
Influence of Lonicera japonica and Radix Puerariae Crude Extracts on the Fecal Microbiome and Nutrient Apparent Digestibility of Finishing Pigs

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Animal Management
2.3. Experimental Diets
2.4. Analysis of Bacterial Community and Data Analysis
2.4.1. DNA Extraction and 16S rRNA Sequencing
2.4.2. Microbiota Data Analysis
2.5. Determination of Apparent Digestibility
2.6. Statistical Analysis
3. Results
3.1. Influences of L. japonica Crude Extract and Radix Puerariae Crude Extract on Growth Performance
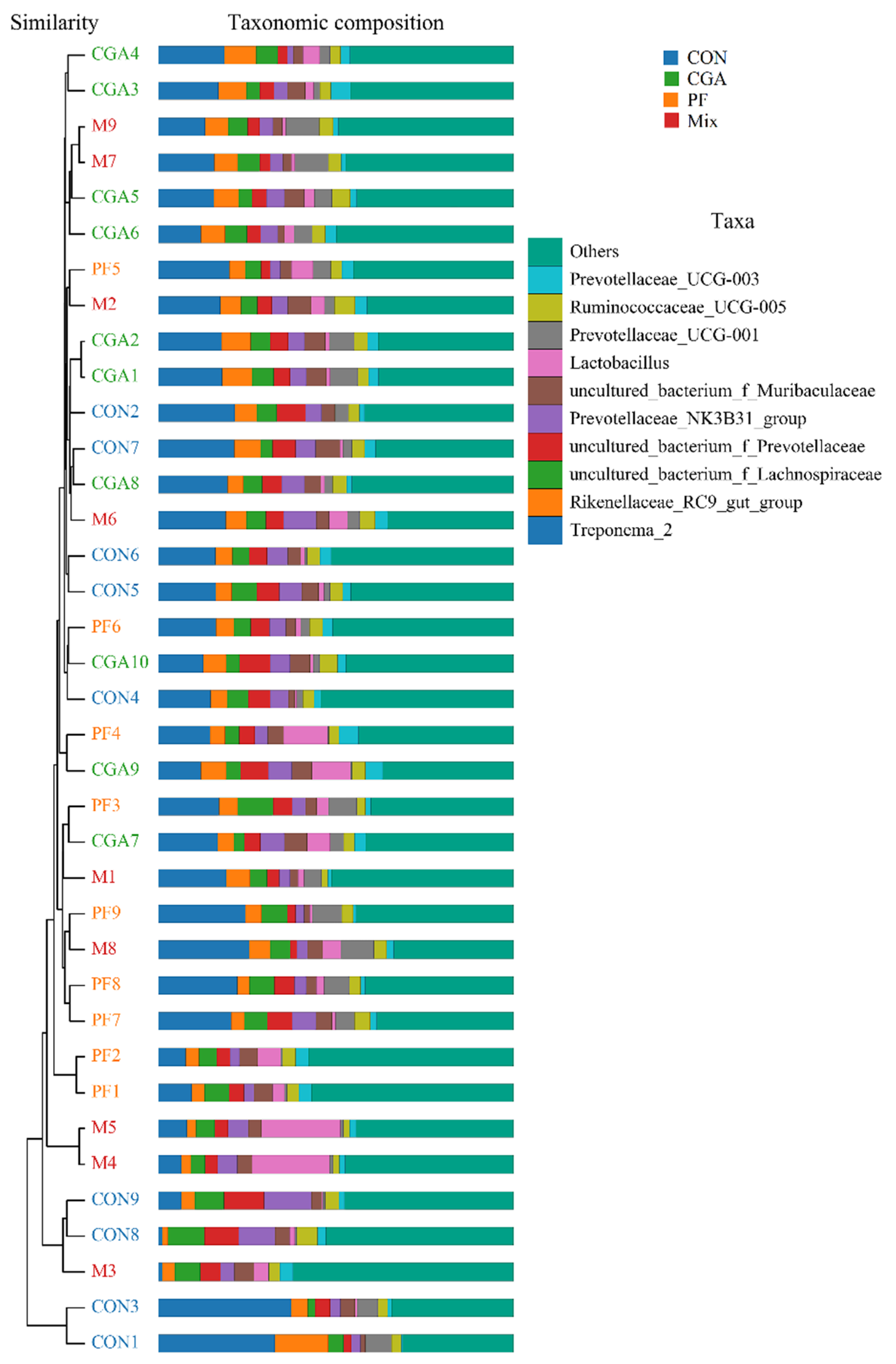
3.2. Validity Analysis of Fecal Microbes
3.3. Operational Taxonomic Units of Rectal Microbes in Finishing Pigs
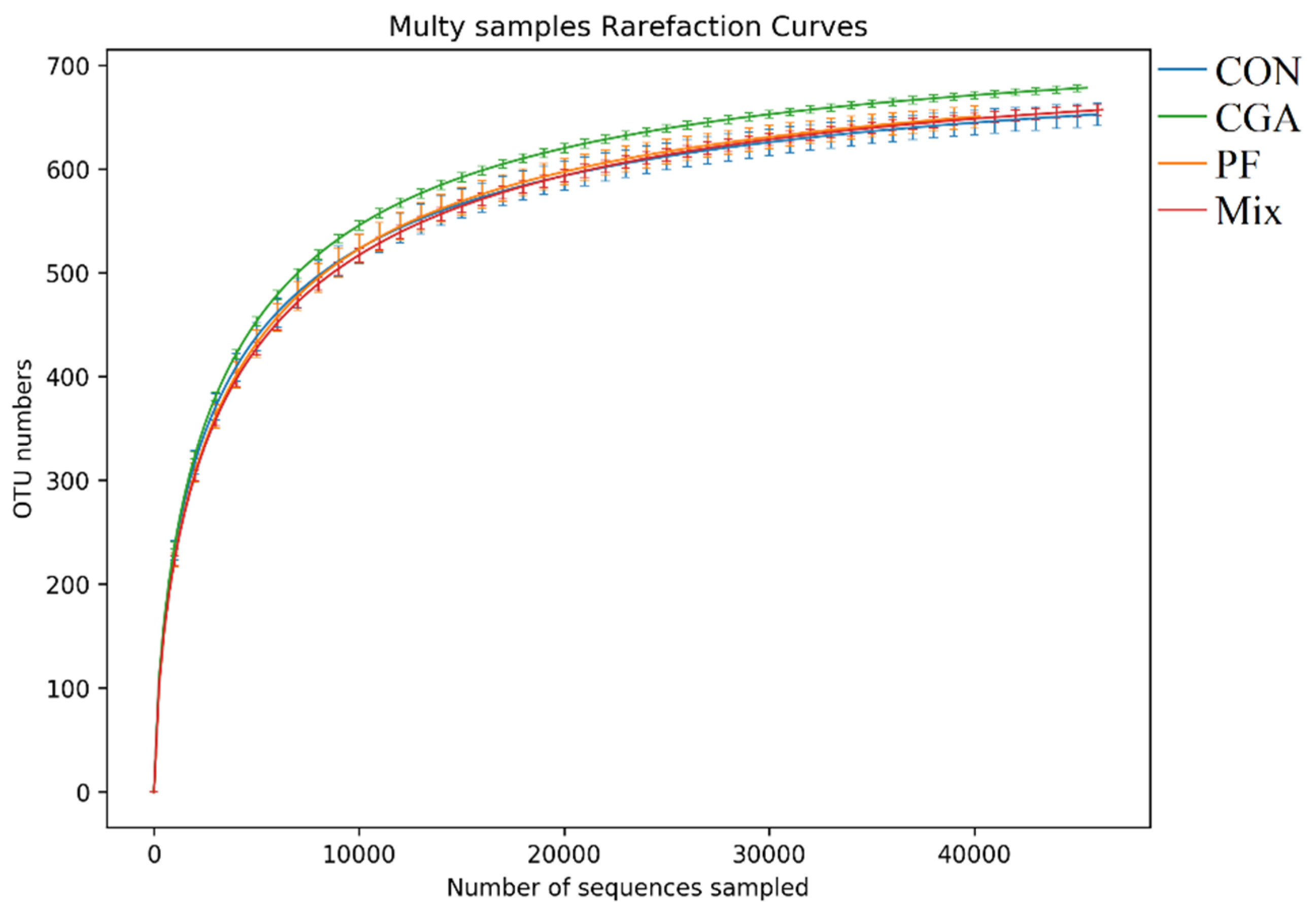
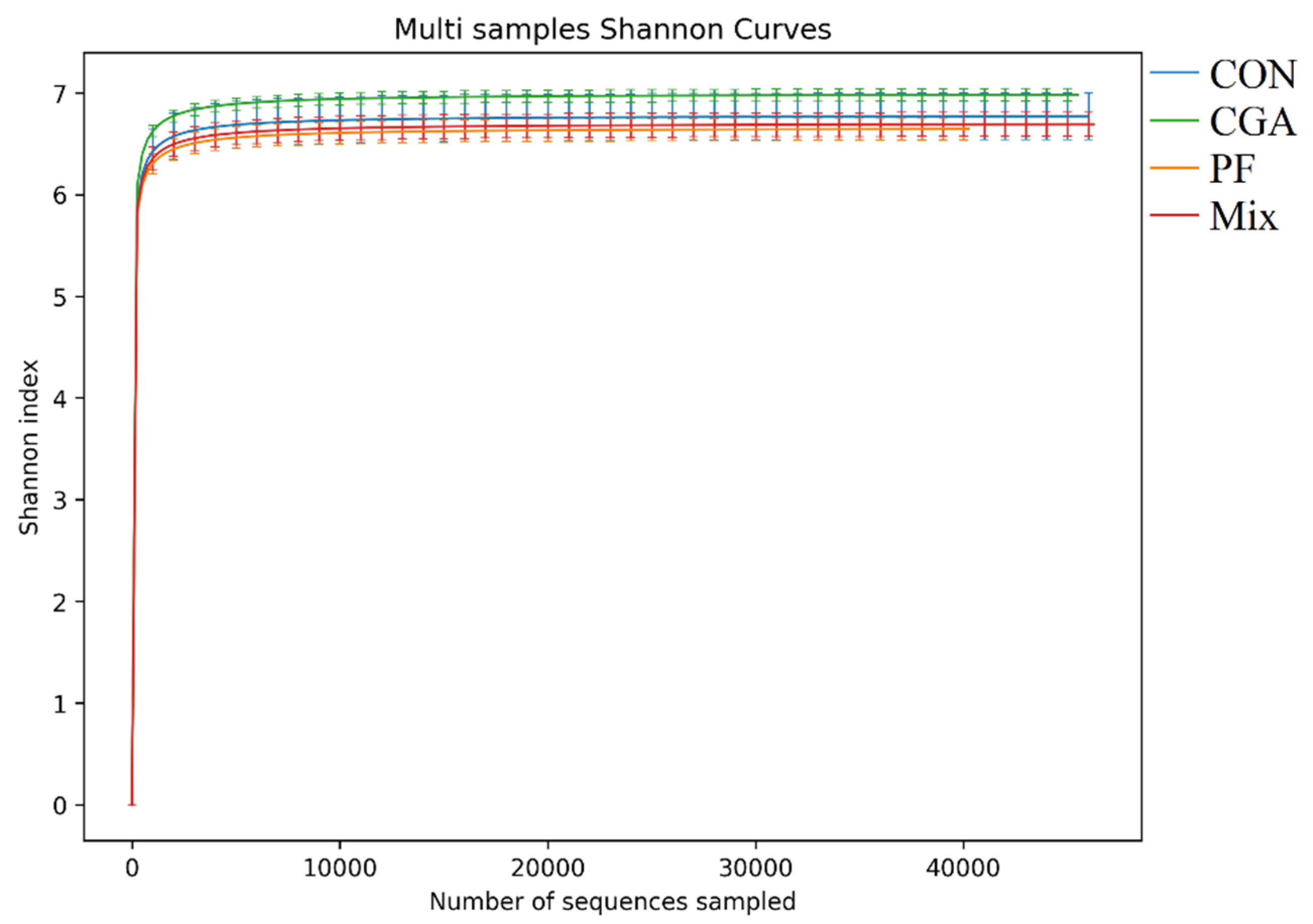
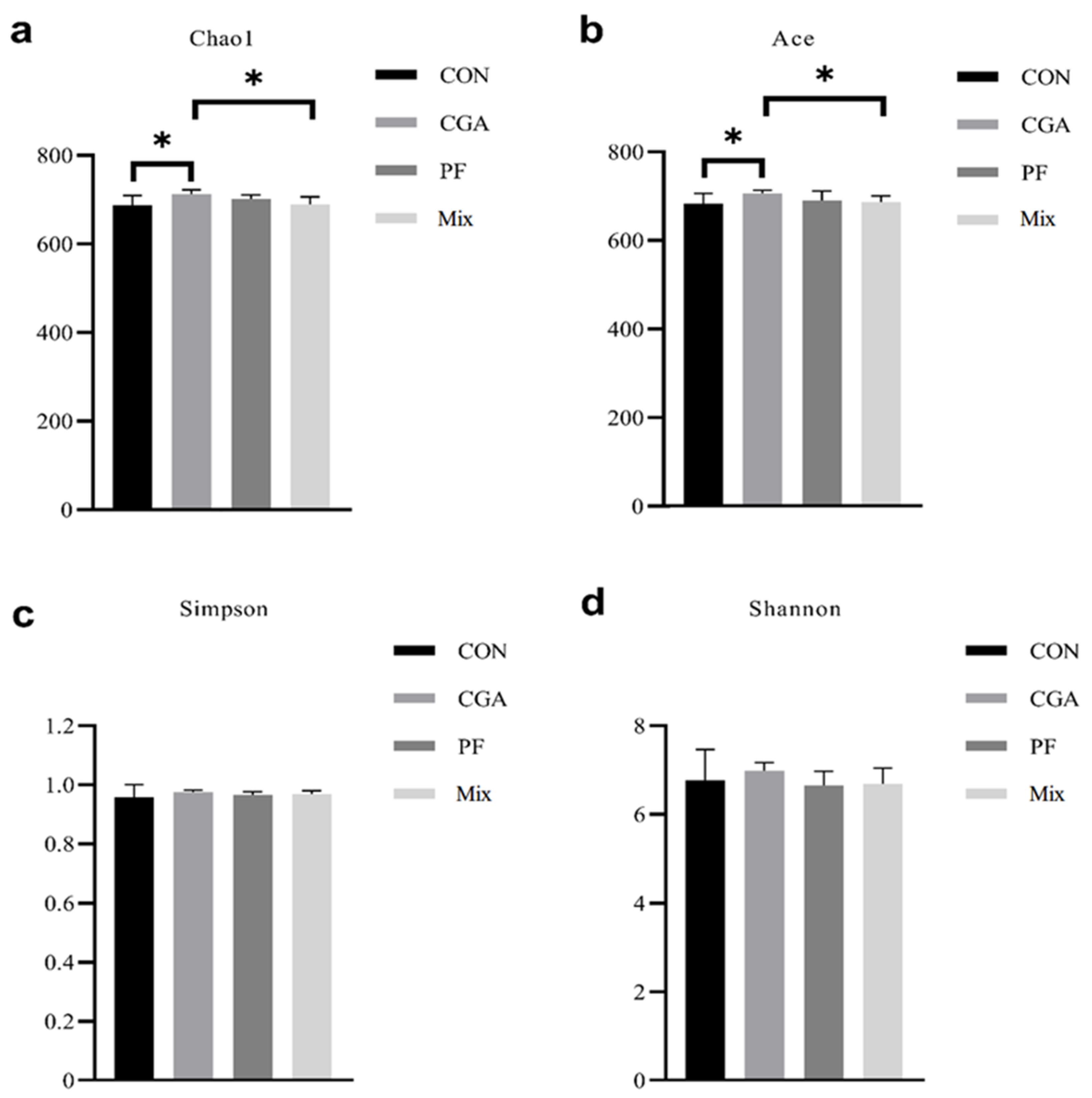
3.4. Alpha Diversity Analysis
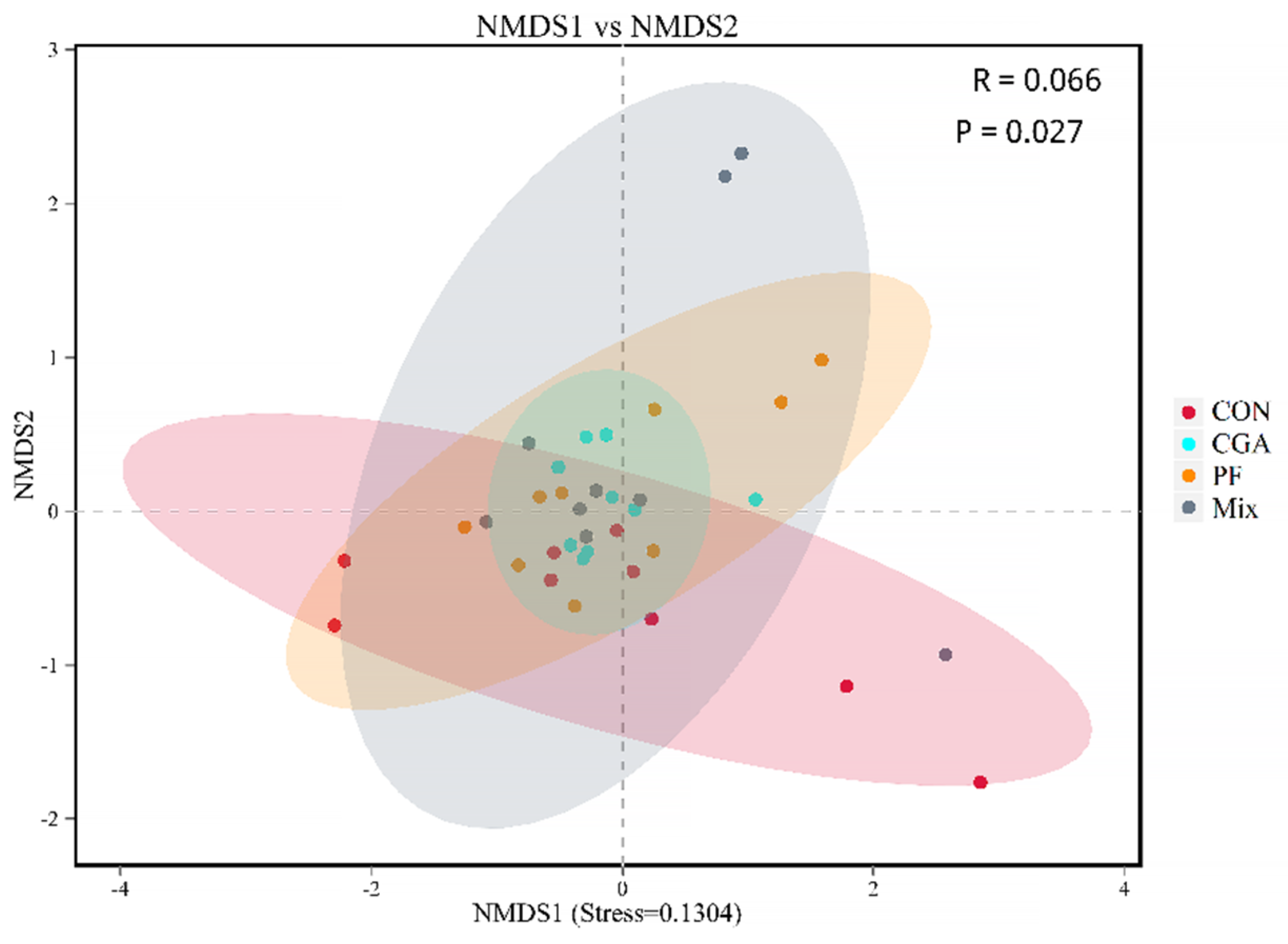
3.5. Beta Diversity Analysis
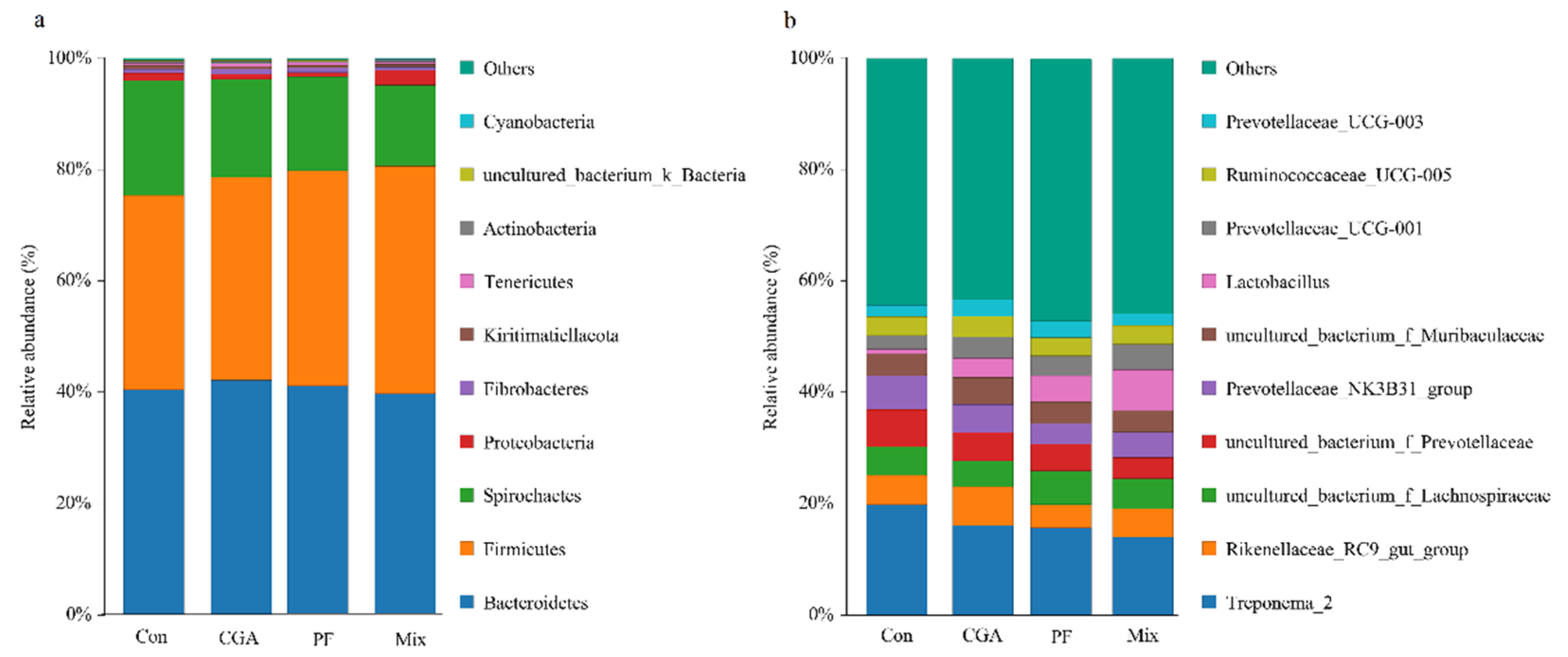
3.6. Annotation and Taxonomic Analysis of Species
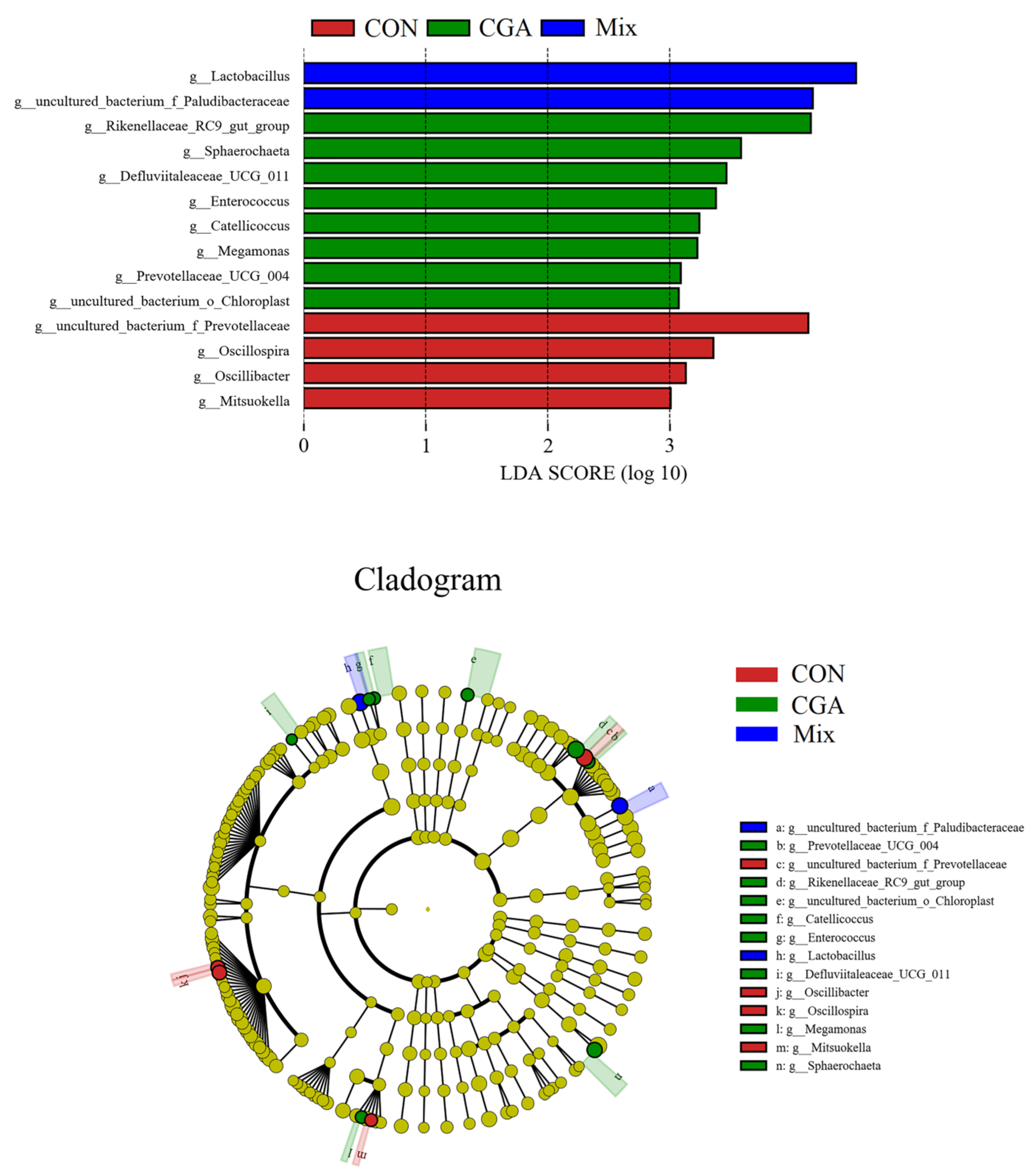
3.7. LEfSe Analysis of Different Groups
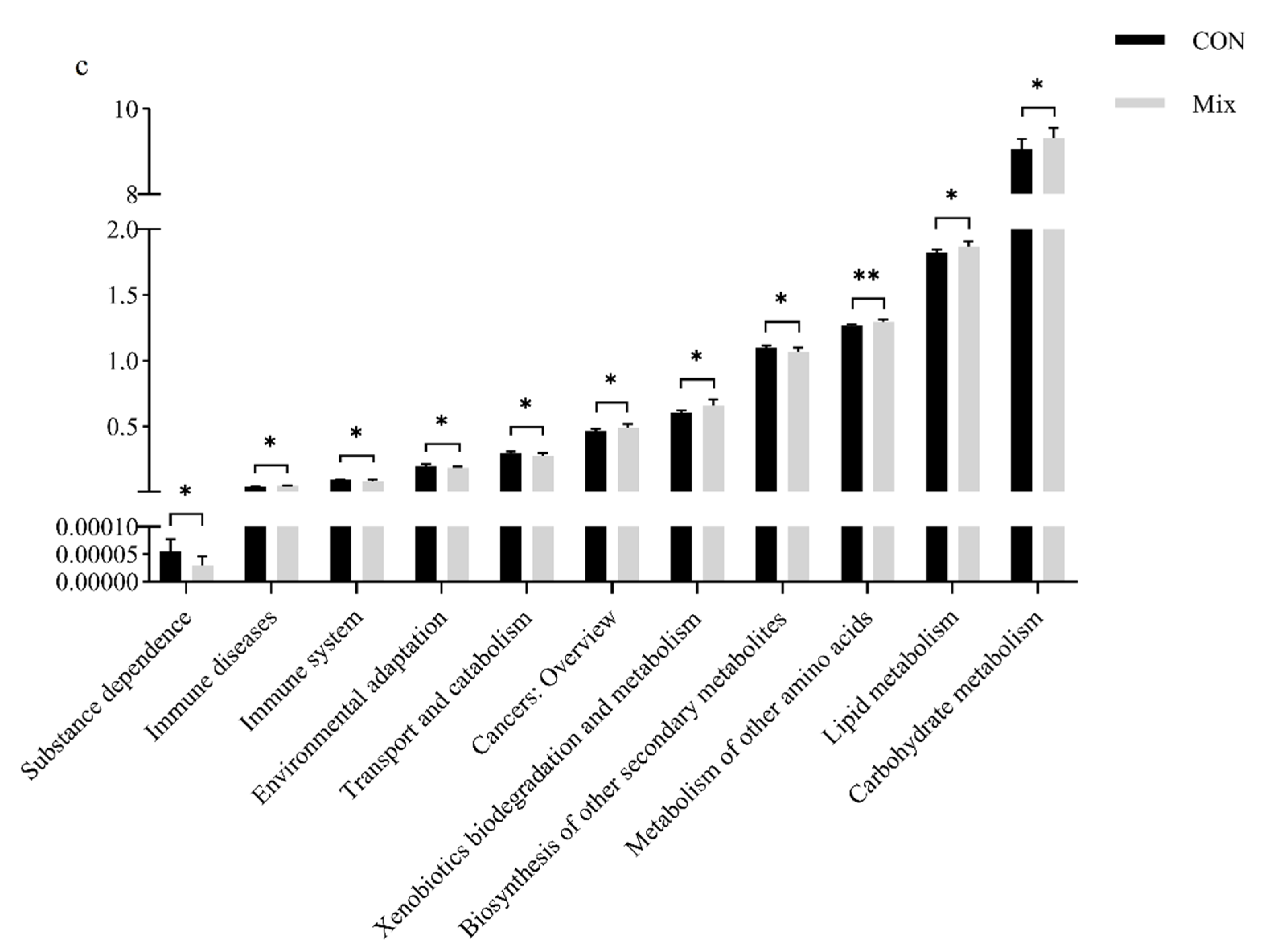
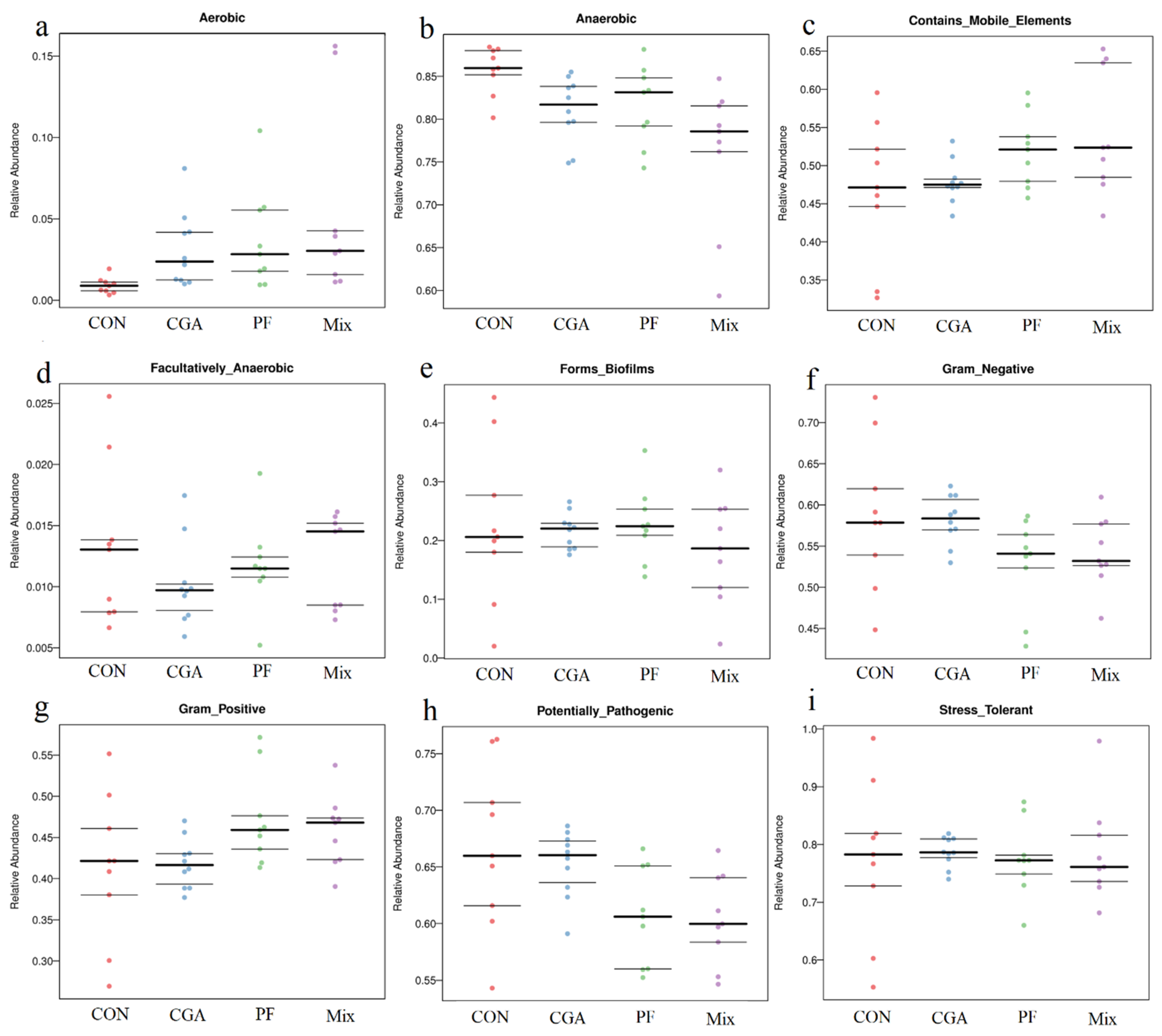
3.8. Functional Gene Prediction Analysis
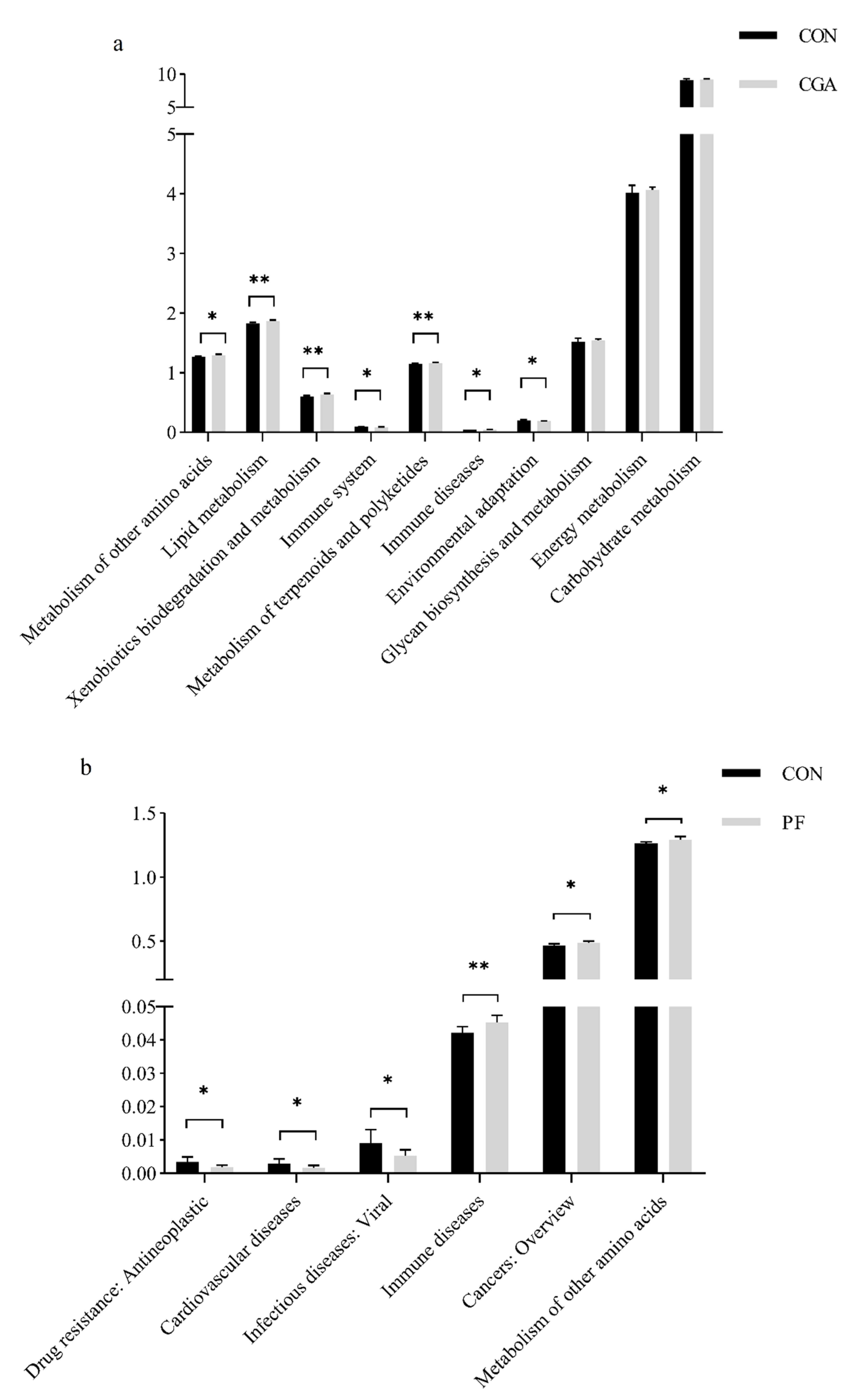
3.8.1. PICRUSt2 Function Prediction
3.8.2. Prediction of BugBase Phenotype
3.9. Effect of L. japonica and Radix Puerariae Crude Extracts on Intestinal ATTD in Finishing Pigs
4. Discussion
4.1. Effects of L. japonica Crude Extract and Radix Puerariae Crude Extract on the Production Performance of Finishing Pigs
4.2. Effects of L. japonica and Radix Puerariae Crude Extracts on the Fecal Microbiome in Finishing Pigs
4.3. Effects of L. japonica and Radix Puerariae Crude Extracts on ATTD in Finishing Pigs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbosa, C.; Trebosc, V.; Kemmer, C.; Rosenstiel, P.; Beardmore, R.; Schulenburg, H.; Jansen, G. Alternative Evolutionary Paths to Bacterial Antibiotic Resistance Cause Distinct Collateral Effects. Mol. Biol. Evol. 2017, 34, 2229–2244. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, Z.; Zhou, Y.; Tan, J.; Sun, H.; Sun, D.; Mu, Y.; Peng, J.; Wei, H. Effects of different amino acid levels and a carvacrol-thymol blend on growth performance and intestinal health of weaned pigs. J. Anim. Sci. Biotechnol. 