
Towards Ecological Management and Sustainable Urban Planning in Seoul, South Korea: Mapping Wild Pollinator Habitat Preferences and Corridors Using Citizen Science Data

 , ,
, ,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
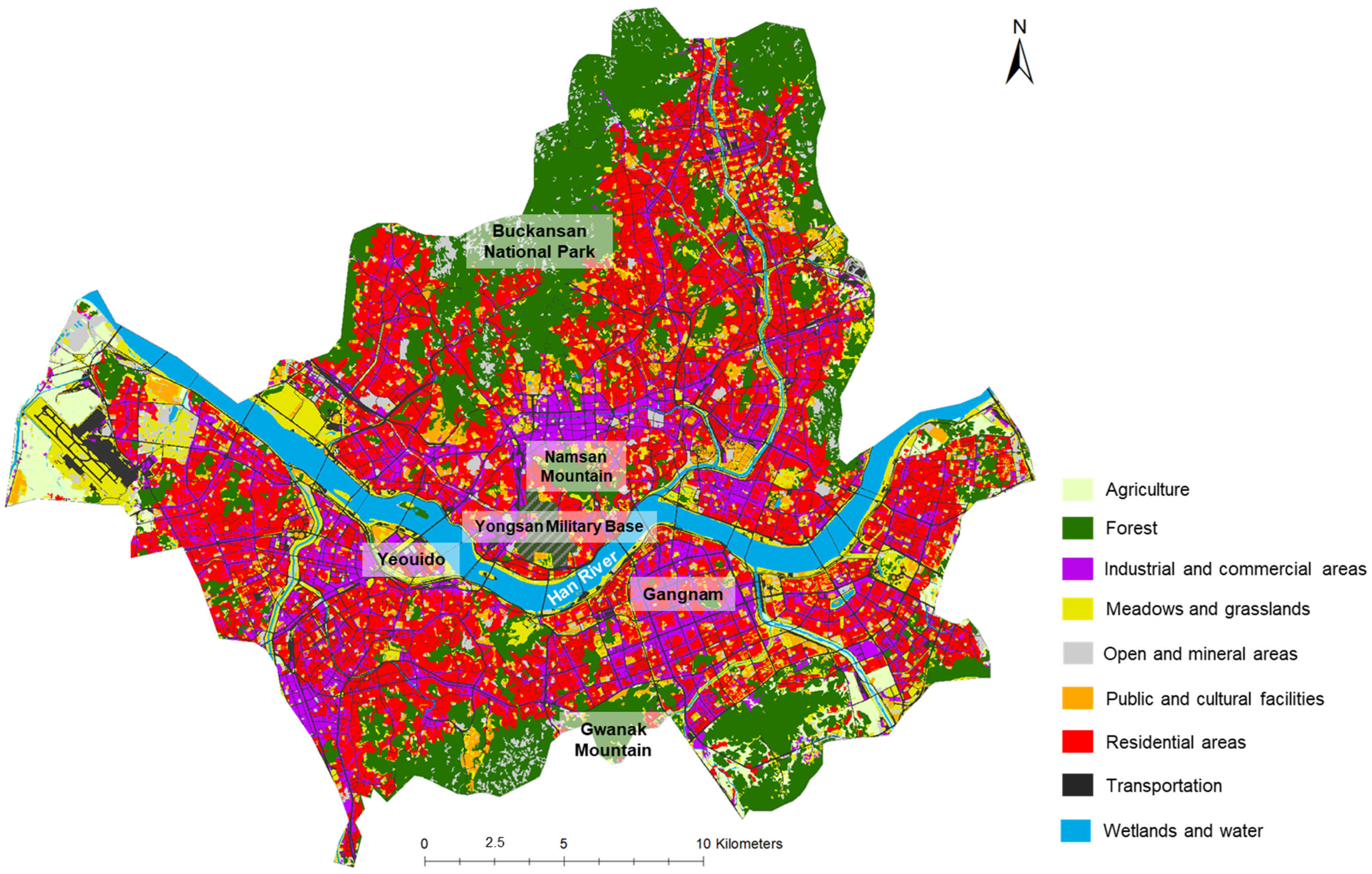
2.1. Study Area
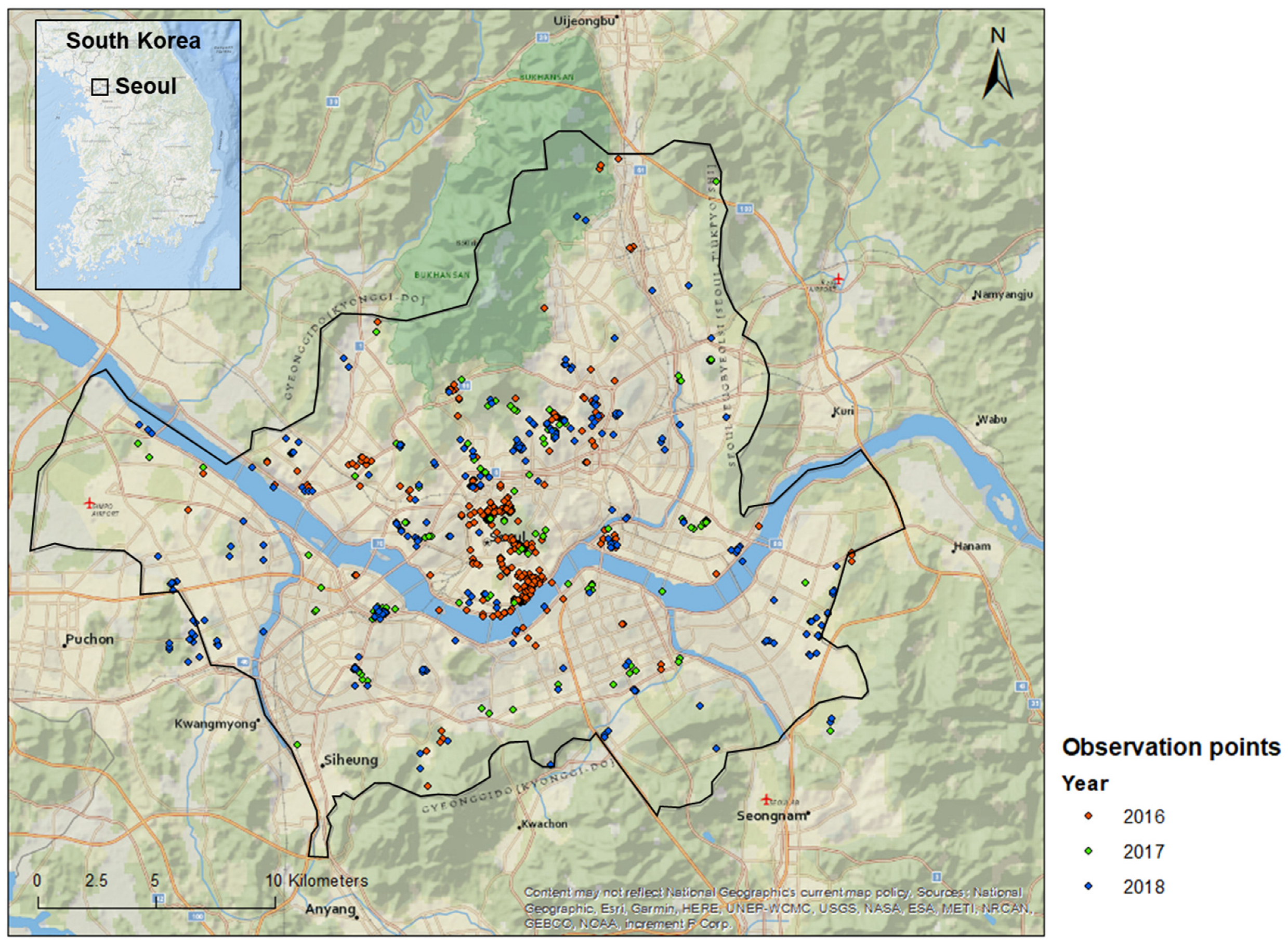
2.2. Data Collection
2.3. Estimating Pollinator Richness
2.4. Land-Use Affinity of Major Pollinator Families
2.5. Habitat Suitability Modeling
2.5.1. Variables
2.5.2. Maximum Entropy
2.5.3. Classification
2.6. Multi-Family Habitats and Corridors
2.7. Map Synthesis for Sustainable Urban Planning and Conservation Strategies
2.8. Corridor Prioritization for Action
3. Results
3.1. Community Composition
3.2. Richness and Land-Use Affinity by Family
3.3. Suitable Habitat by Family
3.4. Multi-Family Habitats and Corridors
3.5. Prioritizing Habitats and Corridors for Conservation
4. Discussion
4.1. Enhancing Scientific Knowledge about Pollinators in Seoul with Citizen Science
4.2. Improving Green Infrastructure for Pollinators in Seoul
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. World Urbanization Prospects: The 2018 Revision; United Nations: New York, NY, USA, 2018. [Google Scholar]
- McKinney, M.L. Urbanization as a Major Cause of Biotic Homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of Urbanization on Species Richness: A Review of Plants and Animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Kong, F.; Yin, H.; Nakagoshi, N.; Zong, Y. Urban Green Space Network Development for Biodiversity Conservation: Identification Based on Graph Theory and Gravity Modeling. Landsc. Urban Plan. 2010, 95, 16–27. [Google Scholar] [CrossRef]
- Oliveira, S.; Andrade, H.; Vaz, T. The Cooling Effect of Green Spaces as a Contribution to the Mitigation of Urban Heat: A Case Study in Lisbon. Build. Environ. 2011, 46, 2186–2194. [Google Scholar] [CrossRef]
- Liu, W.; Chen, W.; Peng, C. Assessing the Effectiveness of Green Infrastructures on Urban Flooding Reduction: A Community Scale Study. Ecol. Model. 2014, 291, 6–14. [Google Scholar] [CrossRef]
- Richardson, E.A.; Pearce, J.; Mitchell, R.; Kingham, S. Role of Physical Activity in the Relationship between Urban Green Space and Health. Public Health 2013, 127, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Tzoulas, K.; Korpela, K.; Venn, S.; Yli-Pelkonen, V.; Kaźmierczak, A.; Niemela, J.; James, P. Promoting Ecosystem and Human Health in Urban Areas Using Green Infrastructure: A Literature Review. Landsc. Urban Plan. 2007, 81, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, S. The Restorative Benefits of Nature: Toward an Integrative Framework. J. Environ. Psychol. 1995, 15, 169–182. [Google Scholar] [CrossRef]
