
Relationship between Vegetation Habitats and Bird Communities in Urban Mountain Parks
 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Study Area and Methods
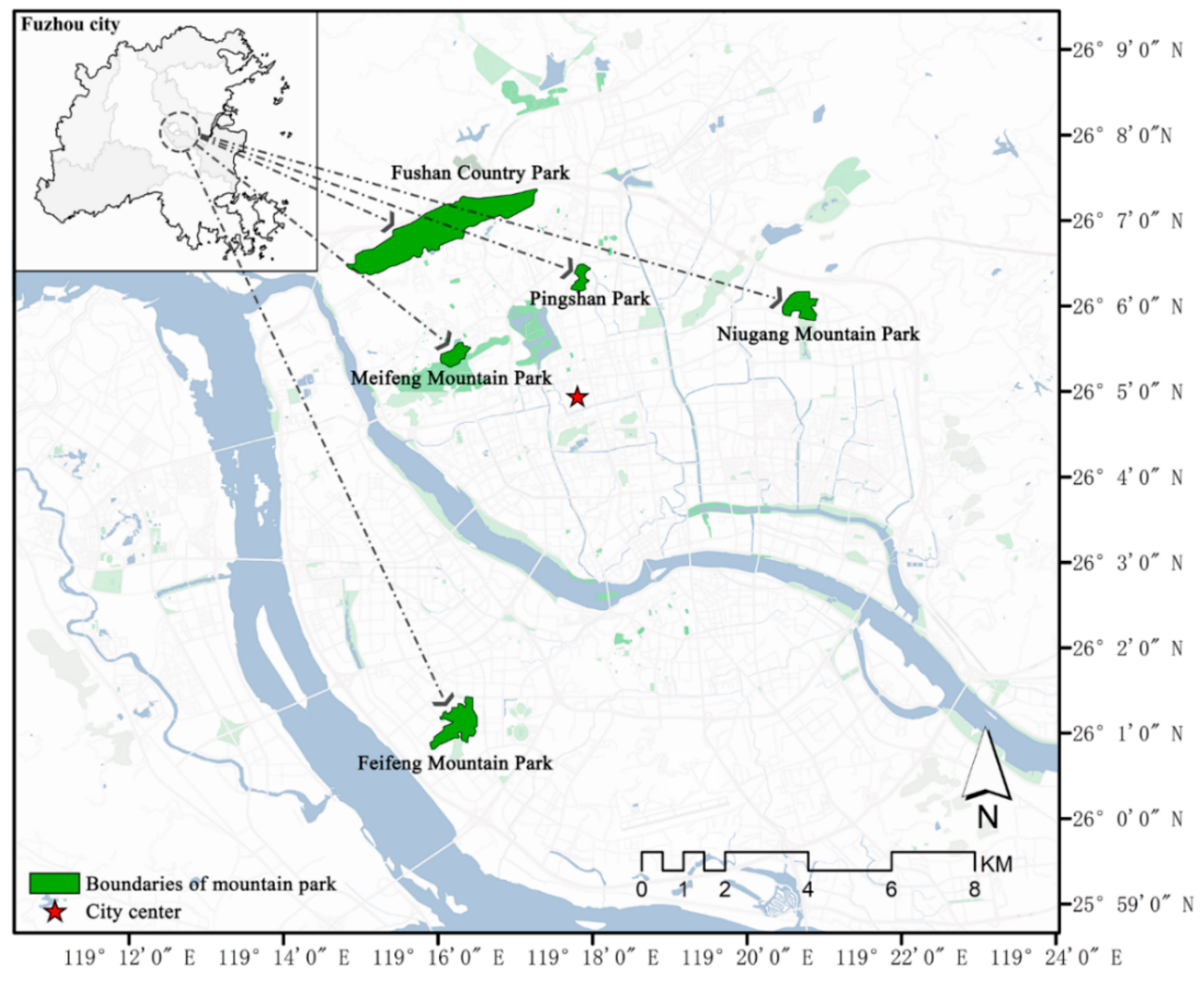
2.1. Study Area
2.2. Vegetation Habitats and Sampling Criteria
2.3. Vegetation Habitat Variables
- (1)
- Diversity of tree layer (DIVt), the diversity index of tree layer within the large sampling square, diversity index was calculated using the Shannon–Wiener index with the formula:H′ = −ΣPilnPiPi is the ratio of the number of individuals of plant species i to the total number of individuals in the community, i = 1, 2, …, S. S is the total number of plant species.
