
Diurnal and Nocturnal Behaviour of Cheetahs (Acinonyx jubatus) and Lions (Panthera leo) in Zoos

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection and Behavioural States
2.2. Data Analyses
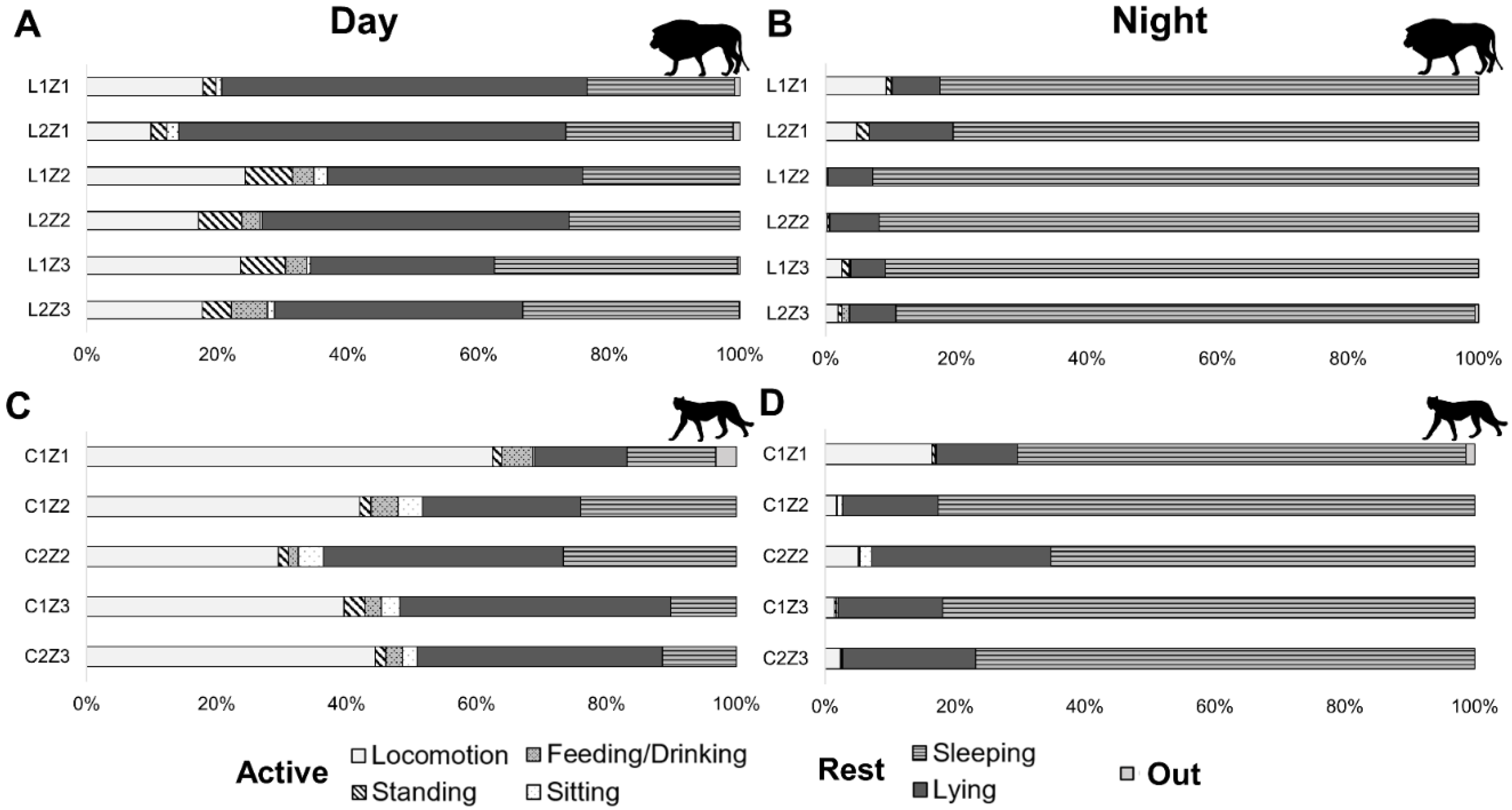
3. Results
4. Discussion
4.1. Behavioural Rhythmicity over 24 Hours
4.2. Sleep Behaviour
4.3. Food as Decisive Factor for the Behavioural Rhythm
4.4. Light and Season Shape Activity and Rest Behaviour
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mistlberger, R.E. Neurobiology of food anticipatory circadian rhythms. Physiol. Behav. 2011, 104, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Aschoff, J.; Pohl, H. Phase relations between a circadian rhythm and its zeitgeber within the range of entrainment. Naturwissenschaften 1978, 65, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, G.; Broekhuis, F.; McNutt, J.W.; Turnbull, L.A.; MacDonald, D.W.; Schmid, B. Fear of the dark or dinner by moonlight? Reduced temporal partitioning among Africa’s large carnivores. Ecology 2012, 93, 2590–2599. [Google Scholar] [CrossRef]
- Gillman, A.G.; Rebec, G.V.; Pecoraro, N.C.; Kosobud, A.E.K. Circadian entrainment by food and drugs of abuse. Behav. Process. 2019, 165, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Merrow, M.; Spoelstra, K.; Roenneberg, T. The circadian cycle: Daily rhythms from behaviour to genes. EMBO Rep. 2005, 6, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Rowcliffe, J.M.; Kays, R.; Kranstauber, B.; Carbone, C.; Jansen, P.A. Quantifying levels of animal activity using camera trap data. Methods Ecol. Evol. 2014, 5, 1170–1179. [Google Scholar] [CrossRef]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Yoakum, E.; Andrews, N. Seasonal and Daily Activity of Two Zoo-Housed Grizzly Bears (Ursus arctos horribilis). JZBG 2020, 1, 1–12. [Google Scholar] [CrossRef]
- Hayward, M.W.; Slotow, R. Temporal partitioning of activity in large African carnivores: Tests of multiple hypotheses. S. Afr. J. Wildl. Res. 2009, 39, 109–125. [Google Scholar] [CrossRef]
- Mallapur, A.; Chellam, R. Environmental influences on stereotypy and the activity budget of Indian leopards (Panthera pardus) in four zoos in Southern India. Zoo Biol. 2002, 21, 585–595. [Google Scholar] [CrossRef]
- Rees, P.A. Activity budgets and the relationship between feeding and stereotypic behaviors in Asian elephants (Elephas maximus) in a Zoo. Zoo Biol. 2009, 28, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.; Robert, R. Evaluating the activity patterns and enclosure usage of a little-studied zoo species, the sitatunga (Tragelaphus spekii). JZAR 2013, 1, 14–19. [Google Scholar] [CrossRef]
- Ancoli-Israel, S.; Cole, R.; Alessi, C.; Chambers, M.; Moorcroft, W.; Pollak, C.P. The role of actigraphy in the study of sleep and circadian rhythms. Sleep 2003, 26, 342–392. [Google Scholar] [CrossRef] [PubMed]