- Korpela, K.; Borodulin, K.; Neuvonen, M.; Paronen, O.; Tyrväinen, L. Analyzing the Mediators between Nature-Based Outdoor Recreation and Emotional Well-Being. J. Environ. Psychol. 2014, 37, 1–7. [Google Scholar] [CrossRef]
- Nutsford, D.; Pearson, A.L.; Kingham, S. An Ecological Study Investigating the Association between Access to Urban Green Space and Mental Health. Public Health 2013, 127, 1005–1011. [Google Scholar] [CrossRef]
- Van den Berg, M.; van Poppel, M.; van Kamp, I.; Andrusaityte, S.; Balseviciene, B.; Cirach, M.; Danileviciute, A.; Ellis, N.; Hurst, G.; Masterson, D.; et al. Visiting Green Space Is Associated with Mental Health and Vitality: A Cross-Sectional Study in Four European Cities. Health Place 2016, 38, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Cho, J. Urban Planning and Urban Sprawl in Korea. Urban Policy Res. 2005, 23, 203–218. [Google Scholar] [CrossRef]
- Lee, J.Y.; Anderson, C.D. The Restored Cheonggyecheon and the Quality of Life in Seoul. J. Urban Technol. 2013, 20, 3–22. [Google Scholar] [CrossRef]
- Kang, T.-H.; Yoo, S.-H.; Kim, I.-K.; Cho, H.-J.; Shin, Y.-U. Change of Avifauna Following Restoration and Management in Cheonggye Stream, Seoul, Korea. J. Korean Nat. 2012, 5, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Kevan, P.G. Pollinators as Bioindicators of the State of the Environment: Species, Activity and Diversity. Agric. Ecosyst. Environ. 1999, 74, 373–393. [Google Scholar] [CrossRef]
- Rollin, O.; Benelli, G.; Benvenuti, S.; Decourtye, A.; Wratten, S.D.; Canale, A.; Desneux, N. Weed-Insect Pollinator Networks as Bio-Indicators of Ecological Sustainability in Agriculture. A Review. Agron. Sustain. Dev. 2016, 36, 8. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, C.; Dajoz, I.; Meriguet, J.; Loreau, M. Functional Diversity of Plant–Pollinator Interaction Webs Enhances the Persistence of Plant Communities. PLoS Biol. 2005, 4, e1. [Google Scholar] [CrossRef]
- Potter, A.; LeBuhn, G. Pollination Service to Urban Agriculture in San Francisco, CA. Urban Ecosyst. 2015, 18, 885–893. [Google Scholar] [CrossRef]
- Hall, D.M.; Camilo, G.R.; Tonietto, R.K.; Ollerton, J.; Ahrné, K.; Arduser, M.; Ascher, J.S.; Baldock, K.C.R.; Fowler, R.; Frankie, G.; et al. The city as a refuge for insect pollinators. Conserv. Biol. 2017, 31, 24–29. [Google Scholar] [CrossRef]