- (2)
- Diversity of shrub layer (DIVs), the diversity index of shrub layer plants within the medium sampling square, calculated by the same formula as the tree layer diversity index.
- (3)
- Richness of tree layer (RICt), the number of species in a tree layer within the large sampling square.
- (4)
- Richness of shrub layer (RICs), the number of species of shrub layer plants in the middle sampling square.
- (5)
- Richness of ground cover (RICg), the number of species of ground cover plants in the small sampling square.
- (6)
- Coverage of tree layer (COVt), the percentage of canopy area of a tree layer to the total area of a large sampling square.
- (7)
- Coverage of shrub layer (COVs), the percentage of shrub layer cover area to medium sampling square.
- (8)
- Coverage of ground cover (COVg), the percentage of ground cover area to a small sampling square.
- (9)
- Abundance of trees (ABUt), the number of individuals in the tree layer in the large sampling square.
- (10)
- Maximum tree height (MAX_HEIt), the maximum tree height of the tree layer in the large sampling square.
- (11)
- Average branch height under trees (AVE_HBt), the average branch height under trees in the tree layer within the large sampling square.
- (12)
- Average shrub heights (AVE_HEIs), the average height of shrub layer plants within the medium sampling square.
- (13)
- Average tree diameter at breast height (AVE_DBH), the average diameter at breast height of plants in the tree layer within the large sampling square.
- (14)
- Average tree height (AVE_HEIt), the average height of plants in the tree layer within the large sampling square.
- (15)
- Number of predators (NoP), the average number of predators (cats, dogs, and other animals that may prey on birds) within the transects during the survey period.
- (16)
- Number of tourists (NoT), the average number of tourists within the transects during the survey.
2.4. Bird Survey
- (1)
- Perching: birds were observed in resting, calling, preening, singing, etc.
- (2)
- Moving: birds were observed moving among different places, chasing, searching, fighting, etc.
- (3)
- Feeding: birds were observed pecking for food or eating worms, plants, or other food resources.
2.5. Analysis
2.5.1. Bird Diversity
2.5.2. Bird Behavior
2.5.3. Non-Metric Multidimensional Scaling
2.5.4. Response of Bird Communities to Different Vegetation Habitats
3. Results
3.1. Overview
3.2. Spatial and Temporal Variation of Bird Communities in Different Habitats
3.2.1. Spatial Variation of Bird Communities in Different Habitats
3.2.2. Temporal Variation of Bird Communities in Different Habitats
3.3. Relationship between Bird Diversity and Vegetation Habitats
4. Discussion
4.1. Distribution of Bird Communities in Different Vegetation Habitats
4.2. Response of Bird Communities to Vegetation Habitat Factors
4.3. Temporal and Spatial Changes in Bird Diversity
4.4. Implications for Urban Mountain Parks Management
5. Conclusions
- (1)
- We found more bird diversity and species in both sparse forest and dense forest habitats. However, fewer species were found in waterfront, shrub, and grassland habitats in different seasons. Moreover, Heyman (2010) [66] confirmed that the removal of 50% of shrubs did not affect local bird diversity, which is useful information for designing management plans for urban green spaces. Previous studies have confirmed that improving bird diversity throughout the city requires enhancing the quality of various types of habitats in urban green spaces, which, in turn, has a contributory effect on bird diversity enhancement.
- (2)
- Both the complexity and structure of vegetation have a significant effect on the bird community in urban mountain parks. Among them, the abundance and diversity of trees are critical vegetation variables affecting bird diversity, while the average branch height under trees has a significant effect on the Pielou index. The diversity and abundance of trees are integral parts of influencing the bird community in urban mountain parks, which facilitates the survival of birds in areas with high levels of human disturbance.