2022, 13, 22. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Pan, H.; Li, M.; Miao, X.; Ding, H. Lonicera japonica Thunb.: Ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2011, 138, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Jaganath, I.B.; Ludwig, I.A.; Crozier, A. Chlorogenic acids and the acyl-quinic acids: Discovery, biosynthesis, bioavailability and bioactivity. Nat. Prod. Rep. 2017, 34, 1391–1421. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.J.; Kim, Y.H.; Kang, Y.H.; Choi, M.H.; Lee, J.H. Antioxidant and antibacterial effects of medicinal plants and their stick-type medicinal concentrated beverages. Food Sci. Biotechnol. 2020, 29, 1413–1423. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Liu, M.; Yu, Q.; Yi, Y.; Xiao, H.; Putra, D.F.; Ke, K.; Zhang, Q.; Li, P. Antiviral activities of Lonicera japonica Thunb. Components against grouper iridovirus in vitro and in vivo. Aquaculture 2020, 519, 734882. [Google Scholar] [CrossRef]
- Liu, C.M.; Ma, J.Q.; Sun, Y.Z. Protective role of puerarin on lead-induced alterations of the hepatic glutathione antioxidant system and hyperlipidemia in rats. Food Chem. Toxicol. 2011, 49, 3119–3127. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, H.; Yang, H.; Nie, L.; Zang, H. Rapid determination of major bioactive isoflavonoid compounds during the extraction process of kudzu (Pueraria lobata) by near-infrared transmission spectroscopy. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2015, 137, 1403–1408. [Google Scholar] [CrossRef]
- Zhou, Y.X.; Zhang, H.; Peng, C. Puerarin: A review of pharmacological effects. Phytother. Res. 2014, 28, 961–975. [Google Scholar] [CrossRef]
- Cherdshewasart, W.; Sutjit, W. Correlation of antioxidant activity and major isoflavonoid contents of the phytoestrogen-rich Pueraria mirifica and Pueraria lobata tubers. Phytomedicine 2008, 15, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yu, J.; Shi, J. Management of Diabetes Mellitus with Puerarin, a Natural Isoflavone from Pueraria lobata. Am. J. Chin. Med. 2018, 46, 1771–1789. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jeon, Y.D.; Lee, Y.M.; Kim, D.K. The suppressive effect of puerarin on atopic dermatitis-like skin lesions through regulation of inflammatory mediators in vitro and in vivo. Biochem. Biophys. Res. Commun. 2018, 498, 707–714. [Google Scholar] [CrossRef]
