
Cloning and Expression Analysis of HAT1 and HDAC1 in the Testes of Mature Yaks and Their Sterile Hybrids

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animals and Sampling
2.3. Sectioning and HE Staining of Yak and Cattle-Yak Epididymis
2.4. RNA Extraction and Reverse Transcription
2.5. Cloning and Sequencing of the HAT1 and HDAC1 Genes of Yaks
2.6. Analysis of HAT1 and HDAC1 mRNA Expressions by Quantitative PCR
2.7. Analysis of HAT1 and HDAC1 Protein Level by Western Blotting
2.8. Quantification of H3K9 Acetylation Level by ELISA
2.9. Statistical Analysis
3. Results
3.1. Histological Comparison of Yak and Cattle-Yak Epididymis
3.2. Cloning and Sequencing of Yak HAT1 and HDAC1 Genes
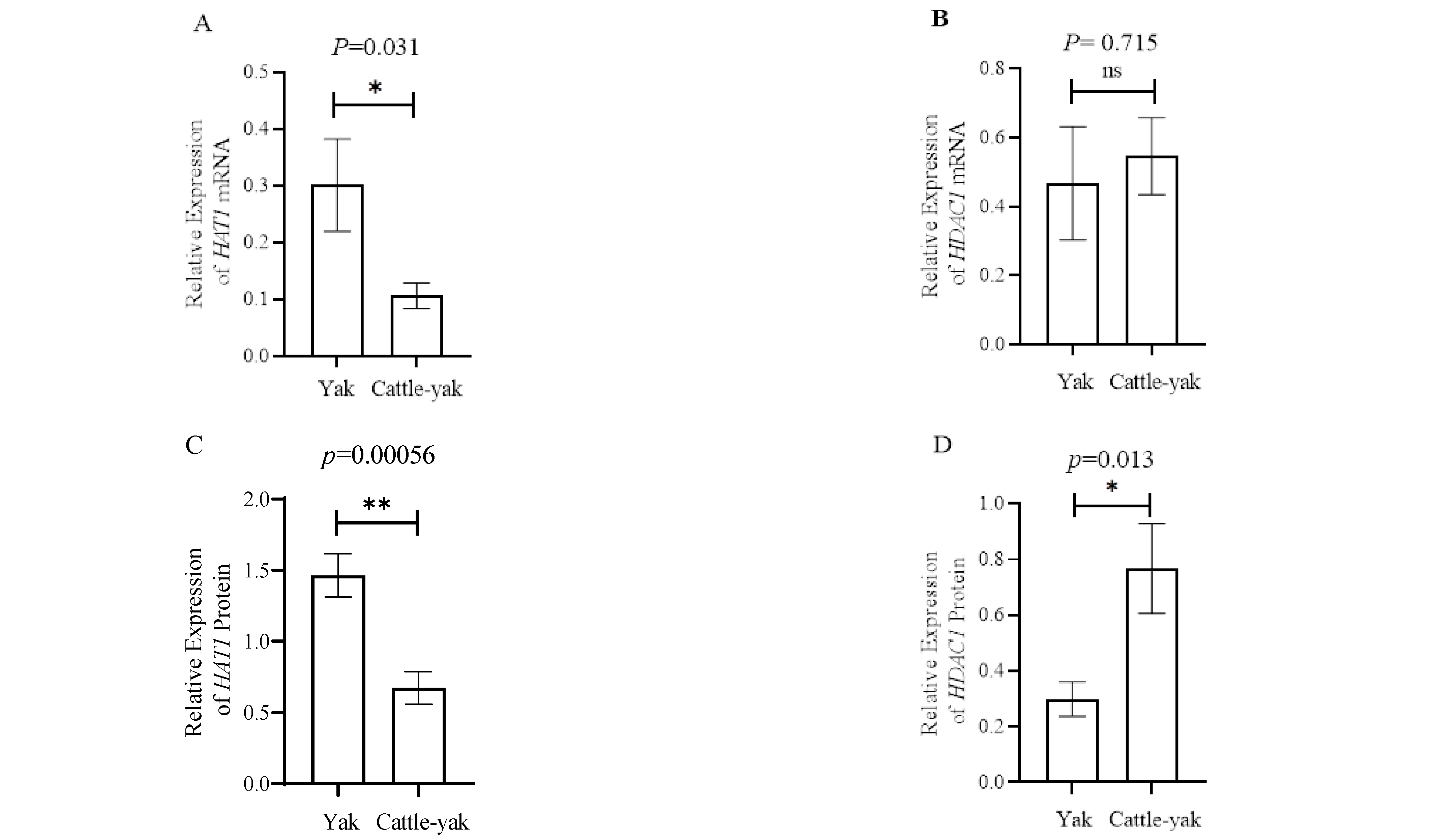
3.3. HAT1 and HDAC1 Expression in the Testes of Yaks and Cattle-Yaks
3.4. The Acetylation Level of H3K9 in the Testes of Yaks and Cattle-Yaks
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, Y.; Gautier, M.; Ding, X.; Zhang, H.; Wang, Y.; Wang, X.; Faruque, O.; Li, J.; Ye, S.; Gou, X.; et al. Species composition and environmental adaptation of indigenous Chinese cattle. Sci. Rep. 2017, 7, 16196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.N.; Liu, W.J.; Wang, C.L.; Huang, L.; Jin, S.Y.; Lin, Y.Q.; Zheng, Y.C. Histological evaluation and Prdm9 expression level in the testis of sterile male cattle-yaks. Livest. Sci. 2014, 160, 208–213. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, J.; Li, B. Research progress on meiotic gene expression and epigenetic regulation of male sterility in cattle. Genetics 2020, 42, 1081–1092. [Google Scholar]
- Tumennasan, K.; Tuya, T.; Hotta, Y.; Takase, H.; Speed, R.M.; Chandley, A.C. Fertility investigations in the F1 hybrid and backcross progeny of cattle (Bos taurus) andyak (B. grunniens) in Mongolia. Cytogenet. Genome Res. 1997, 78, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.-Y.; Li, Q.-F.; Qu, X.-G.; Li, Y.-X.; Li, X.-F.; Xu, H.-T. Expression levels of Cdc2 and Cdc25A mRNA in cattle, yak, and cattle-yak testis. Yi Chuan Hered. 2009, 31, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Li, B.J.; Wu, W.J.; Luo, H.; Liu, Z.; Liu, H.; Li, Q.; Pan, Z. Molecular characterization and epigenetic regulation of Mei1 in cattle and cattle–yak. Gene 2015, 573, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Jia, C.; Qu, X.G.; Zhao, X.B.; Zhong, J.C.; Xie, Z.; Li, Q.F. Cloning of b-Sycp2 gene of cattle, yak and cattle-yak and the expression level of testicular tissue mRNA. Sci. Agric. Sin. 2013, 46, 367–375. [Google Scholar]
- Wang, S.; Pan, Z.; Zhang, Q.; Xie, Z.; Liu, H.; Li, Q. Differential mRNA Expression and Promoter Methylation Status of SYCP3 Gene in Testes of Yaks and Cattle-Yaks. Reprod. Domest. Anim. 2012, 47, 455–462. [Google Scholar] [CrossRef]