- McFrederick, Q.S.; LeBuhn, G. Are Urban Parks Refuges for Bumble Bees Bombus spp. (Hymenoptera: Apidae)? Biol. Conserv. 2006, 129, 372–382. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Aronson, M.F.J.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S. Biodiversity in the City: Fundamental Questions for Understanding the Ecology of Urban Green Spaces for Biodiversity Conservation. BioScience 2017, 67, 799–807. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, J.L.; Zuckerberg, B.; Bonter, D.N. Citizen Science as an Ecological Research Tool: Challenges and Benefits. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 149–172. [Google Scholar] [CrossRef] [Green Version]
- Deguines, N.; Julliard, R.; De Flores, M.; Fontaine, C. The Whereabouts of Flower Visitors: Contrasting Land-Use Preferences Revealed by a Country-Wide Survey Based on Citizen Science. PLoS ONE 2012, 7, e45822. [Google Scholar] [CrossRef] [PubMed]
- Suzuki-Ohno, Y.; Yokoyama, J.; Nakashizuka, T.; Kawata, M. Utilization of Photographs Taken by Citizens for Estimating Bumblebee Distributions. Sci. Rep. 2017, 7, 11215. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Sumner, S.; Bevan, P.; Hart, A.G.; Isaac, N.J.B. Mapping Species Distributions in 2 Weeks Using Citizen Science. Insect Conserv. Divers. 2019, 12, 382–388. [Google Scholar] [CrossRef]
- Giraud, C.; Calenge, C.; Coron, C.; Julliard, R. Capitalizing on Opportunistic Data for Monitoring Relative Abundances of Species. Biometrics 2016, 72, 649–658. [Google Scholar] [CrossRef]
- Horns, J.J.; Adler, F.R.; Şekercioğlu, Ç.H. Using Opportunistic Citizen Science Data to Estimate Avian Population Trends. Biol. Conserv. 2018, 221, 151–159. [Google Scholar] [CrossRef]
- Pearce, J.L.; Boyce, M.S. Modelling Distribution and Abundance with Presence-Only Data. J. Appl. Ecol. 2006, 43, 405–412. [Google Scholar] [CrossRef]
- van Strien, A.J.; van Swaay, C.A.M.; Termaat, T. Opportunistic Citizen Science Data of Animal Species Produce Reliable Estimates of Distribution Trends If Analysed with Occupancy Models. J. Appl. Ecol. 2013, 50, 1450–1458. [Google Scholar] [CrossRef]
- Mair, L.; Harrison, P.J.; Jönsson, M.; Löbel, S.; Nordén, J.; Siitonen, J.; Lämås, T.; Lundström, A.; Snäll, T. Evaluating Citizen Science Data for Forecasting Species Responses to National Forest Management. Ecol. Evol. 2017, 7, 368–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serret, H.; Deguines, N.; Jang, Y.; Lois, G.; Julliard, R. Data Quality and Participant Engagement in Citizen Science: Comparing Two Approaches for Monitoring Pollinators in France and South Korea. Citizen Sci. Theory Pract. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Joo, Y.-M. Megacity Seoul: Urbanization and the Development of Modern South Korea; Routledge: New York, NY, USA, 2019. [Google Scholar]