- Berger, A. Activity patterns, chronobiology and the assessment of stress and welfare in zoo and wild animals. Int. Zoo Yearb. 2011, 45, 80–90. [Google Scholar] [CrossRef]
- Favreau, A.; Richard-Yris, M.-A.; Bertin, A.; Houdelier, C.; Lumineau, S. Social influences on circadian behavioural rhythms in vertebrates. Anim. Behav. 2009, 77, 983–989. [Google Scholar] [CrossRef]
- Rattenborg, N.C.; de La Iglesia, H.O.; Kempenaers, B.; Lesku, J.A.; Meerlo, P.; Scriba, M.F. Sleep research goes wild: New methods and approaches to investigate the ecology, evolution and functions of sleep. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160251. [Google Scholar] [CrossRef]
- Veasey, J.S.; Waran, N.K.; Young, R.J. On comparing the behaviour of zoo housed animals with the wild conspecifics as a welfare indicator. Anim. Welf. 1996, 5, 13–24. [Google Scholar]
- Dawkins, M.S. Using behaviour to assess animal welfare. Anim. Welf. 2004, 13, 3–7. [Google Scholar]
- Hutchins, M.; Wiese, R.J.; Smith, B. Introduction. In Scientific Foundations of Zoos and Aquariums; Kaufman, A.B., Bashaw, M.J., Maple, T.L., Eds.; Cambridge University Press: Cambridge, UK, 2018; pp. 1–42. [Google Scholar]
- Bennie, J.J.; Duffy, J.P.; Inger, R.; Gaston, K.J. Biogeography of time partitioning in mammals. Proc. Natl. Acad. Sci. USA 2014, 111, 13727–13732. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, H.; Wang, H.; Feng, J. Arms race of temporal partitioning between carnivorous and herbivorous mammals. Sci. Rep. 2018, 8, 1713. [Google Scholar] [CrossRef]
- Swanson, A.; Arnold, T.; Kosmala, M.; Forester, J.; Packer, C. In the absence of a “landscape of fear”: How lions, hyenas, and cheetahs coexist. Ecol. Evol. 2016, 6, 8534–8545. [Google Scholar] [CrossRef] [PubMed]
- Siegel, J.M. Do all animals sleep? Trends Neurosci. 2008, 31, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Bissett, C.; Parker, D.M.; Bernard, R.T.; Perry, T.W. Management-induced niche shift? The activity of cheetahs in the presence of lions. Afr. J. Wildl. Res. 2015, 45, 197–203. [Google Scholar] [CrossRef]
- Cajochen, C.; Münch, M.; Knoblauch, V.; Blatter, K.; Wirz-Justice, A. Age-related changes in the circadian and homeostatic regulation of human sleep. Chronobiol. Int. 2006, 23, 461–474. [Google Scholar] [CrossRef]
- Houser, A.; Somers, M.J.; Boast, L.K. Home range use of free-ranging cheetah on farm and conservation land in Botswana. S. Afr. J. Wildl. Res. 2009, 39, 11–22. [Google Scholar] [CrossRef]
- Lesku, J.A.; Roth, T.C.; Amlaner, C.J.; Lima, S.L. A phylogenetic analysis of sleep architecture in mammals: The integration of anatomy, physiology, and ecology. Am. Nat. 2006, 168, 441–453. [Google Scholar] [CrossRef]
