1. Introduction
LPS, the major structural ingredient of gram-negative bacteria’s cell wall, can initiate intense systemic inflammatory reaction evidenced as the production of inflammatory factors and the altered expressions of genes related to immune system and inflammation [
1,
2]. Accumulated evidence from previous studies have shown that LPS stimulation can result in comprised growth performance, enhanced production of inflammatory cytokines and dysfunction of intestine structure and function [
3,
4,
5]. Furthermore, in various pathological conditions such as sepsis, inflammation and trauma, it appears to be an increase of intestinal permeability [
6,
7]. It has been demonstrated that LPS challenge elevated D-lactic acid concentration (D-LA) and diamine oxidase (DAO) activity in serum of broilers administrated with LPS, which are the indicators of intestinal permeability [
8]. In addition, intestinal tight junction protein is an indicator of intestinal permeability [
9]. Intestinal epithelial cells play a critical role in preserving the wholeness of the epithelial barrier, including the intestinal cytomembrane and tight junctions between enterocytes [
10]. These intercellular connections form a barrier for the epithelial cells, which contribute to separating the extracellular fluid at the luminal side from fluid at the serosal side and preventing the interstitial tissues from microbial invasion [
11]. Moreover, the special structures making up cell-cell junctions, which contain tight junctions (TJs) and adherens junctions (AJs), affect the effectiveness and stability of the gastrointestinal epithelial barrier [
12]. It has been reported that more than 40 different tight junction proteins have been found, which include
occludin,
claudin-1, and zonula occludens-1 (
ZO-1) [
13]. However, in the condition of inflammatory or stress responses, the expression of intestinal tight proteins were downregulated. In previous studies, it was demonstrated that LPS challenge reduced the expression levels of tight junction proteins in broilers [
14] and in rats [
15]. In order to maintain the intestine health and growth performance of broilers at an early age, it is therefore necessary to apply nutritional interventions in broiler production to alleviate the adverse effects induced by LPS administration.
Glutamine (Gln), which serves as a preferential substrate for intestine epithelial cell proliferation and survival under inflammatory conditions, is considered to be conditionally essential for gut homeostasis and barrier function [
16,
17]. Additionally, it was demonstrated that Gln could be utilized at a high rate by rapid dividing cells such as immune cells and was necessary for lymphocyte proliferation and cytokine production [
18]. Although Gln has been considered to be the most abundant amino acid in the circulation, the physiological requirement for Gln may exceed the body’s synthesis capacity under some catabolic stresses [
19,
20]. Therefore, exogenous Gln addition maybe an effective method to alleviate immunological stress and improve intestine function in response to stressful conditions. However, to our knowledge, the mechanism of Gln regulating the intestinal epithelial barrier in LPS-challenged broilers remains unknown.
Therefore, the objectives of the present study were to investigate the effects of Gln supplementation on growth performance, inflammatory responses and intestinal mucosa barrier of broilers in immunological stress induced by LPS challenge, and furtherly demonstrate the underlying mechanism of Gln supplementation on inflammatory responses and intestinal mucosa barrier of LPS-challenged broilers.
4. Discussion
In previous studies, LPS challenge resulted in comprised growth performance of broilers [
26,
27]. In the present study, compared with saline treatment, we found that the growth performance of broilers injected with LPS was depressed, as shown by the reduction of ADG and ADFI and the increased F/G. In consistent with these, a previous study reported that LPS exposure resulted in the decrease of ADG and ADFI [
28]. which was mainly attributed to depressed appetite [
29], destroyed the intestinal barrier function [
3], restrained nutrient absorption [
30] and reallocation of nutrients [
5,
31]. In the condition of immune stress induced by LPS, diets nutrients were directed away from growth, but toward processes in support of various inflammatory immune response and synthesis of various mediators such as acute proteins and cytokines [
32]. As a result, the growth performance of broilers by LPS injection was depressed. However, the impaired effects of LPS challenge on ADG and ADFI of broilers were normalized by 1% Gln supplementation. Similar with the results of a previous study, in which it was reported that the growth performance of broilers was improved by 1% Gln supplementation, accompanied by the increased ADFI and ADG and the decreased F/G [
33]. Therefore, the results of mentioned above indicated that 1% Gln supplementation could contribute to ameliorating the adverse effects of immune stress induced by LPS challenge on the growth performance of broilers.