- Palmer, M.W. The Estimation of Species Richness by Extrapolation. Ecology 1990, 71, 1195–1198. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; Stevens, M.H.H. Vegan: Community Ecology Package, Version 2.6-2; Free Software Foundation, Inc.: Boston, MA, USA, 2019. [Google Scholar]
- Wickham, H.; Chang, W.; Wickham, M.H. Package ‘Ggplot2.’ Create Elegant Data Visualisations Using the Grammar of Graphics, Version 2; RStudio: Boston, MA, USA, 2016; pp. 1–189. [Google Scholar]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting Species Distributions for Conservation Decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A Maximum Entropy Approach to Species Distribution Modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; Association for Computing Machinery: New York, NY, USA, 2004; p. 83. [Google Scholar]
- Bradsworth, N.; White, J.G.; Isaac, B.; Cooke, R. Species Distribution Models Derived from Citizen Science Data Predict the Fine Scale Movements of Owls in an Urbanizing Landscape. Biol. Conserv. 2017, 213, 27–35. [Google Scholar] [CrossRef]
- Buckman-Sewald, J.; Whorton, C.R.; Root, K.V. Developing Macrohabitat Models for Bats in Parks Using Maxent and Testing Them with Data Collected by Citizen Scientists. Int. J. Biodivers. Conserv. 2014, 6, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Ropars, L.; Dajoz, I.; Fontaine, C.; Muratet, A.; Geslin, B. Wild Pollinator Activity Negatively Related to Honey Bee Colony Densities in Urban Context. PLoS ONE 2019, 14, e0222316. [Google Scholar] [CrossRef] [Green Version]
- Jueterbock, A.; Smolina, I.; Coyer, J.A.; Hoarau, G. The Fate of the Arctic Seaweed Fucus Distichus under Climate Change: An Ecological Niche Modeling Approach. Ecol. Evol. 2016, 6, 1712–1724. [Google Scholar] [CrossRef] [Green Version]
- Bellamy, C.C.; van der Jagt, A.P.; Barbour, S.; Smith, M.; Moseley, D. A Spatial Framework for Targeting Urban Planning for Pollinators and People with Local Stakeholders: A Route to Healthy, Blossoming Communities? Environ. Res. 2017, 158, 255–268. [Google Scholar] [CrossRef]
- Foltête, J.-C.; Clauzel, C.; Vuidel, G. A Software Tool Dedicated to the Modelling of Landscape Networks. Environ. Model. Softw. 2012, 38, 316–327. [Google Scholar] [CrossRef]
- Galpern, P.; Manseau, M.; Fall, A. Patch-Based Graphs of Landscape Connectivity: A Guide to Construction, Analysis and Application for Conservation. Biol. Conserv. 2011, 144, 44–55. [Google Scholar] [CrossRef]