- Wang, Y.; Sheng, H.F.; He, Y.; Wu, J.Y.; Jiang, Y.X.; Tam, N.F.; Zhou, H.W. Comparison of the levels of bacterial diversity in freshwater, intertidal wetland, and marine sediments by using millions of illumina tags. Appl. Environ. Microbiol. 2012, 78, 8264–8271. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Ba, Q.; Li, M.; Chen, P.; Huang, C.; Duan, X.; Lu, L.; Li, J.; Chu, R.; Xie, D.; Song, H.; et al. Sex-Dependent Effects of Cadmium Exposure in Early Life on Gut Microbiota and Fat Accumulation in Mice. Environ. Health Perspect. 2017, 125, 437–446. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Yeung, M.; Saingam, P.; Xu, Y.; Xi, J. Low-dosage ozonation in gas-phase biofilter promotes community diversity and robustness. Microbiome 2021, 9, 14. [Google Scholar] [CrossRef]
- Li, N.; Zuo, B.; Huang, S.; Zeng, B.; Han, D.; Li, T.; Liu, T.; Wu, Z.; Wei, H.; Zhao, J.; et al. Spatial heterogeneity of bacterial colonization across different gut segments following inter-species microbiota transplantation. Microbiome 2020, 8, 161. [Google Scholar] [CrossRef]
- Ward, T.; Larson, J.; Meulemans, J.; Hillmann, B.; Lynch, J.; Sidiropoulos, D.; Spear, J.R.; Caporaso, G.; Blekhman, R.; Knight, R.; et al. BugBase predicts organism-level microbiome phenotypes. bioRxiv 2017, 133462. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; Green, E.D.; et al. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhao, J.; Deng, W.; Li, K.; Liu, H. Effects of chlorogenic acid-enriched extract from Eucommia ulmoides Oliver leaf on growth performance and quality and oxidative status of meat in finishing pigs fed diets containing fresh or oxidized corn oil. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Y.; Chen, D.; Yu, B.; Zheng, P.; Mao, X.; Luo, Y.; Li, Y.; He, J. Dietary chlorogenic acid supplementation affects gut morphology, antioxidant capacity and intestinal selected bacterial populations in weaned piglets. Food Funct. 2018, 9, 4968–4978. [Google Scholar] [CrossRef]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Mountzouris, K.C.; Balaskas, C.; Fava, F.; Tuohy, K.M.; Gibson, G.R.; Fegeros, K. Profiling of composition and metabolic activities of the colonic microflora of growing pigs fed diets supplemented with prebiotic oligosaccharides. Anaerobe 2006, 12, 178–185. [Google Scholar] [CrossRef]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef]