- Steinmeyer, C.; Kempenaers, B.; Mueller, J.C. Testing for associations between candidate genes for circadian rhythms and individual variation in sleep behaviour in blue tits. Genetica 2012, 140, 219–228. [Google Scholar] [CrossRef]
- Stuber, E.F.; Dingemanse, N.J.; Kempenaers, B.; Mueller, J.C. Sources of intraspecific variation in sleep behaviour of wild great tits. Anim. Behav. 2015, 106, 201–221. [Google Scholar] [CrossRef]
- Berger, A.; Scheibe, K.-M.; Michaelis, S.; Streich, W.J. Evaluation of living conditions of free-ranging animals by automated chronobiological analysis of behavior. Behav. Res. Methods Instrum. Comput. 2003, 35, 458–466. [Google Scholar] [CrossRef]
- Breton, G.; Barrot, S. Influence of enclosure size on the distances covered and paced by captive tigers (Panthera tigris). Appl. Anim. Behav. Sci. 2014, 154, 66–75. [Google Scholar] [CrossRef]
- Broekhuis, F.; Grünewälder, S.; McNutt, J.W.; Macdonald, D.W. Optimal hunting conditions drive circalunar behavior of a diurnal carnivore. Behav. Ecol. 2014, 25, 1268–1275. [Google Scholar] [CrossRef]
- Croft, D.P.; Darden, S.K.; Wey, T.W. Current directions in animal social networks. Curr. Opin. Behav. Sci. 2016, 12, 52–58. [Google Scholar] [CrossRef]
- Hetem, R.S.; Mitchell, D.; De Witt, B.A.; Fick, L.G.; Maloney, S.K.; Meyer, L.C.R.; Fuller, A. Body temperature, activity patterns and hunting in free-living cheetah: Biologging reveals new insights. Integr. Zool. 2019, 14, 30–47. [Google Scholar] [CrossRef]
- Nouvellet, P.; Rasmussen, G.S.A.; MacDonald, D.W.; Courchamp, F. Noisy clocks and silent sunrises: Measurement methods of daily activity pattern. J. Zool. 2012, 286, 179–184. [Google Scholar] [CrossRef]
- Siegel, J.M. Clues to the functions of mammalian sleep. Nature 2005, 437, 1264–1271. [Google Scholar] [CrossRef]
- Siegel, J.M. Sleep in mammals: A state of adaptive inactivity. In Principles and Practice of Sleep Medicine, 6th ed.; Kryger, M.H., Roth, T., Dement, W.C., Eds.; Elsevier Health Sciences: Philadelphia, PA, USA, 2016; pp. 103–114. [Google Scholar]
- Wey, T.; Blumstein, D.T.; Shen, W.; Jordán, F. Social network analysis of animal behaviour: A promising tool for the study of sociality. Anim. Behav. 2008, 75, 333–344. [Google Scholar] [CrossRef]
- Mistlberger, R.E. Circadian food-anticipatory activity: Formal models and physiological mechanisms. Neurosci. Biobehav. Rev. 1994, 12, 171–195. [Google Scholar] [CrossRef]
- Campbell, S.S.; Tobler, I. Animal sleep a review of sleep duration across phylogeny. Neurosci. Biobehav. Rev. 1984, 8, 269–300. [Google Scholar] [CrossRef]
- Bloomsmith, M.A.; Lambeth, S.P. Effects of predictable versus unpredictable feeding schedules on chimpanzee behavior. Appl. Anim. Behav. Sci. 1995, 44, 65–74. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Timberlake, W. Mutual benefits of research collaborations between zoos and academic institutions. Zoo Biol. 2008, 27, 470–487. [Google Scholar] [CrossRef]