- Saura, S.; Pascual-Hortal, L. A New Habitat Availability Index to Integrate Connectivity in Landscape Conservation Planning: Comparison with Existing Indices and Application to a Case Study. Landsc. Urban Plan. 2007, 83, 91–103. [Google Scholar] [CrossRef]
- Harmon-Threatt, A.N.; Hendrix, S.D. Prairie Restorations and Bees: The Potential Ability of Seed Mixes to Foster Native Bee Communities. Basic Appl. Ecol. 2015, 16, 64–72. [Google Scholar] [CrossRef]
- Tonietto, R.K.; Larkin, D.J. Habitat Restoration Benefits Wild Bees: A Meta-Analysis. J. Appl. Ecol. 2018, 55, 582–590. [Google Scholar] [CrossRef]
- Choi, S.-W.; Jung, C. Diversity of Insect Pollinators in Different Agricultural Crops and Wild Flowering Plants in Korea: Literature Review. J. Apic. 2015, 30, 191–201. [Google Scholar] [CrossRef]
- Bird, T.J.; Bates, A.E.; Lefcheck, J.S.; Hill, N.A.; Thomson, R.J.; Edgar, G.J.; Stuart-Smith, R.D.; Wotherspoon, S.; Krkosek, M.; Stuart-Smith, J.F.; et al. Statistical Solutions for Error and Bias in Global Citizen Science Datasets. Biol. Conserv. 2014, 173, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Baldock, K.C.R.; Goddard, M.A.; Hicks, D.M.; Kunin, W.E.; Mitschunas, N.; Morse, H.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; et al. A Systems Approach Reveals Urban Pollinator Hotspots and Conservation Opportunities. Nat. Ecol. Evol. 2019, 3, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Prévot, A.-C.; Cheval, H.; Raymond, R.; Cosquer, A. Routine Experiences of Nature in Cities Can Increase Personal Commitment toward Biodiversity Conservation. Biol. Conserv. 2018, 226, 1–8. [Google Scholar] [CrossRef]
- Ahn, S.-Y.; Baek, H.-J. Academic Achievement-Oriented Society and Its Relationship to the Psychological Well-Being of Korean Adolescents. In The Psychological Well-Being of East Asian Youth; Yi, C.-C., Ed.; Quality of Life in Asia; Springer: Dordrecht, The Netherlands, 2013; pp. 265–279. ISBN 978-94-007-4081-5. [Google Scholar]
- Juon, H.-S.; Nam, J.J.; Ensminger, M.E. Epidemiology of Suicidal Behavior among Korean Adolescents. J. Child Psychol. Psychiatry 1994, 35, 663–676. [Google Scholar] [CrossRef]
- Park, J.-Y.; Chung, I.-J. Adolescent Suicide Triggered by Problems at School in Korea: Analyses Focusing on Depression, Suicidal Ideation, Plan, and Attempts as Four Dimensions of Suicide. Child Indic. Res. 2014, 7, 75–88. [Google Scholar] [CrossRef]
- Collins, R.; Schaafsma, M.; Hudson, M.D. The Value of Green Walls to Urban Biodiversity. Land Use Policy 2017, 64, 114–123. [Google Scholar] [CrossRef]