- Rajender, S.; Avery, K.; Agarwal, A. Epigenetics, spermatogenesis and male infertility. Mutat. Res. Mutat. Res. 2011, 727, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Luense, L.J.; Wang, X.; Schon, S.; Weller, A.H.; Shiao, E.L.; Bryant, J.; Bartolomei, M.; Coutifaris, C.; Garcia, B.A.; Berger, S.L. Comprehensive analysis of histone post-translational modifications in mouse and human male germ cells. Epigenet. Chromatin 2016, 9, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Garcia, B.A. Comprehensive Catalog of Currently Documented Histone Modifications. Cold Spring Harb. Perspect. Biol. 2015, 7, a025064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouzarides, T. Histone acetylases and deacetylases in cell proliferation. Curr. Opin. Genet. Dev. 1999, 9, 40–48. [Google Scholar] [CrossRef]
- Sadoul, K.; Boyault, C.; Pabion, M.; Khochbin, S. Regulation of protein turnover by acetyltransferases and deacetylases. Biochimie 2008, 90, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Sabari, B.R.; Zhang, D.; Allis, C.D.; Zhao, Y. Metabolic regulation of gene expression through histone acylations. Nat. Rev. Mol. Cell Biol. 2017, 18, 90–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterner, D.E.; Berger, S.L. Acetylation of Histones and Transcription-Related Factors. Microbiol. Mol. Biol. Rev. 2000, 64, 435–459. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Endo, D.; Akiyama, N.; Yamamoto-Fukuda, T.; Koji, T. Aberrant levels of histone H3 acetylation induce spermatid anomaly in mouse testis. Histochem. Cell Biol. 2015, 143, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Liu, J.; An, S.; Nishino, T.; Hishikawa, Y.; Koji, T. Immunohistochemical Analysis of Histone H3 Modifications in Germ Cells during Mouse Spermatogenesis. Acta Histochem. Cytochem. 2011, 44, 183–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Zhang, C.; Diao, H. Histone acetylation and mammalian reproduction. J. Chin. J. Cell Biol. 2017, 39, 523–528. [Google Scholar]
- Kenneth, J.L.; Thomas, D.S. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2002, 25, 402–408. [Google Scholar]
- Shen, Z.; Huang, L.; Jin, S.; Zheng, Y. Cloning and Expression Analysis of Two Kdm Lysine Demethylases in the Testes of Mature Yaks and Their Sterile Hybrids. Animals 2020, 10, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Cui, Y. Protein acetylation and sperm DNA stability and sperm motility. J. Int. Reprod. Health Fam. 2016, 35, 118–122. [Google Scholar]
- Sun, L.; Mipam, T.D.; Zhao, F.; Liu, W.; Zhao, W.; Wu, S.; Xu, C.; Yu, S.; Cai, X. Comparative testis proteome of cattleyak from different developmental stages. Animal 2017, 11, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, P.A.A.; Hoover, M.E.; Zhang, P.; Nagarajan, P.; Freitas, M.A.; Parthun, M.R. Identification of multiple roles for histone acetyltransferase 1 in replication-coupled chromatin assembly. Nucleic Acids Res. 2017, 45, 9319–9335. [Google Scholar] [CrossRef] [Green Version]