- Höttges, N.; Hjelm, M.; Hard, T.; Laska, M. How does feeding regime affect behaviour and activity in captive African lions (Panthera leo). JZAR 2019, 7, 117–125. [Google Scholar] [CrossRef]
- Waitt, C.; Buchanan-Smith, H.M. What time is feeding? How delays and anticipation of feeding schedules affect stump-tailed macaque behavior. Appl. Anim. Behav. Sci. 2001, 75, 75–85. [Google Scholar] [CrossRef]
- Shepherdson, D.J.; Carlstead, K.; Mellen, J.D.; Seidensticker, J. The influence of food presentation on the behavior of small cats in confined environments. Zoo Biol. 1993, 12, 203–216. [Google Scholar] [CrossRef]
- Lyons, J.; Young, R.J.; Deag, J.M. The effects of physical characteristics of the environment and feeding regime on the behavior of captive felids. Zoo Biol. 1997, 16, 71–83. [Google Scholar] [CrossRef]
- Brando, S.; Buchanan-Smith, H.M. The 24/7 approach to promoting optimal welfare for captive wild animals. Behav. Process. 2018, 156, 83–95. [Google Scholar] [CrossRef]
- Cagnacci, F.; Boitani, L.; Powell, R.A.; Boyce, M.S. Animal ecology meets GPS-based radiotelemetry: A perfect storm of opportunities and challenges. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2157–2162. [Google Scholar] [CrossRef]
- Gunner, R.M.; Wilson, R.P.; Holton, M.D.; Hopkins, P.; Bell, S.H.; Marks, N.J.; Bennett, N.C.; Ferreira, S.; Govender, D.; Viljoen, P.; et al. Decision rules for determining terrestrial movement and the consequences for filtering high-resolution global positioning system tracks: A case study using the African lion (Panthera leo). J. R. Soc. Interface 2022, 19, 20210692. [Google Scholar] [CrossRef]
- Wijers, M.; Trethowan, P.; Markham, A.; Du Preez, B.; Chamaillé-Jammes, S.; Loveridge, A.; Macdonald, D. Listening to lions: Animal-borne acoustic sensors improve bio-logger calibration and behaviour classification performance. Front. Ecol. Evol. 2018, 6, 171. [Google Scholar] [CrossRef]
- D’Eon, R.G.; Delparte, D. Effects of radio-collar position and orientation on GPS radio-collar performance, and the implications of PDOP in data screening. J. Appl. Ecol. 2005, 42, 383–388. [Google Scholar] [CrossRef]
- Brink, H.; Smith, R.J.; Skinner, K. Methods for lion monitoring: A comparison from the Selous Game Reserve, Tanzania. Afr. J. Ecol. 2013, 51, 366–375. [Google Scholar] [CrossRef]
- Caravaggi, A.; Banks, P.B.; Burton, A.C.; Finlay, C.M.V.; Haswell, P.M.; Hayward, M.W.; Rowcliffe, M.J.; Wood, M.D. A review of camera trapping for conservation behaviour research. Remote Sens. Ecol. Conserv. 2017, 3, 109–122. [Google Scholar] [CrossRef]
- Silveira, L.; Jácomo, A.T.; Diniz-Filho, J.A.F. Camera trap, line transect census and track surveys: A comparative evaluation. Biol. Conserv. 2003, 114, 351–355. [Google Scholar] [CrossRef]