Intestinal morphology is one of behavioral markers to evaluate inflammation [
34]. In a previous study, in which the negative effects of LPS-challenge on intestine structure was confirmed shown as an increase of crypt depth and a decrease in villus height and the ratio of villus height to crypt depth [
28]. Similarly, the same tendency was also found in our study. However, in the present study, diets supplemented with Gln elevated villus height, increased crypt depth, coupled with the increased ratio of villus height to crypt depth in comparison with those administrated by LPS challenge. However, up to now, few studies have been found to study the effects of Gln addition on intestine morphology of broilers challenged by LPS. It has been demonstrated that an increase in villus height and the ration of villus height to crypt depth was observed in broilers under heat stress, which were supplemented with 1% Gln [
35]. Additionally, it was reported that Gln supplementation reduced crypt depth, increased villus height and the ration of villus height to crypt depth of broilers with necrotic enteritis challenge [
36]. Furthermore, it was demonstrated that improvement of intestinal morphology structure including longer intestinal villus height, deeper crypt depth and higher ratio of villus height to crypt depth contributed to alleviating the stress and ameliorating intestinal barrier functions [
37]. The results aforementioned showed that Gln may contribute to protecting intestine morphology and intestinal mucosal of broilers in the state of stress because Gln serves as a sufficient energy substrate for cell proliferation and differentiation [