- Sirbu, B.M.; Couch, F.B.; Feigerle, J.T.; Bhaskara, S.; Hiebert, S.W.; Cortez, D. Analysis of protein dynamics at active, stalled, and collapsed replication forks. Genes Dev. 2011, 25, 1320–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, J.J.; Geller, B.; Lipchik, A.M.; Chen, J.; Salahudeen, A.A.; Ram, A.N.; Ford, J.M.; Kuo, C.J.; Snyder, M.P. HAT1 Coordinates Histone Production and Acetylation via H4 Promoter Binding. Mol. Cell 2019, 75, 711–724.e5. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.M.; Cai, X.; Sun, L.; Yu, S.; Cai, X.; Sun, L.; Zuo, Z.; Mipam, T.D.; Cao, S.; Shen, L.; et al. Comparative iTRAQ proteomics revealed proteins associated with spermatogenic arrest of cattle-yak. J. Proteom. 2016, 142, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Senese, S.; Zaragoza, K.; Minardi, S.; Muradore, I.; Ronzoni, S.; Passafaro, A.; Bernard, L.; Draetta, G.F.; Alcalay, M.; Seiser, C.; et al. Role for Histone Deacetylase 1 in Human Tumor Cell Proliferation. Mol. Cell. Biol. 2007, 27, 4784–4795. [Google Scholar] [CrossRef] [Green Version]
- Peng, M.; Li, Y.; Huang, H.; Jin, F. The expression of GCN5, HDAC1 and DNMT1 in parthenogenetically activated mouse embryos. J. Obstet. Gynaecol. 2015, 35, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Dovey, O.M.; Foster, C.T.; Eisenman, C. Histone deacetylase 1 (hdac1), but not hdac2, controls embryonic stem cell differentiation. Proc. Natl. Acad. Sci. USA 2010, 107, 8242–8247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steilmann, C.; Paradowska, A.; Bartkuhn, M.; Vieweg, M.; Schuppe, H.-C.; Bergmann, M.; Kliesch, S.; Weidner, W.; Steger, K. Presence of histone H3 acetylated at lysine 9 in male germ cells and its distribution pattern in the genome of human spermatozoa. Reprod. Fertil. Dev. 2011, 23, 997–1011. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Accession No. | Gene Name | Primer Sequence (5´–3´) | Annealing Temperature (°C) | Product Size (bp) |
|---|---|---|---|---|
| NM_001034347.1 | HAT1 | F: TCGGAAATGGCGGGTTTGA R: AAGGGAAGTAATTGCAGTGGTA | 60 | 1559 |
| NM_001037444.2 | HDAC1 | F: GGACCGATTGACGGGAGGG R: GGGTTCAAGAGTTTGGGAGGG | 60 | 1699 |
| NM_001034347.1 | HAT1 | F: AGCCTATCAACAATGTTCCGTG R: AGCTTCTTTTTCCAGCAACG | 60 | 159 |
| NM_001037444.2 | HDAC1 | F: CCAGTGCAGTTGTCTTGCAG R: ACGAATGGTGTAGCCACCT | 60 | 158 |
| NR 036642 | 18S rRNA | F: CTGAGAAACGGCTACCACATC R: CAGACTTGCCCTCCAATGG | 59 | 168 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, S.; Shen, Z.; Jin, S.; Huang, L.; Zheng, Y. Cloning and Expression Analysis of HAT1 and HDAC1 in the Testes of Mature Yaks and Their Sterile Hybrids. Animals 2022, 12, 2018. https://doi.org/10.3390/ani12162018
Sun S, Shen Z, Jin S, Huang L, Zheng Y. Cloning and Expression Analysis of HAT1 and HDAC1 in the Testes of Mature Yaks and Their Sterile Hybrids. Animals. 2022; 12(16):2018. https://doi.org/10.3390/ani12162018
Chicago/Turabian StyleSun, Shijie, Zhenhua Shen, Suyu Jin, Lin Huang, and Yucai Zheng. 2022. "Cloning and Expression Analysis of HAT1 and HDAC1 in the Testes of Mature Yaks and Their Sterile Hybrids" Animals 12, no. 16: 2018. https://doi.org/10.3390/ani12162018