- Madre, F.; Clergeau, P.; Machon, N.; Vergnes, A. Building Biodiversity: Vegetated Façades as Habitats for Spider and Beetle Assemblages. Glob. Ecol. Conserv. 2015, 3, 222–233. [Google Scholar] [CrossRef] [Green Version]
- MacIvor, J.S.; Ksiazek, K. Invertebrates on Green Roofs. In Green Roof Ecosystems; Sutton, R.K., Ed.; Ecological Studies; Springer International Publishing: Cham, Switzerland, 2015; pp. 333–355. ISBN 978-3-319-14983-7. [Google Scholar]
- Williams, N.S.G.; Lundholm, J.; Scott MacIvor, J. FORUM: Do Green Roofs Help Urban Biodiversity Conservation? J. Appl. Ecol. 2014, 51, 1643–1649. [Google Scholar] [CrossRef] [Green Version]
- Omar, M.; Al Sayed, N.; Barré, K.; Halwani, J.; Machon, N. Drivers of the Distribution of Spontaneous Plant Communities and Species within Urban Tree Bases. Urban For. Urban Green. 2018, 35, 174–191. [Google Scholar] [CrossRef]
- Säumel, I.; Weber, F.; Kowarik, I. Toward Livable and Healthy Urban Streets: Roadside Vegetation Provides Ecosystem Services Where People Live and Move. Environ. Sci. Policy 2016, 62, 24–33. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Variables | Betamultiplier | Threshold | TSS | AUC |
|---|---|---|---|---|---|
| Apidae | 13 | 2 | 0.4079 | 0.6047 ± 0.0936 | 0.8306 ± 0.0334 |
| Halictidae | 14 | 2 | 0.3635 | 0.6742 ± 0.1308 | 0.8706 ± 0.0330 |
| Lycaenidae | 53 | 2 | 0.4605 | 0.5860 ± 0.1181 | 0.9057 ± 0.0547 |
| Megachilidae | 9 | 3 | 0.4785 | 0.5302 ± 0.1867 | 0.8624 ± 0.0563 |
| Pieridae | 11 | 2 | 0.4529 | 0.5496 ± 0.0816 | 0.8276 ± 0.0411 |
| Syrphidae | 15 | 2 | 0.4276 | 0.5726 ± 0.0554 | 0.8272 ± 0.0324 |
| Classification | Total Occurrences | Percentage of Observations |
|---|---|---|
| Hymenoptera | 1061 | 57.6% |
| Apoidae | 901 | 48.9% |
| Apidae | 684 | 37.1% |
| Halictidae | 147 | 8.0% |
| Megachilidae | 37 | 2.0% |
| Andrenidae | 10 | 0.5% |
| Crabronidae | 6 | 0.3% |
| Colletidae | 2 | 0.1% |
| Other or not identifiable at the family scale | 15 | 0.8% |
| Formicoidae | 54 | 2.9% |
| Vespoidae | 37 | 2.0% |
| Tenthredinoidae | 8 | 0.4% |
| Chrysidoidea | 2 | 0.1% |
| Ichneumonoidae | 2 | 0.1% |
| Other/not identifiable at the super-family scale | 57 | 3.1% |
| Diptera | 326 | 17.7% |
| Syrphidae | 225 | 12.2% |
| Muscomorpha | 30 | 1.6% |
| Sarcophagoidae | 17 | 0.9% |
| Other or not identifiable at the super-family scale | 54 | 2.9% |
| Lepidoptera | 288 | 15.6% |
| Papilionoidae | 248 | 13.5% |
| Lycaenidae | 74 | 4.0% |
| Nymphalidae | 22 | 1.2% |
| Papilionidae | 10 | 0.5% |
| Pieridae | 142 | 7.7% |
| Hesperioidea | 13 | 0.7% |
| Bombycoidae | 5 | 0.3% |
| Zygaenidae | 2 | 0.1% |
| Other or not identifiable at the super-family scale | 18 | 1.0% |
| Hemiptera | 105 | 5.7% |
| Lygaeoidea | 48 | 2.6% |
| Coreoidae | 18 | 1.0% |
| Pentatomoidae | 13 | 0.7% |
| Other or not identifiable at the super-family scale | 26 | 1.4% |
| Coleoptera | 49 | 2.7% |
| Scarabaeoidae | 10 | 0.5% |
| Chrysomeloidae | 8 | 0.4% |
| Cucujoidae | 7 | 0.4% |
| Curculionoidae | 5 | 0.3% |
| Other or not identifiable at the super-family scale | 19 | 1.0% |
| Othoptera | 6 | 0.3% |
| Arachnida | 6 | 0.3% |
| Thomisidae | 5 | 0.3% |
| Other or not identifiable at the family scale | 1 | 0.1% |
| Odonata | 1 | 0.1% |
| Land Class | Observed Richness | Estimated Jackknife | Estimated Bootstrap |
|---|---|---|---|
| Agriculture | 53 | 83.7 ± 5.5 | 66.2 ± 2.6 |
| Forest | 44 | 75.7 ± 5.6 | 55.9 ± 2.5 |
| Industry & Commercial | 29 | 47.7 ± 4.3 | 36.7 ± 1.2 |
| Meadows and grasslands | 61 | 92.8 ± 5.6 | 74.5 ± 2.8 |
| Open and mineral areas | 30 | 47.7 ± 4.2 | 37.1 ± 1.9 |
| Public and cultural facilities | 37 | 61.7 ± 4.9 | 46.9 ± 2.3 |
| Residential areas | 52 | 79.9 ± 5.3 | 63.3 ± 2.6 |
| Transportation | 34 | 51.9 ± 4.2 | 41.1 ± 2.1 |
| Wetlands and water | 9 | 14.7 ± 2.3 | 11.2 ± 1.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serret, H.; Andersen, D.; Deguines, N.; Clauzel, C.; Park, W.-H.; Jang, Y. Towards Ecological Management and Sustainable Urban Planning in Seoul, South Korea: Mapping Wild Pollinator Habitat Preferences and Corridors Using Citizen Science Data. Animals 2022, 12, 1469. https://doi.org/10.3390/ani12111469
Serret H, Andersen D, Deguines N, Clauzel C, Park W-H, Jang Y. Towards Ecological Management and Sustainable Urban Planning in Seoul, South Korea: Mapping Wild Pollinator Habitat Preferences and Corridors Using Citizen Science Data. Animals. 2022; 12(11):1469. https://doi.org/10.3390/ani12111469
Chicago/Turabian StyleSerret, Hortense, Desiree Andersen, Nicolas Deguines, Céline Clauzel, Wan-Hyeok Park, and Yikweon Jang. 2022. "Towards Ecological Management and Sustainable Urban Planning in Seoul, South Korea: Mapping Wild Pollinator Habitat Preferences and Corridors Using Citizen Science Data" Animals 12, no. 11: 1469. https://doi.org/10.3390/ani12111469
APA StyleSerret, H., Andersen, D., Deguines, N., Clauzel, C., Park, W.-H., & Jang, Y. (2022). Towards Ecological Management and Sustainable Urban Planning in Seoul, South Korea: Mapping Wild Pollinator Habitat Preferences and Corridors Using Citizen Science Data. Animals, 12(11), 1469. https://doi.org/10.3390/ani12111469