- Castillo, M.; Martín-Orúe, S.M.; Anguita, M.; Pérez, J.F.; Gasa, J. Adaptation of gut microbiota to corn physical structure and different types of dietary fibre. Livestock Sci. 2007, 109, 149–152. [Google Scholar] [CrossRef]
- Wei-ping, Y.; Jian-Gang, W.; Bin-Yun, C. Composition and Influence Factors of Pigs’ Gut Microbial Community. China Anim. Husb. J. 2017, 53, 12–16. [Google Scholar] [CrossRef]
- Min, W.; Zuohua, L.; Renli, Q. Growth and Development of Muscle in Animals Controlled by Intestinal Microbes. Chin. J. Anim. Nutr. 2019, 31, 3976–3982. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Petri, D.; Hill, J.E.; Van Kessel, A.G. Microbial succession in the gastrointestinal tract (GIT) of the preweaned pig. Livestock Sci. 2010, 133, 107–109. [Google Scholar] [CrossRef]
- Sood, A.; Midha, V.; Makharia, G.K.; Ahuja, V.; Singal, D.; Goswami, P.; Tandon, R.K. The probiotic preparation, VSL#3 induces remission in patients with mild-to-moderately active ulcerative colitis. Clin. Gastroenterol. Hepatol. 2009, 7, 1202–1209. [Google Scholar] [CrossRef]
- Xiaoman, Y.; Fengyuan, Z.; Fu, X.; Zhenzhu, D.; Chenmin, Y.; Shisheng, L. Honeysuckle Extract Promotes Host Health by Improving Intestinal Microbes and Enhancing Intestinal Mucosal Immunity. Genom. Appl. Biol. 2020, 39, 1257–1263. [Google Scholar] [CrossRef]
- Chen, J.; Yu, B.; Chen, D.; Zheng, P.; Luo, Y.; Huang, Z.; Luo, J.; Mao, X.; Yu, J.; He, J. Changes of porcine gut microbiota in response to dietary chlorogenic acid supplementation. Appl. Microbiol. Biotechnol. 2019, 103, 8157–8168. [Google Scholar] [CrossRef]
- Ferreyra, J.A.; Wu, K.J.; Hryckowian, A.J.; Bouley, D.M.; Weimer, B.C.; Sonnenburg, J.L. Gut microbiota-produced succinate promotes C. difficile infection after antibiotic treatment or motility disturbance. Cell Host Microbe 2014, 16, 770–777. [Google Scholar] [CrossRef]
- Duncan, S.H.; Louis, P.; Thomson, J.M.; Flint, H.J. The role of pH in determining the species composition of the human colonic microbiota. Environ. Microbiol. 2009, 11, 2112–2122. [Google Scholar] [CrossRef]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Huang, C.; Song, P.; Fan, P.; Hou, C.; Thacker, P.; Ma, X. Dietary Sodium Butyrate Decreases Postweaning Diarrhea by Modulating Intestinal Permeability and Changing the Bacterial Communities in Weaned Piglets. J. Nutr. 2015, 145, 2774–2780. [Google Scholar] [CrossRef] [PubMed]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; Ian McKenzie, C.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [PubMed]
- Diao, H.; Jiao, A.R.; Yu, B.; Mao, X.B.; Chen, D.W. Gastric infusion of short-chain fatty acids can improve intestinal barrier function in weaned piglets. Genes Nutr. 2019, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, K.; Chen, H.; Su, Y.; Zhu, W. Caecal infusion of the short-chain fatty acid propionate affects the microbiota and expression of inflammatory cytokines in the colon in a fistula pig model. Microb. Biotechnol. 2018, 11, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef]
- Precup, G.; Vodnar, D.C. Gut Prevotella as a possible biomarker of diet and its eubiotic versus dysbiotic roles: A comprehensive literature review. Br. J. Nutr. 2019, 122, 131–140. [Google Scholar] [CrossRef]
- Wang, L.L.; Guo, H.H.; Huang, S.; Feng, C.L.; Han, Y.X.; Jiang, J.D. Comprehensive evaluation of SCFA production in the intestinal bacteria regulated by berberine using gas-chromatography combined with polymerase chain reaction. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1057, 70–80. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-Gut Microbiota Metabolic Interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.A.; Gálvez, E.J.C.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and cultivation study of Muribaculaceae reveals novel species, host preference, and functional potential of this yet undescribed family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef]
- Lin, H.; Haoxuan, D.; Jie, F. Effects of Eucommia ulmoides Oliv. Extract on Growth Performance and Nutrient Digestion and Utilization of Yellow Feather Broilers. Chin. J. Anim. Sci. 2020, 56, 144–149. [Google Scholar] [CrossRef]
- Miller, C.A.; Holm, H.C.; Horstmann, L.; George, J.C.; Fredricks, H.F.; Van Mooy, B.A.S.; Apprill, A. Coordinated transformation of the gut microbiome and lipidome of bowhead whales provides novel insights into digestion. Isme J. 2020, 14, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Engevik, A.C.; Kaji, I.; Engevik, M.A.; Meyer, A.R.; Weis, V.G.; Goldstein, A.; Hess, M.W.; Müller, T.; Koepsell, H.; Dudeja, P.K.; et al. Loss of MYO5B Leads to Reductions in Na Na+ Absorption with Maintenance of CFTR-Dependent Cl− Secretion in Enterocytes. Gastroenterology. 2018, 155, 1883–1897. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.; Yu, B.; Chen, D.; Mao, X.; Zheng, P.; Luo, J.; He, J. Dietary chlorogenic acid improves growth performance of weaned pigs through maintaining antioxidant capacity and intestinal digestion and absorption function. J. Anim. Sci. 2018, 96, 1108–1118. [Google Scholar] [CrossRef]
- Jing-cheng, B.; Xin-ying, W.; Jie-shou, L. Research progresses of intestinal alkaline phosphatase in intestinal barrier maintenance. Parenter. Enteral Nutr. 2015, 4, 244–247. [Google Scholar] [CrossRef]
- Goldberg, R.F.; Austen, W.G., Jr.; Zhang, X.; Munene, G.; Mostafa, G.; Biswas, S.; McCormack, M.; Eberlin, K.R.; Nguyen, J.T.; Tatlidede, H.S.; et al. Intestinal alkaline phosphatase is a gut mucosal defense factor maintained by enteral nutrition. Proc. Natl. Acad. Sci. USA 2008, 105, 3551–3556. [Google Scholar] [CrossRef]
- Malo, M.S.; Alam, S.N.; Mostafa, G.; Zeller, S.J.; Johnson, P.V.; Mohammad, N.; Chen, K.T.; Moss, A.K.; Ramasamy, S.; Faruqui, A.; et al. Intestinal alkaline phosphatase preserves the normal homeostasis of gut microbiota. Gut 2010, 59, 1476–1484. [Google Scholar] [CrossRef]
- Luo, G.; Ling, N.; Nannipieri, P.; Chen, H.; Raza, W.; Wang, M.; Guo, S.; Shen, Q. Long-term fertilisation regimes affect the composition of the alkaline phosphomonoesterase encoding microbial community of a vertisol and its derivative soil fractions. Biol. Fertil. Soils 2017, 53, 375–388. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Contents |
|---|---|
| Corn, % | 63.38 |
| Wheat Bran, % | 14.40 |
| Soybean meal, % | 13.00 |
| Distillers dried grains with solubles (DDGS), % | 5.0 |
| CaHPO4, % | 1.20 |
| Limestone, % | 0.90 |
| NaCl, % | 0.30 |
| L-lysine hydrochloride (98.5%), % | 0.27 |
| L-threonine (97.5%), % | 0.04 |
| DL-methionine (99%), % | 0.01 |
| Choline chloride, % | 0.10 |
| Premix 1,2, % | 1.40 |
| Nutrient levels | |
| Dry matter, % | 89.62 |
| Gross energy (GE) 3, MJ/kg | 16.63 |
| Crude protein, % | 16.09 |
| Crude fiber, % | 4.80 |
| Ca, % | 0.74 |
| Total P, % | 0.69 |
| Lysine (Lys), % | 0.90 |
| Threonine (Thr), % | 0.58 |
| Methionine + Cysteine 3, % | 0.63 |
| Items | Con | CGA | PF | Mix |
|---|---|---|---|---|
| Initial weight (kg) | 94.44 ± 0.32 | 92.56 ± 0.75 | 93.28 ± 1.75 | 92.47 ± 0.97 |
| Final weight (kg) | 130.91 ± 1.09 | 129.67 ± 0.30 | 131.47 ± 2.27 | 131.94 ± 1.81 |
| ADFI (kg) | 3.00 ± 0.02 | 3.04 ± 0.03 | 3.03 ± 0.03 | 3.05 ± 0.03 |
| ADG (kg) | 0.83 ± 0.02 b | 0.87 ± 0.03 ab | 0.86 ± 0.02 ab | 0.90 ± 0.02 a |
| FCR | 3.63 ± 0.10 a | 3.50 ± 0.15 ab | 3.55 ± 0.12 ab | 3.40 ± 0.06 b |
| Item | CON | CGA | PF | Mix |
|---|---|---|---|---|
| Bacteroidetes | 0.4162 ± 0.0623 | 0.4179 ± 0.0212 | 0.4059 ± 0.0526 | 0.3971 ± 0.028 |
| Firmicutes | 0.3533 ± 0.0623 b | 0.3665 ± 0.0163 ab | 0.3815 ± 0.0339 ab | 0.4072 ± 0.0557 a |
| Spirochaetes | 0.1943 ± 0.1141 | 0.1745 ± 0.0262 | 0.1788 ± 0.0551 | 0.1476 ± 0.0756 |
| Proteobacteria | 0.0114 ± 0.0115 | 0.0095 ± 0.0054 | 0.0080 ± 0.0033 | 0.0266 ± 0.0349 |
| Fibrobacteres | 0.0064 ± 0.0046 | 0.0082 ± 0.008 | 0.0084 ± 0.0089 | 0.0060 ± 0.0053 |
| Kiritimatiellaeota | 0.0055 ± 0.004 | 0.0050 ± 0.0019 | 0.0040 ± 0.0027 | 0.0045 ± 0.0022 |
| Tenericutes | 0.0051 ± 0.0051 | 0.0042 ± 0.0027 | 0.0044 ± 0.0017 | 0.0045 ± 0.0021 |
| Actinobacteria | 0.0033 ± 0.0019 | 0.0042 ± 0.0023 | 0.0034 ± 0.0015 | 0.0032 ± 0.0008 |
| uncultured_bacterium_k_Bacteria | 0.0009 ± 0.0013 | 0.0021 ± 0.0021 | 0.0015 ± 0.0012 | 0.001 ± 0.0011 |
| Cyanobacteria | 0.0007 ± 0.0006 | 0.0018 ± 0.001 | 0.0009 ± 0.0007 | 0.0006 ± 0.0003 |
| Others | 0.003 ± 0.0016 | 0.0026 ± 0.0009 | 0.0029 ± 0.0013 | 0.0019 ± 0.0006 |
| Item | CON | CGA | PF | Mix |
|---|---|---|---|---|
| Treponema-2 | 0.19 ± 0.11 | 0.16 ± 0.03 | 0.17 ± 0.06 | 0.14 ± 0.08 |
| Rikenellaceae-RC9-gut-group | 0.06 ± 0.04 ab | 0.07 ± 0.02 a | 0.04 ± 0.01 b | 0.05 ± 0.02 ab |