- (3)
- In all regression models, the number of tourists and predators found no significant effect on bird diversity in urban mountain parks. The reason may be that urban mountain parks have good natural vegetation resources, which are the main places for bird species to roost and shelter, and this also emphasizes the maintenance and protection of the existing green space resources in mountain parks. In addition, we hypothesize that bird communities may be greatly influenced by the degree of urbanization or disturbance of other environmental factors.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Savard, J.L.; Clergeau, P.; Mennechez, G. Biodiversity concepts and urban ecosystems. Landsc. Urban Plan. 2000, 48, 131–142. [Google Scholar] [CrossRef]
- Kowarik, I. Novel urban ecosystems, biodiversity, and conservation. Environ. Pollut. 2011, 159, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, H. World Urbanization: Trends and Prospects; Routledge: London, UK, 2017; pp. 43–64. [Google Scholar]
- Simberloff, D. The role of science in the preservation of forest biodiversity. For. Ecol. Manag. 1999, 115, 101–111. [Google Scholar] [CrossRef]
- Hedblom, M.; Murgui, E. Urban Bird Research in a Global Perspective; Springer: Cham, Switzerland, 2017; pp. 3–10. [Google Scholar]
- Wolch, J.R.; Byrne, J.; Newell, J.P. Urban green space, public health, and environmental justice: The challenge of making cities ‘just green enough’. Landsc. Urban Plan. 2014, 125, 234–244. [Google Scholar] [CrossRef]
- Li, F.; Liu, X.; Zhang, X.; Zhao, D.; Liu, H.; Zhou, C.; Wang, R. Urban ecological infrastructure: An integrated network for ecosystem services and sustainable urban systems. J. Clean. Prod. 2017, 163, S12–S18. [Google Scholar] [CrossRef]
- Koskimies, P. Birds as a tool in environmental monitoring. Ann. Zool. Fenn. 1989, 26, 153–166. [Google Scholar]
- Bonier, F.; Martin, P.R.; Wingfield, J.C. Urban birds have broader environmental tolerance. Biol. Lett. 2007, 3, 670–673. [Google Scholar] [CrossRef]
- Sulaiman, S.; Mohamad, N.H.N.; Idilfitri, S. Contribution of vegetation in urban parks as habitat for selective bird community. Procedia-Soc. Behav. Sci. 2013, 85, 267–281. [Google Scholar]
- Gray, E.R.; van Heezik, Y. Exotic trees can sustain native birds in urban woodlands. Urban Ecosyst. 2016, 19, 315–329. [Google Scholar] [CrossRef]
- Ciach, M.; Fröhlich, A. Habitat type, food resources, noise and light pollution explain the species composition, abundance and stability of a winter bird assemblage in an urban environment. Urban Ecosyst. 2017, 20, 547–559. [Google Scholar] [CrossRef]
- McKinney, M.L.; Kowarik, I.; Kendal, D. Wild urban ecosystems: Challenges and opportunities for urban development. Urban For. Urban Green. 2018, 29, 334–403. [Google Scholar] [CrossRef]
- Jokimäki, J.; Suhonen, J.; Kaisanlahti-Jokimäki, M. Urban core areas are important for species conservation: A European-level analysis of breeding bird species. Landsc. Urban Plan. 2018, 178, 73–81. [Google Scholar] [CrossRef]
- Chamberlain, D.; Kibuule, M.; Skeen, R.Q.; Pomeroy, D. Urban bird trends in a rapidly growing tropical city. Ostrich 2018, 89, 275–280. [Google Scholar] [CrossRef]
- Maseko, M.S.; Zungu, M.M.; Ehlers Smith, D.A.; Ehlers Smith, Y.C.; Downs, C.T. Effects of habitat-patch size and patch isolation on the diversity of forest birds in the urban-forest mosaic of Durban, South Africa. Urban Ecosyst. 2020, 23, 533–542. [Google Scholar] [CrossRef]
- Da Silva, B.F.; Pena, J.C.; Viana-Junior, A.B.; Vergne, M.; Pizo, M.A. Noise and tree species richness modulate the bird community inhabiting small public urban green spaces of a Neotropical city. Urban Ecosyst. 2021, 24, 71–81. [Google Scholar] [CrossRef]
- Rico-Silva, J.F.; Cruz-Trujillo, E.J.; Colorado, Z.G.J. Influence of environmental factors on bird diversity in greenspaces in an Amazonian city. Urban Ecosyst. 2021, 24, 365–374. [Google Scholar] [CrossRef]
- Huang, B.; Chiou, S.; Li, W. Landscape pattern and ecological network structure in urban green space planning: A case study of Fuzhou city. Land 2021, 10, 769. [Google Scholar] [CrossRef]
- Curzel, F.E.; Bellocq, M.I.; Leveau, L.M. