
Feeding Dairy Ewes with Fresh or Dehydrated Sulla (Sulla coronarium L.) Forage. 2. Effects on Cheese Enrichment in Bioactive Molecules
 ,
,  , ,
, ,  , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, and Feeding Treatments
2.2. Milk Sampling and Cheese Manufacturing
2.3. Milk and Cheeses Analyses
2.3.1. Milk Physico-Chemical Traits
2.3.2. Cheese Physical Traits
2.3.3. Cheese Chemical Composition
2.3.4. Cheese Antioxidant Properties
2.3.5. Cheese Oxidative Stability
2.3.6. Cheese Fatty Acid Profile
2.4. Statistical Analysis
3. Results and Discussion
3.1. Milk Physico-Chemical Traits
3.2. Cheese Physico-Chemical Traits
3.3. Cheese Fatty Acid Profile
3.4. Cheese Biomarkers of Animals’ Feeding Regime
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Makkar, H.P.S. Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effect of feeding tannin rich feeds. Small Rumin. Res. 2003, 49, 241–256. [Google Scholar] [CrossRef]
- Vasta, V.; Luciano, G. The effects of dietary consumption of plants secondary compounds on small ruminants’ products quality. Small Rumin. Res. 2011, 101, 150–159. [Google Scholar] [CrossRef]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef] [PubMed]
- Soldado, D.; Bessa, R.J.; Jerónimo, E. Condensed Tannins as Antioxidants in Ruminants -Effectiveness and Action Mechanisms to Improve Animal Antioxidant Status and Oxidative Stability of Products. Animals 2021, 11, 3243. [Google Scholar] [CrossRef]
- Piluzza, G.; Sulas, L.; Bullitta, S. Tannins in forage plants and their role in animal husbandry and environmental sustainability: A review. Grass Forage Sci. 2014, 69, 32–48. [Google Scholar] [CrossRef]
- Min, B.R.; Barry, T.N.; Attwood, G.T.; McNabb, W.C. The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: A review. Anim. Feed Sci. Technol. 2003, 106, 3–19. [Google Scholar] [CrossRef]
- Molle, G.; Decandia, M.; Giovannetti, V.; Cabiddu, A.; Fois, N.; Sitzia, M. Responses to condensed tannins of flowering sulla (Hedysarum coronarium L.) grazed by dairy sheep. Part 1: Effects on feeding behaviour, intake, diet digestibility and performance. Livest. Sci. 2009, 123, 138–146. [Google Scholar] [CrossRef]
- Frutos, P.; Hervás, G.; Natalello, A.; Luciano, G.; Fondevila, M.; Priolo, A.; Toral, P.G. Ability of tannins to modulate ruminal lipid metabolism and milk and meat fatty acid profiles. Anim. Feed Sci. Technol. 2020, 269, 114623. [Google Scholar] [CrossRef]
- Di Trana, A.; Bonanno, A.; Cecchini, S.; Giorgio, D.; Di Grigoli, A.; Claps, S. Effects of Sulla forage (Sulla coronarium L.) on the oxidative status and milk polyphenol content in goats. J. Dairy Sci. 2015, 98, 37–46. [Google Scholar] [CrossRef]
- Giorgio, D.; Di Trana, A.; Di Gregorio, P.; Rando, A.; Avondo, M.; Bonanno, A.; Valenti, B.; Di Grigoli, A. Oxidative status of goats with different CSN1S1 genotypes fed ad libitum with fresh and dry forages. Antioxidants 2020, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- Ruisi, P.; Siragusa, M.; Di Giorgio, G.; Graziano, D.; Amato, G.; Carimi, F.; Giambalvo, D. Pheno-morphological agronomic and genetic diversity among natural populations of sulla (Hedysarum coronarium L.) collected in Sicily, Italy. Gen. Res. Crop. Evol. 2011, 58, 245–257. [Google Scholar] [CrossRef]
- Piluzza, G.; Bullitta, S. The dynamics of phenolic concentration in some pasture species and implications for animal husbandry. J. Sci. Food Agric. 2010, 90, 1452–1459. [Google Scholar] [CrossRef]
- Tava, A.; Biazzi, E.; Ronga, D.; Mella, M.; Doria, F.; D’Addabbo, T.; Candido, V.; Avato, P. Chemical identification of specialized metabolites from sulla (Hedysarum coronarium L.) collected in southern Italy. Molecules 2021, 26, 4606. [Google Scholar] [CrossRef]
- Barry, T.N.; McNabb, W.C. The implications of condensed tannins on the nutritive value of temperate forages fed to ruminants. Br. J. Nutr. 1999, 81, 263–272. [Google Scholar] [CrossRef]
- Amato, G.; Di Miceli, G.; Giambalvo, D.; Scarpello, C.; Stringi, L. Condensed Tannins Content in Sulla (Hedysarum cor-onarium L.) as Affected by Environment, Genotype and Growth Stage. In Bioactive Compounds in Pasture Species for Phytotherapy and Animal Welfare; Bullitta, S., Ed.; CNR-ISPAAM: Sassari, Italy, 2005; pp. 41–54. [Google Scholar]
- Bonanno, A.; Di Grigoli, A.; Di Trana, A.; Di Gregorio, P.; Tornambè, G.; Bellina, V.; Claps, S.; Maggio, G.; Todaro, M. Influence of fresh forage-based diets and αS1-casein (CSN1S1) genotype on nutrient intake and productive, metabolic, and hormonal responses in milking goats. J. Dairy Sci. 2013, 96, 2107–2117. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Montalbano, M.; Bellina, V.; Mazza, F.; Todaro, M. Effects of diet on casein and fatty acid profiles of milk from goats differing in genotype for αS1-casein synthesis. Eur. Food Res. Technol. 2013, 237, 951–963. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Mazza, F.; De Pasquale, C.; Giosuè, C.; Vitale, F.; Alabiso, M. Effects of ewes grazing sulla or ryegrass pasture for different daily durations on forage intake, milk production and fatty acid composition of cheese. Animal 2016, 10, 2074–2082. [Google Scholar] [CrossRef]
- Cabiddu, A.; Molle, G.; Decandia, M.; Spada, S.; Fiori, M.; Piredda, G.; Addis, M. Responses to condensed tannins of flowering sulla (Hedysarum coronarium L.) grazed by dairy sheep. Part 2: Effects on milk fatty acid profile. Livest. Sci. 2009, 123, 230–240. [Google Scholar] [CrossRef]
- Priolo, A.; Bella, M.; Lanza, M.; Galofaro, V.; Biondi, L.; Barbagallo, D.; Ben Salem, P.; Pennisi, P. Carcass and meat quality of lambs fed fresh sulla (Hedysarum coronarium L.) with or without polyethylene glycol or concentrate. Small Rumin. Res. 2005, 59, 281–288. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Miceli, G.; Di Grigoli, A.; Frenda, A.S.; Tornambè, G.; Giambalvo, D.; Amato, G. Effects of feeding green forage of sulla (Hedysarum coronarium L.) on lamb growth, gastrointestinal nematode infection, and carcass and meat quality. Animal 2011, 5, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Gladine, C.; Rock, E.; Morand, C.; Cauchart, D.; Durand, D. Bioavailability and antioxidant capacity of plant extracts rich in polyphenols, given as a single acute dose, in sheep made highly susceptible to lipoperoxidation. Br. J. Nutr. 2007, 98, 691–701. [Google Scholar] [CrossRef]
- Cabiddu, A.; Delgadillo-Puga, C.; Decandia, M.; Molle, G. Extensive ruminant production systems and milk quality with emphasis on unsaturated fatty acids, volatile compounds, antioxidant protection degree and phenol content. Animals 2019, 9, 771. [Google Scholar] [CrossRef]
- Di Grigoli, A.; Ponte, M.; Bonanno, A.; Maniaci, G.; Alabiso, M. Effects of Grazing Season on Physico-Chemical Characteristics and Fatty Acids of Nutritional Interest of Caciocavallo Palermitano Cheese. Animals 2022, 12, 544. [Google Scholar] [CrossRef]
- Segato, S.; Galaverna, G.; Contiero, B.; Berzaghi, P.; Caligiani, A.; Marseglia, A.; Cozzi, G. Identification of lipid biomarkers to discriminate between the different production systems for Asiago PDO cheese. J. Agric. Food Chem. 2017, 65, 9887–9892. [Google Scholar] [CrossRef]
- Maniaci, G.; Di Grigoli, A.; Bonanno, A.; Giosuè, C.; Ilardi, V.; Alabiso, M. Fatty Acids as Biomarkers of the Production Season of Caciocavallo Palermitano Cheese. Animals 2021, 11, 2675. [Google Scholar] [CrossRef]
- Vicente, F.; Santiago, C.; Jimenez-Calderòn, J.D.; Martínez-Fernàndez, A. Capacity of milk composition to identify the feeding system used to feed dairy cows. J. Dairy Res. 2017, 84, 254–263. [Google Scholar] [CrossRef]
- Gannuscio, R.; Ponte, M.; Di Grigoli, A.; Maniaci, G.; Di Trana, A.; Bacchi, M.; Alabiso, M.; Bonanno, A.; Todaro, M. Feeding dairy ewes with fresh or dehydrated sulla (Sulla coronarium L.) forage. 1. Effects on feed utilization, milk production and oxidative status. Animals 2022, 12, 2317. [Google Scholar] [CrossRef]
- International Standard FIL-IDF 29; Determination of the Casein Content of Milk. International Dairy Federation: Brussels, Belgium, 1964.
- International Standard FIL-IDF 20B; Determination of Total Nitrogen in Milk. International Dairy Federation: Brussels, Belgium, 1993.
- CIE (Commission International de l’Eclairage). Colorimetry–Official Recommendations of the International Commission on Illumination. In CIE Publication No. 15.2; CIE Central Bureau: Vienna, Austria, 1986. [Google Scholar]
- Standard FIL-IDF 4A; Cheese and Processed Cheese Product. Determination of the Total Solids Content. International Dairy Federation: Brussels, Belgium, 1982.
- Standard FIL-IDF 5B; Cheese and Processed Cheese Product. Determination of Fat Content-Gravimetric Method (Reference Method). International Dairy Federation: Brussels, Belgium, 1986.
- Standard FIL-IDF 25; Determination of the Protein Content of Processed Cheese Products. International Dairy Federation: Brussels, Belgium, 1964.
- Standard FIL-IDF 27; Determination of the Ash Content of Processed Cheese Products. International Dairy Federation: Brussels, Belgium, 1964.
- Panfili, G.; Manzi, P.; Pizzoferrato, L. High-performance Liquid Chromatographic Method for the Simultaneous Determination of Tocopherols, Carotenes, and Retinol and its Geometric Isomers in Italian Cheese. Analyst 1994, 169, 1161–1165. [Google Scholar] [CrossRef]
- Manzi, P.; Panfili, G.; Pizzoferrato, L. Normal and Reversed- Phase HPLC for more complete evaluation of tocopherols, retinols, carotenes and sterols in dairy products. Chromatographia 1996, 43, 89–93. [Google Scholar] [CrossRef]
- Rashidinejad, A.E.; Birch, J.; Sun-Waterhouse, D.; Everett, D.W. Effects of catechin on the phenolic content and antioxidant properties of low-fat cheese. Int. J. Food Sci. Technol. 2013, 48, 2448–2455. [Google Scholar] [CrossRef]
- Porter, L.J.; Hrstick, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyniadin and delphinidin. Phytochemistry 1986, 25, 223–230. [Google Scholar] [CrossRef]
- Tava, A.; De Benedetto, M.G.; Tedesco, D.; Di Miceli, G.; Piluzza, G. Proanthocyanidins from Hedysarum, Lotus and Onobrychis spp. growing in Sardinia and Sicily and their antioxidant activity. In Proceedings of the 20th International. Grassland Congress “Grassland: A global resource”, Dublin, Ireland; Wageningen, The Netherlands, 26 June–1 July 2005; p. 271. [Google Scholar]
- ISO (International Organization for Standardization). Determination of Substances Characteristic of Green and Black Tea. Part 1: Content of Total Polyphenols in Tea. Colorimetric Method Using Folin-Ciocalteu Reagent; International Organization for Standardization: Geneva, Switzerland, 2005; Volume ISO 14502-1, p. 28. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Vitale, F.; Di Miceli, G.; Todaro, M.; Alabiso, M.; Gargano, M.L.; Venturella, G.; Anike, F.N.; Isikhuemhen, O.S. Effects of feeding diets supplemented with medicinal mushrooms myceliated grains on some production, health and oxidation traits of dairy ewes. Int. J. Med. Mushrooms 2019, 21, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Standard FIL-IDF 74A:1991; Determination of the Peroxide Value. International Dairy Federation: Brussels, Belgium, 1991.
- Tarladgis, B.G.; Watts, B.M.; Younathan, M.T.; Dugan, L., Jr. A distillation method for the quantitative determination of malonaldehyde in rancid foods. J. Am. Oil Chem. Soc. 1960, 37, 44–48. [Google Scholar] [CrossRef]
- Mele, M.; Contarini, G.; Cercaci, L.; Serra, A.; Buccioni, A.; Povolo, M.; Conte, G.; Funaro, A.; Banni, S.; Lercker, G.; et al. Enrichment of pecorino cheese with conjugated linoleic acid by feeding dairy ewes with extruded linseed: Effect on fatty acid and triglycerides composition and on oxidative stability. Int. Dairy J. 2011, 21, 365–372. [Google Scholar] [CrossRef]
- Lee, M.R.F.; Tweed, J.K.S. Isomerisation of cis-9 trans-11 conjugated linoleic acid (CLA) to trans-9 trans-11 CLA during acidic methylation can be avoided by a rapid base catalysed methylation of milk fat. J. Dairy Res. 2008, 75, 354–356. [Google Scholar] [CrossRef]
- Kramer, J.K.; Cruz-Hernandez, C.; Deng, Z.; Zhou, J.; Jahreis, G.; Dugan, M.E. Analysis of conjugated linoleic acid and trans 18:1 isomers in synthetic and animal products. Am. J. Clin. Nutr. 2004, 79, 32. [Google Scholar] [CrossRef]
- Luna, P.; de la Fuente, M.A.; Juárez, M. Conjugated linoleic acid in processed cheeses during the manufacturing stages. J. Agric. Food Chem. 2005, 53, 2690–2695. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Chen, S.; Bobe, G.; Zimmerman, S.; Hammond, E.G.; Luhman, C.M.; Boylston, T.D.; Freeman, A.E.; Beitz, D.C. Physical and sensory properties of dairy products from cows with various milk fatty acid compositions. J. Agric. Food Chem. 2004, 52, 3422–3428. [Google Scholar] [CrossRef]
- Santos-Silva, J.; Bessa, R.J.B.; Santos-Silva, F. Effect of genotype, feeding system and slaughter weight on the quality of light lambs. II. Fatty acid composition of meat. Livest. Prod. Sci. 2002, 77, 187–194. [Google Scholar] [CrossRef]
- Giorgio, D.; Di Trana, A.; Di Napoli, M.A.; Sepe, L.; Cecchini, S.; Rossi, R.; Claps, S. Comparison of cheeses from goats fed 7 forages based on a new health index. J. Dairy Sci. 2019, 102, 6790–6801. [Google Scholar] [CrossRef]
- SAS (Statistical Analysis Systems Institute). SAS/STAT Qualification Tools User’s Guide, version 9.2; SAS Institute Inc.: Cary, NC, USA, 2010. [Google Scholar]
- Cappio-Borlino, A.; Portolano, B.; Todaro, M.; Macciotta, N.P.P.; Giaccone, P.; Pulina, G. Lactation curves of Valle del Belice dairy ewes for yields of milk, fat, and protein estimated with test day models. J. Dairy Sci. 1997, 80, 3023–3029. [Google Scholar] [CrossRef]
- Olagaray, K.E.; Bradford, B.J. Plant flavonoids to improve productivity of ruminants—A review. Anim. Feed Sci. Technol. 2019, 251, 21–36. [Google Scholar] [CrossRef]
- Nozière, P.; Graulet, B.; Lucas, A.; Martin, B.; Grolier, P.; Doreau, M. Carotenoids for ruminants: From forages to dairy products. Anim. Feed Sci. Technol. 2006, 131, 418–450. [Google Scholar] [CrossRef]
- Chowdhury, R.; Warnakula, S.; Kunutsor, S.; Crowe, F.; Ward, H.A.; Johnson, L.; Franco, O.H.; Butterworth, A.S.; Forouhi, N.G.; Thompson, S.G.; et al. Association of dietary, circulating, and supplement fatty acids with coronary risk. A systematic review and meta-analysis. Ann. Intern. Med. 2014, 160, 398–406. [Google Scholar] [CrossRef]
- Dehghan, M.; Mente, A.; Zhang, X.; Swaminathan, S.; Li, W.; Mohan, V.; Iqbal, R.; Kumar, R.; Wentzel-Viljoen, E.; Rosengren, A.; et al. Associations of fats and carbohydrate intake with cardiovascular disease and mortality in 18 countries from five continents (PURE): A prospective cohort study. Lancet 2017, 390, 2050–2062. [Google Scholar] [CrossRef]
- Kang, Z.Q.; Yang, Y.; Xiao, B. Dietary saturated fat intake and risk of stroke: Systematic review and dose–response meta-analysis of prospective cohort studies. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 179–189. [Google Scholar] [CrossRef]
- Astrup, A.; Magkos, F.; Bier, D.M.; Brenna, J.T.; de Oliveira Otto, M.C.; Hill, J.O.; King, J.C.; Mente, A.; Ordovas, J.M.; Volek, J.S.; et al. Saturated fats and health: A reassessment and proposal for food-based recommendations: JACC state-of-the-art review. J. Am. Coll. Cardiol. 2020, 76, 844–857. [Google Scholar] [CrossRef]
- Vlaeminck, B.; Fievez, V.; Cabrita, A.R.J.; Fonseca, A.J.M.; Dewhurst, R.J. Factors affecting odd-and branched-chain fatty acids in milk: A review. Anim. Feed Sci. Technol. 2006, 131, 389–417. [Google Scholar] [CrossRef]
- Wongtangtintharn, S.; Oku, H.; Iwasaki, H.; Toda, T. Effect of branched-chain fatty acids on fatty acid biosynthesis of human breast cancer cells. J. Nutr. Sci. Vitamonol. 2004, 50, 137–143. [Google Scholar] [CrossRef]
- Yang, Y.; Shangpei, L.; Chen, H.; Huang, M.; Zheng, J. Induction of apoptotic cell death and in vivo growth inhibition of human cancer cells by a saturated branched chain fatty acid, 13-methyltetradecanoic acid. Cancer Res. 2000, 60, 505–509. [Google Scholar]
- Chilliard, Y.; Ferlay, A.; Mansbridge, R.M.; Doreau, M. Ruminant milk fat plasticity: Nutritional control of saturated, polyunsaturated, trans and conjugated fatty acids. Ann. Zootech. 2000, 49, 181–205. [Google Scholar] [CrossRef]
- Nitasha Thakur, N.; Raigond, P.; Singh, Y.; Mishra, T.; Singh, B.; Lal, M.K.; Dutt, S. Recent updates on bioaccessibility of phytonutrients. Trends Food Sci. Technol. 2020, 97, 366–380. [Google Scholar] [CrossRef]
- Parodi, P.W. Milk Fat Nutrition. In Dairy Fats and Related Products; Tamime, A.Y., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2009; pp. 28–51. [Google Scholar]
- French, P.C.; Stanton, C.; Lawless, F.; O’Riordan, G.; Monahan, F.J.; Caffrey, P.J. Fatty acid composition including conjugated linoleic acid, of intramuscular fat from steers offered grazed grass, grass silage or concentrate-based diets. J. Anim. Sci. 2000, 78, 2849–2855. [Google Scholar] [CrossRef]
- Hur, S.J.; Kim, H.S.; Bahk, Y.Y.; Park, Y. Overview of conjugated linoleic acid formation and accumulation in animal products. Livest. Sci. 2017, 195, 105–111. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Diet, Nutrition and the Prevention of Chronic Diseases: Report of a Joint WHO/FAO Expert Consultation; WHO Technical Report Series No. 916; WHO: Geneva, Switzerland, 2003. [Google Scholar]
- FAO/WHO (Food and Agriculture Organization/World Health Organization). Codex Alimentarius: Foods for Special Dietary Uses Including Foods for Infants and Children, 2nd ed.; FAO: Rome, Italy, 1994. [Google Scholar]
- Di Trana, A.; Di Rosa, A.R.; Addis, M.; Fiori, M.; Di Grigoli, A.; Morittu, V.M.; Spina, A.A.; Claps, S.; Chiofalo, V.; Licitra, G.; et al. The Quality of Five Natural, Historical Italian Cheeses Produced in Different Months: Gross Composition, Fat-Soluble Vitamins, Fatty Acids, Total Phenols, Antioxidant Capacity, and Health Index. Animals 2022, 12, 199. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Diet (D) | Parity (p) | Significance p < (1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SHL | DSF2 | FSF2 | FSF4 | FSFL | SEM | TL | FL | SEM | D | p | |
| Milk yield, g/day | 1649 b | 1822 a | 1669 b | 1663 b | 1845 a | 82.52 | 1954 | 1505 | 98.48 | 0.0055 | 0.0057 |
| Lactose, % | 4.55 | 4.51 | 4.55 | 4.53 | 4.58 | 0.052 | 4.59 | 4.50 | 0.048 | 0.8514 | 0.2020 |
| Fat, % | 5.12 | 5.15 | 5.02 | 4.96 | 4.63 | 0.21 | 4.90 | 5.05 | 0.17 | 0.3246 | 0.5318 |
| Protein, % | 4.96 | 5.05 | 4.91 | 5.09 | 5.28 | 0.12 | 4.84 | 5.27 | 0.12 | 0.0588 | 0.0229 |
| Casein, % | 3.79 b | 3.93 ab | 3.75 b | 3.87 ab | 4.03 a | 0.10 | 3.72 | 4.03 | 0.11 | 0.0384 | 0.0598 |
| Whey protein, % | 0.90 | 0.90 | 0.90 | 0.98 | 1.00 | 0.037 | 0.87 | 1.00 | 0.029 | 0.0916 | 0.0082 |
| Non-protein nitrogen (NPN), % | 0.042 | 0.036 | 0.040 | 0.040 | 0.042 | 0.002 | 0.041 | 0.039 | 0.001 | 0.1906 | 0.2858 |
| Urea, mg/dL | 27.05 | 28.50 | 33.98 | 30.15 | 27.77 | 1.99 | 31.56 | 27.42 | 1.61 | 0.0875 | 0.0882 |
| Somatic cells count, n × 1000/mL | 164.19 ab | 62.21 b | 107.37 b | 302.20 a | 93.18 b | 41.35 | 176.70 | 114.96 | 38.00 | 0.0032 | 0.2699 |
| Somatic cells count, log10 n/mL | 5.02 b | 4.95 b | 5.03 ab | 5.31 a | 4.86 b | 0.078 | 5.07 | 4.99 | 0.069 | 0.0054 | 0.4166 |
| Total bacterial count, ufc × 1000/mL | 321.31 | 179.72 | 286.62 | 187.31 | 480.47 | 94.91 | 282.25 | 299.92 | 68.01 | 0.2297 | 0.8585 |
| Total bacterial count, log10 ufc/mL | 5.34 | 5.25 | 5.29 | 5.16 | 5.35 | 0.11 | 5.27 | 5.28 | 0.085 | 0.7215 | 0.9071 |
| pH | 6.48 ab | 6.44 b | 6.46 ab | 6.52 ab | 6.55 a | 0.024 | 6.50 | 6.48 | 0.022 | 0.0136 | 0.4208 |
| Titratable acidity, °SH/50 mL | 4.98 | 5.26 | 5.27 | 5.08 | 5.10 | 0.18 | 4.92 | 5.36 | 0.14 | 0.7793 | 0.0445 |
| Coagulation time (r), min | 17.44 | 18.61 | 17.64 | 17.91 | 18.34 | 1.30 | 18.74 | 17.24 | 1.14 | 0.9048 | 0.3733 |
| Curd firming time (k20), min | 1.89 | 1.73 | 1.44 | 1.68 | 1.83 | 0.19 | 1.94 | 1.48 | 0.14 | 0.4498 | 0.0385 |
| Curd firmness (a30), mm | 51.91 | 53.73 | 56.26 | 52.61 | 52.14 | 3.26 | 50.15 | 56.51 | 2.62 | 0.8326 | 0.1114 |
| Curd firmness (a2r), mm | 57.87 c | 62.24 a | 61.51 a | 58.33 bc | 60.47 ab | 0.97 | 58.66 | 61.51 | 0.96 | 0.0199 | 0.0584 |
| Diet (D) | Parity (p) | Significance p < (1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SHL | DSF2 | FSF2 | FSF4 | FSFL | SEM | TL | FL | SEM | D | p | |
| Cheese weight at 48 h, g | 216.32 | 230.18 | 226.34 | 225.32 | 219.85 | 8.71 | 239.70 | 207.51 | 8.91 | 0.7327 | 0.0220 |
| Cheese yield at 48 h, g/100 g milk | 16.88 | 16.45 | 16.82 | 16.87 | 15.45 | 0.44 | 16.22 | 16.77 | 0.38 | 0.0522 | 0.3251 |
| Dry matter (DM), % | 54.00 | 53.70 | 51.85 | 51.16 | 54.72 | 1.45 | 52.40 | 53.77 | 1.19 | 0.2474 | 0.4294 |
| Fat, % DM | 47.82 | 48.22 | 47.10 | 47.91 | 47.12 | 1.05 | 48.06 | 47.21 | 0.95 | 0.8840 | 0.5366 |
| Protein, % DM | 43.53 b | 43.39 b | 44.75 ab | 44.66 ab | 46.21 a | 0.83 | 43.90 | 45.11 | 0.79 | 0.0488 | 0.2970 |
| Ash, % DM | 5.40 | 4.95 | 5.18 | 5.47 | 5.44 | 0.16 | 5.28 | 5.30 | 0.11 | 0.1587 | 0.8756 |
| Vitamin A, mg/kg DM | 3.98 c | 5.02 b | 4.44 bc | 4.99 b | 5.96 a | 0.33 | 5.14 | 4.62 | 0.39 | <0.0001 | 0.3510 |
| Vitamin E, mg/kg DM | 2.97 b | 5.56 a | 2.74 b | 5.26 a | 5.09 a | 0.54 | 4.61 | 4.04 | 0.60 | <0.0001 | 0.5000 |
| Condensed tannins, g DE/kg DM | 0.86 ab | 0.79 b | 0.87 ab | 0.96 a | 0.86 ab | 0.026 | 0.88 | 0.85 | 0.017 | 0.0012 | 0.2170 |
| Polyphenols, g GAE/kg DM | 6.39 b | 6.61 ab | 7.12 ab | 7.22 ab | 7.40 a | 0.25 | 6.94 | 6.96 | 0.14 | 0.0050 | 0.8985 |
| TEAC, mmol/kg DM | 30.27 c | 31.20 bc | 36.02 a | 38.62 a | 34.77 ab | 1.66 | 35.20 | 33.15 | 1.40 | 0.0001 | 0.3024 |
| Peroxide value, mEq O2/kg fat | 0.33 b | 0.62 a | 0.74 a | 0.66 a | 0.42 b | 0.077 | 0.49 | 0.62 | 0.084 | <0.0001 | 0.2854 |
| TBARS, mg MDA/kg DM | 0.36 a | 0.27 b | 0.25 b | 0.22 b | 0.23 b | 0.058 | 0.25 | 0.28 | 0.080 | <0.0001 | 0.7346 |
| Internal colour | |||||||||||
| Lightness, L* | 87.36 | 87.46 | 88.56 | 87.05 | 86.13 | 1.53 | 88.11 | 86.51 | 1.30 | 0.7493 | 0.3893 |
| Redness, a* | −3.20 ab | −3.82 bc | −2.93 a | −3.49 abc | −4.24 c | 0.26 | −3.44 | −3.63 | 0.26 | <0.0001 | 0.6199 |
| Yellowness, b* | 13.16 | 13.69 | 12.68 | 13.68 | 15.19 | 0.94 | 13.26 | 14.14 | 0.80 | 0.1905 | 0.4411 |
| External colour | |||||||||||
| Lightness, L* | 89.66 ab | 89.24 b | 91.32 a | 89.65 ab | 89.02 b | 0.59 | 90.50 | 89.06 | 0.67 | <0.0001 | 0.1339 |
| Redness, a* | −1.62 b | −2.50 c | −1.09 a | −2.21 c | −2.12 c | 0.20 | −1.66 | −2.15 | 0.24 | <0.0001 | 0.1476 |
| Yellowness, b* | 9.04 a | 10.30 a | 7.20 b | 9.71 a | 9.67 a | 0.57 | 8.43 | 9.93 | 0.64 | <0.0001 | 0.1092 |
| Hardness, N/mm2 | 0.74 b | 0.76 b | 0.81 ab | 0.77 ab | 0.90 a | 0.037 | 0.74 | 0.86 | 0.030 | 0.0097 | 0.0074 |
| Diet (D) | Parity (p) | Significance p < (1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SHL | DSF2 | FSF2 | FSF4 | FSFL | SEM | TL | FL | SEM | D | p | |
| C4:0 | 2.46 | 2.43 | 2.50 | 2.60 | 2.41 | 0.10 | 2.61 | 2.36 | 0.068 | 0.6780 | 0.0188 |
| C6:0 | 2.53 b | 2.91 a | 2.71 ab | 2.67 ab | 2.77 ab | 0.074 | 2.77 | 2.66 | 0.051 | 0.0267 | 0.1450 |
| C7:0 | 0.018 | 0.030 | 0.022 | 0.027 | 0.035 | 0.007 | 0.021 | 0.032 | 0.005 | 0.5113 | 0.1488 |
| C8:0 | 2.44 b | 3.06 a | 2.70 ab | 2.60 b | 2.83 ab | 0.10 | 2.71 | 2.74 | 0.085 | 0.0042 | 0.8252 |
| C9:0 | 0.041 | 0.057 | 0.054 | 0.052 | 0.073 | 0.010 | 0.046 | 0.064 | 0.007 | 0.2802 | 0.0875 |
| C10:0 | 6.79 c | 9.04 a | 7.88 abc | 7.31 bc | 8.20 ab | 0.39 | 7.59 | 8.10 | 0.34 | 0.0028 | 0.3073 |
| C11:0 | 0.32 | 0.39 | 0.35 | 0.34 | 0.28 | 0.028 | 0.33 | 0.34 | 0.023 | 0.1204 | 0.6614 |
| C12:0 | 3.82 b | 4.89 a | 4.35 ab | 4.08 ab | 4.61 ab | 0.24 | 4.20 | 4.51 | 0.22 | 0.0132 | 0.3440 |
| C13:0 iso | 0.034 a | 0.010 b | 0.019 ab | 0.013 b | 0.004 b | 0.005 | 0.016 | 0.016 | 0.003 | 0.0032 | 0.9009 |
| C13:0 anteiso | 0.049 | 0.052 | 0.050 | 0.051 | 0.039 | 0.005 | 0.046 | 0.050 | 0.004 | 0.1425 | 0.4999 |
| C13:0 | 0.13 | 0.16 | 0.16 | 0.14 | 0.16 | 0.016 | 0.14 | 0.16 | 0.013 | 0.7360 | 0.1992 |
| C14:0 iso | 0.16 | 0.15 | 0.14 | 0.16 | 0.12 | 0.012 | 0.14 | 0.15 | 0.009 | 0.1159 | 0.4176 |
| C14:0 | 11.08 b | 12.28 a | 11.35 ab | 11.04 b | 10.80 b | 0.26 | 11.48 | 11.14 | 0.22 | 0.0038 | 0.3108 |
| C15:0 iso | 0.32 a | 0.24 b | 0.26 b | 0.24 b | 0.17 c | 0.011 | 0.25 | 0.24 | 0.008 | <0.0001 | 0.7338 |
| C15:0 anteiso | 0.49 a | 0.39 bc | 0.47 ab | 0.46 ab | 0.35 c | 0.024 | 0.42 | 0.44 | 0.020 | 0.0004 | 0.5105 |
| C14:1 c9 | 0.23 | 0.22 | 0.21 | 0.21 | 0.17 | 0.020 | 0.21 | 0.21 | 0.019 | 0.1039 | 0.9120 |
| C15:0 | 1.01 | 1.07 | 1.06 | 1.05 | 0.95 | 0.047 | 1.01 | 1.05 | 0.035 | 0.4427 | 0.4117 |
| C16:0 iso | 0.35 | 0.35 | 0.34 | 0.34 | 0.32 | 0.015 | 0.33 | 0.35 | 0.013 | 0.5364 | 0.3126 |
| C16:0 | 28.45 a | 27.96 a | 27.71 a | 27.51 a | 25.01 b | 0.56 | 27.58 | 27.07 | 0.57 | <0.0001 | 0.5339 |
| C17:0 iso | 0.62 a | 0.47 b | 0.56 ab | 0.58 ab | 0.59 ab | 0.029 | 0.59 | 0.55 | 0.021 | 0.0189 | 0.2267 |
| C16:1 t9 | 0.041 b | 0.051 ab | 0.054 ab | 0.069 ab | 0.094 a | 0.012 | 0.076 | 0.047 | 0.011 | 0.0092 | 0.0721 |
| C17:0 anteiso | 0.23 c | 0.24 bc | 0.22 c | 0.27 ab | 0.31 a | 0.010 | 0.052 | 0.26 | 0.008 | <0.0001 | 0.7058 |
| C16:1 c9 | 1.31 a | 1.13 bc | 1.17 ab | 1.24 ab | 1.01 c | 0.050 | 1.12 | 1.22 | 0.053 | 0.0002 | 0.2130 |
| C17:0 | 0.71 | 0.71 | 0.70 | 0.71 | 0.66 | 0.018 | 0.68 | 0.72 | 0.014 | 0.2219 | 0.0630 |
| C17:1 c9 | 0.19 | 0.19 | 0.19 | 0.18 | 0.18 | 0.008 | 0.17 | 0.19 | 0.008 | 0.2774 | 0.0773 |
| Diet (D) | Parity (p) | Significance p< (1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SHL | DSF2 | FSF2 | FSF4 | FSFL | SEM | TL | FL | SEM | D | p | |
| C18:0 iso | 0.047 | 0.038 | 0.038 | 0.036 | 0.067 | 0.009 | 0.034 | 0.056 | 0.005 | 0.1162 | 0.0122 |
| C18:0 | 7.97 | 6.26 | 7.09 | 7.62 | 6.36 | 0.61 | 7.34 | 6.78 | 0.50 | 0.1576 | 0.4448 |
| C18:1 t11, TVA | 1.51 b | 1.29 b | 1.90 b | 1.95 b | 3.44 a | 0.28 | 1.88 | 2.16 | 0.18 | 0.0006 | 0.2867 |
| C18:1 c9 | 17.29 a | 13.20 bc | 15.00 ab | 15.28 ab | 12.70 c | 0.61 | 14.88 | 14.50 | 0.54 | 0.0006 | 0.6231 |
| C19:0 | 0.010 b | 0.014 b | 0.009 b | 0.028 ab | 0.094 a | 0.015 | 0.021 | 0.041 | 0.009 | 0.0051 | 0.1552 |
| C18:2 n 6, LA | 2.31 ab | 2.14 b | 2.34 ab | 2.50 ab | 2.69 a | 0.10 | 2.39 | 2.40 | 0.079 | 0.0087 | 0.9481 |
| C20:0 | 0.34 ab | 0.27 b | 0.29 b | 0.40 a | 0.40 a | 0.027 | 0.34 | 0.34 | 0.026 | 0.0012 | 0.8365 |
| C18:3 n-6, | 0.031 | 0.026 | 0.033 | 0.019 | 0.031 | 0.006 | 0.024 | 0.031 | 0.005 | 0.4680 | 0.3428 |
| C20:1 c9 | 0.060 a | 0.036 b | 0.059 a | 0.057 ab | 0.073 a | 0.005 | 0.055 | 0.059 | 0.003 | 0.0035 | 0.4238 |
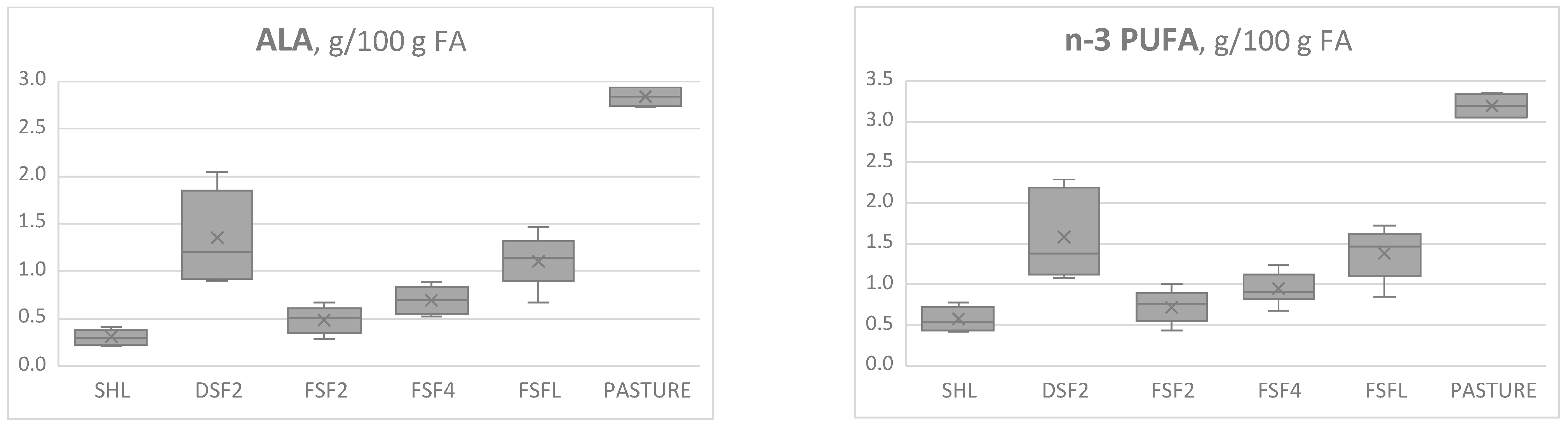
| C18:3 n-3, ALA | 0.30 c | 1.35 a | 0.57 b | 0.59 b | 1.15 a | 0.068 | 0.78 | 0.80 | 0.060 | <0.0001 | 0.8460 |
| C18:2 c9 t11 CLA, RA | 0.82 ab | 0.72 b | 0.93 ab | 0.92 ab | 1.12 a | 0.086 | 0.80 | 0.95 | 0.066 | 0.0329 | 0.7775 |
| C20:2 n-6 | 0.003 b | 0.005 b | 0.004 b | 0.003 b | 0.017 a | 0.004 | 0.003 | 0.009 | 0.002 | 0.0450 | 0.1153 |
| C22:0 | 0.15 ab | 0.13 b | 0.13 b | 0.17 a | 0.15 ab | 0.011 | 0.14 | 0.14 | 0.011 | 0.0063 | 0.9847 |
| C20:3 n-6 | 0.027 | 0.012 | 0.016 | 0.011 | 0.011 | 0.004 | 0.011 | 0.020 | 0.003 | 0.0564 | 0.0365 |
| C20:4 n-6, AA | 0.11 | 0.094 | 0.11 | 0.11 | 0.11 | 0.006 | 0.097 | 0.11 | 0.005 | 0.2775 | 0.0220 |
| C20:5 n-3, EPA | 0.10 | 0.11 | 0.11 | 0.11 | 0.13 | 0.006 | 0.11 | 0.12 | 0.005 | 0.1194 | 0.2291 |
| C22:5 n-3, DPA | 0.11 | 0.095 | 0.11 | 0.11 | 0.10 | 0.008 | 0.10 | 0.11 | 0.006 | 0.3268 | 0.3953 |
| C22:6 n-3, DHA | 0.012 | 0.006 | 0.015 | 0.013 | 0.030 | 0.008 | 0.016 | 0.014 | 0.006 | 0.3485 | 0.7662 |
| Other C16:1 cis | 0.020 b | 0.058 ab | 0.016 b | 0.038 ab | 0.072 a | 0.013 | 0.043 | 0.038 | 0.009 | 0.0130 | 0.7076 |
| Other C18:1 trans | 2.44 ab | 1.73 b | 2.46 ab | 2.49 ab | 3.61 a | 0.38 | 2.34 | 2.76 | 0.28 | 0.0411 | 0.3064 |
| Other C18:1 cis | 1.63 c | 2.11 abc | 1.97 bc | 2.25 ab | 2.70 a | 0.14 | 2.20 | 2.06 | 014 | 0.0001 | 0.4601 |
| Other C18:2 n-6 | 0.95 b | 1.24 b | 0.89 b | 1.22 b | 2.12 a | 0.17 | 1.14 | 1.43 | 0.11 | 0.0010 | 0.0791 |
| Other CLA isomers | 0.15 b | 0.21 ab | 0.16 b | 0.17 b | 0.29 a | 0.022 | 0.19 | 0.20 | 0.015 | 0.0014 | 0.7156 |
| Total CLA isomers | 0.97 b | 0.94 b | 1.09 ab | 1.09 ab | 1.40 a | 0.10 | 1.08 | 1.12 | 0.074 | 0.0274 | 0.7463 |
| Diet (D) | Parity (p) | Significance p < (1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SHL | DSF2 | FSF2 | FSF4 | FSFL | SEM | TL | FL | SEM | D | p | |
| Total FA, % DM | 43.59 | 44.96 | 44.15 | 44.90 | 45.23 | 0.79 | 44.96 | 44.17 | 0.78 | 0.2132 | 0.4838 |
| Non identified FA | 0.13 b | 0.33 a | 0.14 b | 0.19 ab | 0.30 ab | 0.050 | 0.17 | 0.26 | 0.032 | 0.0382 | 0.0601 |
| Branched chain FA | 2.31 a | 1.94 b | 2.10 ab | 2.16 ab | 1.97 b | 0.064 | 2.08 | 2.12 | 0.054 | 0.0014 | 0.6150 |
| Saturated FA, SFA | 70.50 b | 73.73 a | 71.15 ab | 70.54 bc | 67.69 c | 0.87 | 71.08 | 70.36 | 0.77 | 0.0016 | 0.5164 |
| Monounsaturated FA | 24.52 a | 19.99 b | 23.16 a | 23.67 a | 24.16 a | 0.63 | 22.98 | 23.21 | 0.60 | 0.0002 | 0.7876 |
| Polyunsaturated FA, PUFA | 4.93 c | 6.04 b | 5.24 bc | 5.85 bc | 7.74 a | 0.37 | 5.76 | 6.16 | 0.26 | 0.0005 | 0.2872 |
| Unsaturated FA, UFA | 29.34 ab | 25.96 c | 28.69 bc | 29.29 ab | 32.02 a | 0.84 | 28.75 | 29.38 | 0.77 | 0.0009 | 0.5700 |
| PUFA/SFA | 0.070 b | 0.082 b | 0.074 b | 0.084 b | 0.12 a | 0.006 | 0.082 | 0.089 | 0.005 | 0.0009 | 0.2640 |
| UFA/SFA | 0.42 ab | 0.35 c | 0.40 bc | 0.42 ab | 0.47 a | 0.018 | 0.41 | 0.42 | 0.015 | 0.0015 | 0.4860 |