- Rovero, F.; Zimmermann, F. Camera Trapping for Wildlife Research; Pelagic Publishing: Exeter, UK, 2016. [Google Scholar]
- Azlan, J.M.; Sharma, D.S.K. The diversity and activity patterns of wild felids in a secondary forest in Peninsular Malaysia. Oryx 2006, 40, 36–41. [Google Scholar] [CrossRef]
- Johnson, A.; Vongkhamheng, C.; Hedemark, M.; Saithongdam, T. Effects of human? Carnivore conflict on tiger (Panthera tigris) and prey populations in Lao PDR. Anim. Conserv. 2006, 9, 421–430. [Google Scholar] [CrossRef]
- Vogt, K.; Zimmermann, F.; Kölliker, M.; Breitenmoser, U. Scent-marking behaviour and social dynamics in a wild population of Eurasian lynx Lynx lynx. Behav. Process. 2014, 106, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Brereton, J.E.; Tuke, J.; Fernandez, E.J. A simulated comparison of behavioural observation sampling methods. Sci. Rep. 2022, 12, 3096. [Google Scholar] [CrossRef] [PubMed]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef]
- Burger, A.L.; Fennessy, J.; Fennessy, S.; Dierkes, P.W. Nightly selection of resting sites and group behavior reveal antipredator strategies in giraffe. Ecol. Evol. 2020, 10, 2917–2927. [Google Scholar] [CrossRef]
- Gübert, J.; Hahn-Klimroth, M.; Dierkes, P.W. BOVIDS: A deep learning-based software package for pose estimation to evaluate nightly behavior and its application to common elands (Tragelaphus oryx) in zoos. Ecol. Evol. 2022, 12, e8701. [Google Scholar] [CrossRef]
- Marks, M.; Qiuhan, J.; Sturman, O.; Von Ziegler, L.; Kollmorgen, S.; von der Behrens, W.; Mante, V.; Bohacek, J.; Yanik, M.F. Deep-learning based identification, tracking, pose estimation, and behavior classification of interacting primates and mice in complex environments. Nat. Mach. Intell. 2022, 4, 331–340. [Google Scholar] [CrossRef]
- Fazio, J.M.; Barthel, T.; Freeman, E.W.; Garlick-Ott, K.; Scholle, A.; Brown, J.L. Utilizing camera traps, closed circuit cameras and behavior observation software to monitor activity budgets, habitat use, and social interactions of zoo-housed asian elephants (Elephus maximus). Animals 2020, 10, 2026. [Google Scholar] [CrossRef] [PubMed]
- European Association of Zoos and Aquaria. EAZA Standards for the Accommodation and Care of Animals in Zoos and Aquaria; EAZA: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Bauer, H.; Packer, C.; Funston, P.F.; Henschel, P.; Nowell, K. Panthera leo. 2016, p. e.T15951A115130419. Available online: https://doi.org/10.2305/IUCN.UK.2016-3.RLTS.T15951A107265605.en (accessed on 20 April 2022).
- Durant, S.; Mitchell, N.; Ipavec, A.; Groom, R. Acinonyx jubatus. 2015, p. e.T219A50649567. Available online: https://doi.org/10.2305/IUCN.UK.2015-4.RLTS.T219A50649567.en (accessed on 20 April 2022).
- Burger, A.L.; Hartig, J.; Dierkes, P.W. Shedding light into the dark: Age and light shape nocturnal activity and sleep behaviour of giraffe. Appl. Anim. Behav. Sci. 2020, 229, 105012. [Google Scholar] [CrossRef]
- Hill, S.P.; Broom, D.M. Measuring zoo animal welfare: Theory and practice. Zoo Biol. 2009, 28, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Watters, J.V.; Margulis, S.W.; Atsalis, S. Behavioral monitoring in zoos and aquariums: A tool for guiding husbandry and directing research. Zoo Biol. 2009, 28, 35–48. [Google Scholar] [CrossRef]
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Regaiolli, B.; Rizzo, A.; Ottolini, G.; Miletto Petrazzini, M.E.; Spiezio, C.; Agrillo, C. Motion Illusions as Environmental Enrichment for Zoo Animals: A Preliminary Investigation on Lions (Panthera leo). Front. Psychol. 2019, 10, 2220. [Google Scholar] [CrossRef] [PubMed]
- Bashaw, M.J.; Kelling, A.S.; Bloomsmith, M.A.; Maple, T.L. Environmental effects on the behavior of zoo-housed lions and tigers with a case study of the effects of a visual barrier on pacing. J. Appl. Anim. Welf. Sci. 2007, 10, 95–109. [Google Scholar] [CrossRef]
- Siahaan, D.A.S.; Berliani, K.; Hartanto, A.; Tanjung, H.M.M.; Nurbayti. Study on daily activity pattern of captive lion (Panthera leo) in Siantar Zoo, North Sumatra, Indonesia. J. Phys. Conf. Ser. 2020, 1462, 12049. [Google Scholar] [CrossRef]
- Fink, L.B.; Scarlata, C.D.; VanBeek, B.; Bodner, T.E.; Wielebnowski, N.C. Applying behavioral and physiological measures to assess the relative impact of the prolonged COVID-19 pandemic closure on two mammal species at the Oregon Zoo: Cheetah (A. jubatus) and giraffe (G. c. reticulata and G. c. tippelskirchii). Animals 2021, 11, 3526. [Google Scholar] [CrossRef]
- Quirke, T.; O’Riordan, R.M.; Zuur, A. Factors influencing the prevalence of stereotypical behaviour in captive cheetahs (Acinonyx jubatus). Appl. Anim. Behav. Sci. 2012, 142, 189–197. [Google Scholar] [CrossRef]
- Quirke, T.; O’Riordan, R.M. The effect of different types of enrichment on the behaviour of cheetahs (Acinonyx jubatus) in captivity. Appl. Anim. Behav. Sci. 2011, 133, 87–94. [Google Scholar] [CrossRef]
- Skibiel, A.L.; Trevino, H.S.; Naugher, K. Comparison of several types of enrichment for captive felids. Zoo Biol. 2007, 26, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Clubb, R.; Mason, G.J. Natural behavioural biology as a risk factor in carnivore welfare: How analysing species differences could help zoos improve enclosures. Appl. Anim. Behav. Sci. 2007, 102, 303–328. [Google Scholar] [CrossRef]
- Munita, C.; Tadich, T.A.; Briceño, C. Comparison of 2 behavioral sampling methods to establish a time budget in a captive female cheetah (Acinonyx jubatus). J. Vet. Behav. 2016, 13, 1–5. [Google Scholar] [CrossRef]
- Rose, P.E.; Riley, L.M. Conducting behavioural research in the zoo: A guide to ten important methods, concepts and theories. JZBG 2021, 2, 421–444. [Google Scholar] [CrossRef]
- Halle, S.; Stenseth, N. Activity Patterns in Small Mammals: An ecological Approach; Springer: Berlin/Heidelberg, Germany, 2000; ISBN 978-3-642-62128-4. [Google Scholar]
- Burger, A.L.; Hartig, J.; Dierkes, P.W. Biological and environmental factors as sources of variation in nocturnal behavior of giraffe. Zoo Biol. 2021, 40, 171–181. [Google Scholar] [CrossRef]
- Cavallini, P.; Dragoo, J.; Garshelis, D.; Gaubert, P.; Gilchrist, J.; Goodmann, S.; Holekamp, K. Carnivores; Lynx: Barcelona, Spain, 2009. [Google Scholar]
- Haas, S.K.; Hayssen, V.; Krausman, P.R. Panthera leo. Mamm. Species 2005, 762, 1–11. [Google Scholar] [CrossRef]
- Hayward, M.W.; Hayward, G.J. Activity patterns of reintroduced lion Panthera leo and spotted hyaena Crocuta crocuta in the Addo Elephant National Park, South Africa. Afr. J. Ecol. 2006, 45, 135–141. [Google Scholar] [CrossRef]
- Kohari, D.; Sunada, A.; Matsui, Y.; Ootaki, A.; Hori, H. Behavioral restriction effects on activity motivation of a captive lion (Panthera leo persica). J. Vet. Behav. 2017, 17, 14–18. [Google Scholar] [CrossRef]
- Suraci, J.P.; Frank, L.G.; Oriol-Cotterill, A.; Ekwanga, S.; Williams, T.M.; Wilmers, C.C. Behavior-specific habitat selection by African lions may promote their persistence in a human-dominated landscape. Ecology 2019, 100, e02644. [Google Scholar] [CrossRef]
- Bashaw, M.J.; Bloomsmith, M.A.; Marr, M.J.; Maple, T.L. To hunt or not to hunt? A feeding enrichment experiment with captive large felids. Zoo Biol. 2003, 22, 189–198. [Google Scholar] [CrossRef]
- Altman, J.D.; Gross, K.L.; Lowry, S.R. Nutritional and behavioral effects of gorge and fast feeding in captive lions. J. Appl. Anim. Welf. Sci. 2005, 8, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Bissett, C.; Bernard, R.T.F. Habitat selection and feeding ecology of the cheetah (Acinonyx jubatus) in thicket vegetation: Is the cheetah a savanna specialist? J. Zool. 2007, 271, 310–317. [Google Scholar] [CrossRef]
- Wilson, A.M.; Lowe, J.C.; Roskilly, K.; Hudson, P.E.; Golabek, K.A.; McNutt, J.W. Locomotion dynamics of hunting in wild cheetahs. Nature 2013, 498, 185–189. [Google Scholar] [CrossRef]
- Funston, P.J.; Mills, M.G.L.; Biggs, H.C. Factors affecting the hunting success of male and female lions in the Kruger National Park. J. Zool. 2001, 253, 419–431. [Google Scholar] [CrossRef]
- Rigoudy, N.L.A.; Clinchy, M.; Peel, M.J.S.; Huebner, S.; Packer, C.; Zanette, L.Y. Hierarchy of fear: Experimentally testing ungulate reactions to lion, African wild dog and cheetah. Behav. Ecol. 2022, 33, 789–797. [Google Scholar] [CrossRef]
- Wilson, A.M.; Hubel, T.Y.; Wilshin, S.D.; Lowe, J.C.; Lorenc, M.; Dewhirst, O.P.; Bartlam-Brooks, H.L.A.; Diack, R.; Bennitt, E.; Golabek, K.A.; et al. Biomechanics of predator–prey arms race in lion, zebra, cheetah and impala. Nature 2018, 554, 183–188. [Google Scholar] [CrossRef]
- Hilborn, A.; Pettorelli, N.; Caro, T.; Kelly, M.J.; Laurenson, M.K.; Durant, S.M. Cheetahs modify their prey handling behavior depending on risks from top predators. Behav. Ecol. Sociobiol. 2018, 72, 74. [Google Scholar] [CrossRef]
- McPhee, M.E. Intact carcasses as enrichment for large felids: Effects on on- and off-exhibit behaviors. Zoo Biol. 2002, 21, 37–47. [Google Scholar] [CrossRef]
- Sengenberger, K.; Bus, H.; Versteege, L. EAZA-Best practice guidelines cheetah (Acinonyx jubatus). Recuper. El 2018, 1, 28–29. [Google Scholar]
- Kiffner, C.; Kioko, J.; Leweri, C.; Krause, S. Seasonal patterns of mixed species groups in large East African mammals. PLoS ONE 2014, 9, e113446. [Google Scholar] [CrossRef]
- Young, T.; Finegan, E.; Brown, R.D. Effects of summer microclimates on behavior of lions and tigers in zoos. Int. J. Biometeorol. 2013, 57, 381–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behaviour State | Definition | Behaviour Image |
|---|---|---|
| Active | ||
| Feeding/Drinking | Cat searches for food, ingests food by means of chewing or ingests water. |  |
| Locomotion | Walking, running or jumping with a forward locomotion. |  |
| Standing | Cat is in an upright position, immobile, with all four paws on the ground and legs extended, supporting the body. |  |
| Sitting | Body is in an upright position, with the hind legs flexed and touching the ground, while front legs are extended and straight. |  |
| Rest | ||
| Lying | Cat’s body is on the ground in a horizontal position, including on its side, back, belly or curled in a circular formation. |  |
| Sleeping | Cat is lying on the ground with its head down, performing minimal head or leg movement and is not easily disturbed. |  |
| Out | The animal is not visible on records |  |
| Lion (n = 6) | Cheetah (n = 5) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Behaviour State | Day (%) | Night (%) | p-Value | T | df | Day (%) | Night (%) | p-Value | T | df |
| Active | 26.8 ± 7.4 | 4.1 ± 3.4 | <0.001 | 22.29 | 26 | 51.2 ± 11.5 | 6.3 ± 6.2 | <0.001 | 38.38 | 26 |
| Locomote | 18.4 ± 4.8 | 3.1 ± 3.2 | <0.001 | 18.11 | 26 | 43.6 ± 10.7 | 5.4 ± 5.7 | <0.001 | 28.62 | 26 |
| Standing | 4.9 ± 2.1 | 0.8 ± 0.6 | <0.001 | 14.38 | 26 | 1.9 ± 0.7 | 0.2 ± 0.1 | <0.001 | 9.27 | 26 |
| Feeding/Drinking | 2.4 ± 1.9 | 0.2 ± 0.4 | <0.001 | 4.78 | 26 | 3.1 ± 1.2 | 0.0 ± 0.0 | <0.001 | 14.05 | 26 |
| Sitting | 1.1 ± 0.6 | 0.0 ± 0.0 | <0.001 | 5.65 | 26 | 2.6 ± 1.3 | 0.7 ± 0.7 | <0.001 | 7.92 | 26 |
| Rest | 72.8 ± 7.4 | 95.8 ± 3.4 | <0.001 | −22.29 | 26 | 48.2 ± 11.5 | 93.4 ± 6.2 | <0.001 | −38.24 | 26 |
| Lying | 44.6 ± 10.7 | 7.9 ± 2.3 | <0.001 | 32.17 | 26 | 31.0 ± 10.2 | 18.3 ± 5.3 | <0.001 | 10.37 | 26 |
| Sleeping | 28.2 ± 5.2 | 87.9 ± 4.7 | <0.001 | −43.30 | 26 | 17.2 ± 6.8 | 75.1 ± 6.9 | <0.001 | −50.55 | 26 |
| Out | 0.4 ± 0.4 | 0.1 ± 0.2 | n.s. | 3.16 | 26 | 0.6 ± 1.3 | 0.3 ± 0.5 | n.s. | 1.03 | 26 |
| Cheetah | Lion | p-Value | T | df | ||
|---|---|---|---|---|---|---|
| Total sleep time [hr] | Day | 2.05 ± 0.23 | 3.38 ± 0.43 | <0.001 | 7.481 | 27 |
| Night | 9.01 ± 0.65 | 10.57 ± 1.5 | <0.001 | 12.27 | 27 | |
| Sleep behaviour [%] | Day | 17.1 ± 6.8 | 28.2 ± 5.2 | <0.001 | 16.34 | 27 |
| Night | 75.1 ± 6.9 | 87.9 ± 4.7 | <0.001 | 59.04 | 27 | |
| Duration per sleep event [min] | Day | 11.0 ± 13.8 | 18.6 ± 25.4 | <0.001 | 14.79 | 27 |
| Night | 28.6 ± 38.9 | 78.9 ± 87.7 | <0.001 | 9.79 | 27 | |
| Sleep events (no.) | Day | 11.2 ± 5.3 | 10.9 ± 4.6 | <0.001 | 22.60 | 27 |
| Night | 18.9 ± 5.7 | 8.0 ± 4.4 | <0.001 | 10.76 | 27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seyrling, I.; Dierkes, P.W.; Burger, A.L. Diurnal and Nocturnal Behaviour of Cheetahs (Acinonyx jubatus) and Lions (Panthera leo) in Zoos. Animals 2022, 12, 2367. https://doi.org/10.3390/ani12182367
Seyrling I, Dierkes PW, Burger AL. Diurnal and Nocturnal Behaviour of Cheetahs (Acinonyx jubatus) and Lions (Panthera leo) in Zoos. Animals. 2022; 12(18):2367. https://doi.org/10.3390/ani12182367
Chicago/Turabian StyleSeyrling, Isabel, Paul Wilhelm Dierkes, and Anna Lena Burger. 2022. "Diurnal and Nocturnal Behaviour of Cheetahs (Acinonyx jubatus) and Lions (Panthera leo) in Zoos" Animals 12, no. 18: 2367. https://doi.org/10.3390/ani12182367
APA StyleSeyrling, I., Dierkes, P. W., & Burger, A. L. (2022). Diurnal and Nocturnal Behaviour of Cheetahs (Acinonyx jubatus) and Lions (Panthera leo) in Zoos. Animals, 12(18), 2367. https://doi.org/10.3390/ani12182367