | 8 | 22 | 19 | 74 |
| Max (M) | 8 | 19 | 8 | 16 |
| Group 2 | ||||
| George (M) | 0 | 2 | 20 | 15 |
| Horst (M) | 0 | 6 | 11 | 17 |
| Laggie (M) | 0 | 18 | 13 | 23 |
| Louise (F) | 0 | 8 | 1 | 22 |
| Nobel (F) | 3 | 11 | 13 | 25 |
| Tom (M) | 0 | 22 | 19 | 38 |
| Mean ± SD | 2.30 ± 3.335 | 15.70 ± 7.55 | 17.70 ± 8.06 | 29.80 ± 18.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallup, A.C.; Schild, A.B.; Ühlein, M.A.; Bugnyar, T.; Massen, J.J.M. No Evidence for Contagious Yawning in Juvenile Ravens (Corvus corax): An Observational Study. Animals 2022, 12, 1357. https://doi.org/10.3390/ani12111357
Gallup AC, Schild AB, Ühlein MA, Bugnyar T, Massen JJM. No Evidence for Contagious Yawning in Juvenile Ravens (Corvus corax): An Observational Study. Animals. 2022; 12(11):1357. https://doi.org/10.3390/ani12111357
Chicago/Turabian StyleGallup, Andrew C., Anja B. Schild, Markus A. Ühlein, Thomas Bugnyar, and Jorg J. M. Massen. 2022. "No Evidence for Contagious Yawning in Juvenile Ravens (Corvus corax): An Observational Study" Animals 12, no. 11: 1357. https://doi.org/10.3390/ani12111357
APA StyleGallup, A. C., Schild, A. B., Ühlein, M. A., Bugnyar, T., & Massen, J. J. M. (2022). No Evidence for Contagious Yawning in Juvenile Ravens (Corvus corax): An Observational Study. Animals, 12(11), 1357. https://doi.org/10.3390/ani12111357