| n-6 PUFA | 3.43 b | 3.54 b | 3.41 b | 3.84 b | 4.97 a | 0.25 | 3.67 | 4.01 | 0.17 | 0.0017 | 0.1791 |
| n-3 PUFA | 0.55 c | 1.57 a | 0.80 bc | 0.87 b | 1.40 a | 0.079 | 1.02 | 1.06 | 0.063 | <0.0001 | 0.6675 |
| n-6/n-3 | 6.37 a | 2.44 d | 4.99 b | 4.04 c | 3.74 c | 0.20 | 4.26 | 4.37 | 0.16 | <0.0001 | 0.6152 |
| LA/ALA | 7.94 a | 1.79 d | 5.12 b | 3.63 c | 2.55 cd | 0.25 | 4.16 | 4.25 | 0.17 | <0.0001 | 0.7314 |
| Thrombogenic index (TI) | 3.03 a | 2.79 a | 2.89 a | 2.87 a | 2.21 b | 0.11 | 2.81 | 2.70 | 0.11 | <0.0001 | 0.4851 |
| Health Promoting Index (HPI) | 0.37 ab | 0.31 c | 0.36 bc | 0.38 ab | 0.42 a | 0.015 | 0.36 | 0.38 | 0.014 | 0.0005 | 0.3409 |
| h/H | 0.51 a | 0.43 b | 0.47 ab | 0.49 a | 0.47 ab | 0.017 | 0.47 | 0.48 | 0018 | 0.0089 | 0.8557 |
| GHIC | 16.39 c | 25.58 b | 26.19 b | 29.27 b | 37.76 a | 1.84 | 26.92 | 27.15 | 1.74 | <0.0001 | 0.9277 |
| C14:1 c9 Δ-9 desaturase ratio | 0.020 a | 0.017 ab | 0.019 ab | 0.018 ab | 0.016 b | 0.001 | 0.018 | 0.018 | 0.001 | 0.0428 | 0.7585 |
| C16:1 c9 Δ-9 desaturase ratio | 0.044 a | 0.039 b | 0.041 ab | 0.043 a | 0.039 b | 0.002 | 0.039 | 0.043 | 0.002 | 0.0476 | 0.1495 |
| C18:1 c9 Δ-9 desaturase ratio | 0.68 | 0.68 | 0.69 | 0.67 | 0.68 | 0.017 | 0.67 | 0.69 | 0.015 | 0.9682 | 0.3418 |
| RA Δ-9 desaturase ratio | 0.35 a | 0.37 a | 0.33 a | 0.34 a | 0.25 b | 0.021 | 0.33 | 0.32 | 0.018 | 0.0025 | 0.8372 |
| X, Biomarker | Intercept | Slope | R2 | |||
|---|---|---|---|---|---|---|
| a | b | c | ||||
| linear | α-linolenic acid, ALA | g/100 FA | −0.91 | 38.45 | 0.8896 | |
| n-3 PUFA | g/100 FA | −8.33 | 35.96 | 0.8697 | ||
| C17:0 anteiso | g/100 FA | −88.07 | 445.75 | 0.7482 | ||
| LA/ALA | 79.57 | −10.57 | 0.7307 | |||
| PUFA | g/100 FA | −58.04 | 14.49 | 0.6895 | ||
| n-6/n-3 | 98.17 | −14.66 | 0.6679 | |||
| PUFA/SFA | −43.62 | 845.13 | 0.6098 | |||
| quadratic | α-linolenic acid, ALA | g/100 FA | −18.84 | 78.16 | −12.77 | 0.9435 |
| n-3 PUFA | g/100 FA | −28.63 | 68.61 | −8.85 | 0.9043 | |
| C17:0 anteiso | g/100 FA | 5.09 | −227.05 | 1158.35 | 0.7701 | |
| LA/ALA | 117.01 | −30.35 | 1.93 | 0.9213 | ||
| PUFA | g/100 FA | −120.74 | 33.32 | −1.31 | 0.7031 | |
| n-6/n-3 | 148.10 | −40.20 | 2.80 | 0.7773 | ||
| PUFA/SFA | −111.48 | 2251.82 | −6615.17 | 0.6382 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponte, M.; Maniaci, G.; Di Grigoli, A.; Gannuscio, R.; Ashkezary, M.R.; Addis, M.; Pipi, M.; Alabiso, M.; Todaro, M.; Bonanno, A. Feeding Dairy Ewes with Fresh or Dehydrated Sulla (Sulla coronarium L.) Forage. 2. Effects on Cheese Enrichment in Bioactive Molecules. Animals 2022, 12, 2462. https://doi.org/10.3390/ani12182462
Ponte M, Maniaci G, Di Grigoli A, Gannuscio R, Ashkezary MR, Addis M, Pipi M, Alabiso M, Todaro M, Bonanno A. Feeding Dairy Ewes with Fresh or Dehydrated Sulla (Sulla coronarium L.) Forage. 2. Effects on Cheese Enrichment in Bioactive Molecules. Animals. 2022; 12(18):2462. https://doi.org/10.3390/ani12182462
Chicago/Turabian StylePonte, Marialetizia, Giuseppe Maniaci, Antonino Di Grigoli, Riccardo Gannuscio, Mansour Rabie Ashkezary, Margherita Addis, Marianna Pipi, Marco Alabiso, Massimo Todaro, and Adriana Bonanno. 2022. "Feeding Dairy Ewes with Fresh or Dehydrated Sulla (Sulla coronarium L.) Forage. 2. Effects on Cheese Enrichment in Bioactive Molecules" Animals 12, no. 18: 2462. https://doi.org/10.3390/ani12182462
APA StylePonte, M., Maniaci, G., Di Grigoli, A., Gannuscio, R., Ashkezary, M. R., Addis, M., Pipi, M., Alabiso, M., Todaro, M., & Bonanno, A. (2022). Feeding Dairy Ewes with Fresh or Dehydrated Sulla (Sulla coronarium L.) Forage. 2. Effects on Cheese Enrichment in Bioactive Molecules. Animals, 12(18), 2462. https://doi.org/10.3390/ani12182462