| uncultured-bacterium-f-Lachnospiraceae | 0.06 ± 0.03 | 0.05 ± 0.01 | 0.06 ± 0.02 | 0.05 ± 0.01 |
| uncultured-bacterium-f-Prevotellaceae | 0.07 ± 0.03 a | 0.05 ± 0.02 ab | 0.05 ± 0.01 b | 0.04 ± 0.01 b |
| Prevotellaceae-NK3B31-group | 0.06 ± 0.03 a | 0.05 ± 0.02 ab | 0.04 ± 0.01 b | 0.05 ± 0.02 ab |
| uncultured-bacterium-f-Muribaculaceae | 0.04 ± 0.02 | 0.05 ± 0.01 | 0.04 ± 0.01 | 0.04 ± 0.01 |
| Lactobacillus | 0.01 ± 0.01 b | 0.03 ± 0.03 ab | 0.04 ± 0.04 ab | 0.07 ± 0.08 a |
| Prevotellaceae-UCG−001 | 0.03 ± 0.02 | 0.04 ± 0.02 | 0.04 ± 0.03 | 0.05 ± 0.04 |
| Ruminococcaceae-UCG−005 | 0.04 ± 0.01 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 |
| Prevotellaceae-UCG−003 | 0.02 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.02 ± 0.01 |
| Others | 0.44 ± 0.08 | 0.43 ± 0.04 | 0.46 ± 0.07 | 0.46 ± 0.09 |
| Items | CON | CGA | PF | Mix |
|---|---|---|---|---|
| Crude protein (CP) | 86.62 ± 0.99 b | 88.07 ± 1.61 a | 88.11 ± 1.07 a | 87.5 ± 0.62 ab |
| Crude fiber (CF) | 46.23 ± 4.23 | 46.70 ± 3.82 | 46.42 ± 3.44 | 46.50 ± 2.97 |
| Ether extract (EE) | 74.37 ± 1.76 | 75.14 ± 1.69 | 75.65 ± 1.58 | 74.97 ± 1.57 |
| Ca | 57.41 ± 5.76 | 57.38 ± 3.27 | 56.89 ± 1.95 | 58.95 ± 6.11 |
| Total phosphorus (TP) | 55.84 ± 2.71 b | 59.51 ± 1.38 a | 58.50 ± 2.37 ab | 56.39 ± 2.39 b |
| Gross energy (GE) | 88.54 ±0.94 | 88.8 ± 0.52 | 88.52 ± 0.77 | 88.60 ± 0.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Li, N.; Zheng, Z.; Zhang, C.; Liu, Z.; Song, C.; Yan, J.; Mu, S. Influence of Lonicera japonica and Radix Puerariae Crude Extracts on the Fecal Microbiome and Nutrient Apparent Digestibility of Finishing Pigs. Animals 2022, 12, 2109. https://doi.org/10.3390/ani12162109
Liu Z, Li N, Zheng Z, Zhang C, Liu Z, Song C, Yan J, Mu S. Influence of Lonicera japonica and Radix Puerariae Crude Extracts on the Fecal Microbiome and Nutrient Apparent Digestibility of Finishing Pigs. Animals. 2022; 12(16):2109. https://doi.org/10.3390/ani12162109
Chicago/Turabian StyleLiu, Zhonghao, Ning Li, Zi Zheng, Chunhua Zhang, Zhengqun Liu, Chunling Song, Jun Yan, and Shuqin Mu. 2022. "Influence of Lonicera japonica and Radix Puerariae Crude Extracts on the Fecal Microbiome and Nutrient Apparent Digestibility of Finishing Pigs" Animals 12, no. 16: 2109. https://doi.org/10.3390/ani12162109