35].
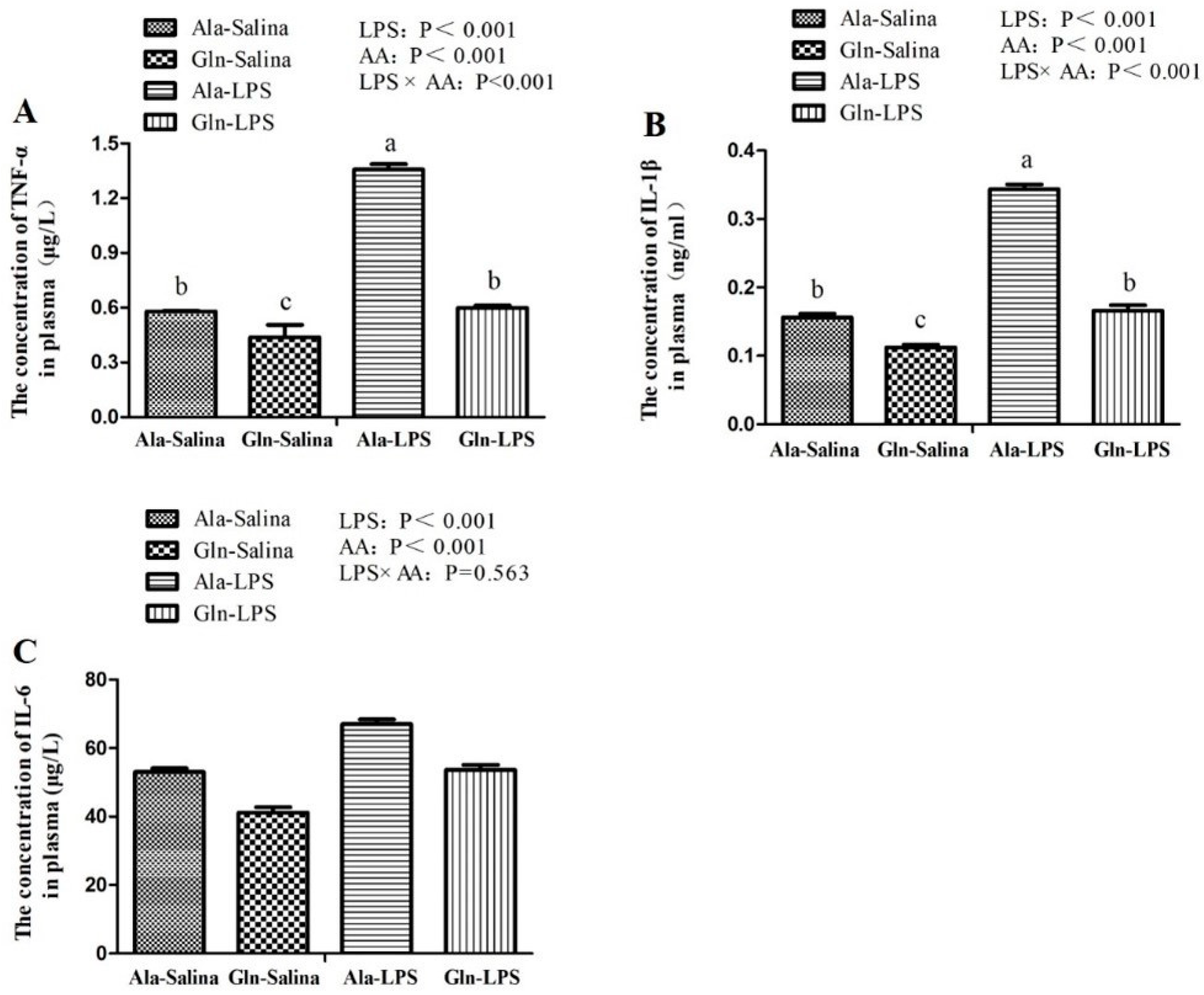
LPS is known to trigger the activity of immune system. Subsequently, the innate immune cells are activated, followed by the release of pro-inflammatory cytokines and anti-inflammatory cytokines [
5]. TNF-α, IL-6 and IL-1β, originated from macrophages, are the important cellular messengers that play a major role in diverse inflammatory responses and are highly bound up with the severity of inflammation [
38]. In our study, LPS challenge increased the concentration of TNF-α, IL-1β and IL-6 in plasma of broilers in comparison with those receiving saline injection. In consistent with the results of ours, the similar results were also observed by Wu et al. [
8] and Chen et al. [
39], who reported that the concentration of TNF-α, IL-1β and IL-6 of broilers were elevated in response to LPS challenge. In contrast, in the present study, Gln administration effectively depressed the increase in the contents of TNF-α, IL-1β and IL-6 in plasma induced by LPS challenge. Similarly, a study conducted by Soares et al. [
20] showed the similar results. The results of a previous study conducted by Xu et al. [
40] also showed that treatment with Gln significantly decreased the levels of TNF-α and IL-6. Moreover, it has been proved that Gln deficiency aggravates the production of proinflammatory cytokines, but Gln supplementation inhibits the inflammatory response in vitro [
41]. These therefore suggest that Gln may contribute to alleviating the negative effects of inflammatory responses in response to LPS challenge.
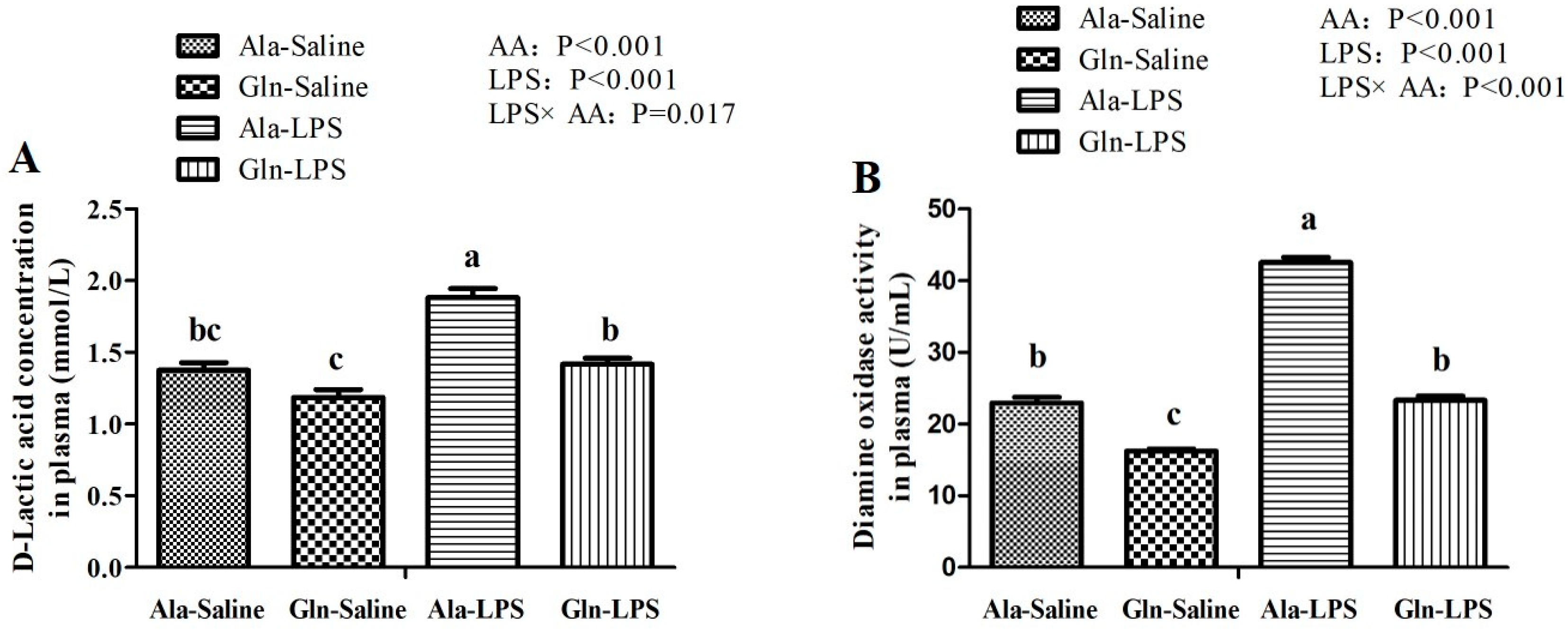
DAO, the class of copper-containing amine oxidases that catalyzes the deamination of histamines through oxidation. The levels of DAO is an indicator of intestinal mucosal barrier function and intestine permeability [
42]. DAO is localized in the intestinal epithelial cells and enters the blood circulation through the ruined intestinal barrier. D-LA, a product of fermentation produced by intestinal bacteria, is a sensitive biomarker to reflect intestine injury and to monitor intestinal permeability [
9]. In the present study, both DAO and D-LA levels in circulating were elevated by LPS administration, indicating that LPS challenge might seriously impaired intestinal barrier function and damage the intestinal structure, which was in consistent with the results of intestine morphology in the present study mentioned above. Interestingly, the deleterious effect of LPS was markedly reduced by administration of Gln. In agreement with a previous study conducted by Wu et al. [
35], who observed that Gln supplementation decreased the D-LA and DAO activity in the plasma of broilers. Indeed, Gln deprivation leads to the impaired paracellular permeability [
41]. These results indicated that the intestinal mucosa permeability of broilers supplemented with Gln had somewhat been improved.
The permeability of the intestinal mucosa is influenced by tight junction proteins [
42], Claudin family proteins belong to the transmembrane proteins family and are the most important tight junction protein, but ZO family proteins belong to peripheral membrane proteins and are important to tight junction assembly [
11]. In the condition of inflammatory responses such as trauma or sepsis, the expression of intestinal tight junction proteins were inhibited [
35]. Indeed, in the present study, the mRNA expression of ZO-1, claudin-1 and occludin in duodenum, jejunum and ileum were downregulated. In agreement with the results of ours, the similar results were also found by Gadde et al. [
6] and Li et al. [
3]. In contrast, Gln supplementation reversed the adverse effects of LPS on intestinal mucosa barriers. Recently, it has been established that Gln are involved in the regulation of gut tight junction proteins [
41]. Restoration of tight junctions has been reported after Gln supplementation both in vivo [
43] and in vitro [
44]. In addition, it was found that Gln deprivation in intestine epithelial cells was associated with a loss of tight junction proteins, whereas Gln addition rescued the phenotype of barrier dysfunction [
45]. The results mentioned above indicated that Gln may contribute to preserving the intestinal barrier function of LPS-challenged broilers and furtherly improving the intestinal permeability.
TLR4, which is a signaling receptor of LPS, is thought to be participated in the first immunologic barrier of digestive tract. In addition, TLR4 can initiate the activation of a complex signaling molecules containing various adaptor proteins, kinases and transcriptional factors. It has been proven that TLR4 can trigger the activation of the mitogen-activated protein kinase pathway through MyD88, which is a common adaptor molecule that is recruited towards the Toll/IL-1 receptor domain of the TLRs [
40]. FAK, an adaptor protein, plays a critical role in focal adhesion dynamics and initiating the TLR4 signal cascades. Moreover, MyD88 participated in the cytokine release induced by FAK-regulated protein I/II and the increase in the expression of IRAK4 [
46]. In our study, LPS stimulation dramatically increased the expression of TLR4, FAK, MyD88 and IRAK4 in duodenum, jejunum and ileum of LPS-challenged broilers compared with those receiving saline injection. In consistent with the results of ours, Guo et al. [
46] reported that LPS induces the activation of TLR4 signal transduction cascade, leading to the activation of FAK in intestinal epithelial cells followed by the activation of MyD88 and IRAK4, which contributes to inducing the open of intestinal tight junction. In addition, Luo et al. [
47] suggested that TLR4 signaling pathway mediated by LPS challenge maybe involved in the alteration of tight junction proteins via upregulating the transcription and translation of downstream inflammatory factors. These indicated that LPS could result in a quick increase in intestinal permeability and intestinal mucosal destruction by the TLR4/FAK/MyD88 signal transduction axis. To the best of our knowledge, the modulatory effect of Gln on LPS-challenged broilers was not available. Only a previous study about the regulation of Gln intestinal permeability and intestinal mucosa junction tight protein was focused on rats, in which Gln addition increased the expression of TLR4 and MyD88 associated with the morphological destruction of intestinal mucosa and the elevated expression of intestinal tight junction proteins [
40]. Furthermore, it has been shown that Gln supplementation may help to reduce the severity of infection, probably associated with the reduction of mucosal cytokine responses and the protection of intestinal barrier function [
48]. A study conducted used with piglets infected with Escherichia coli suggested that Gln addition tended to increase the expression of occudin in intestinal mucosa [
49]. Additionally, amino acids have important roles in the expression of tight junction proteins. Chen et al. [
39] suggested that L-threonine supplementation attenuated inflammatory responses and intestinal barrier damage induced by LPS injection. In the present study, Gln supplementation resulted in a greater effect on suppressing the expression of the related signaling molecules of TLR4/FAK/MyD88/IRAK4 pathway. Furthermore, an increase in the intestinal permeability of LPS-induced mouse was completely prevented in deficient mice of TLR4 or MyD88 or knockdown of FAK [
47], indicating that intestinal permeability and intestinal mucosa integrity were regulated by TLR4-dependent activation of FAK/MyD88/IRAK4 signal axis. Therefore, it was reasonable to speculate that Gln supplementation may be contribute to protecting the intestinal mucosa barrier and the prevention of intestinal barrier dysfunction of LPS-challenged broilers through TLR4/FAK/MyD88 signal transduction axis.


 ,
, 


{kind=link}
{kind=link}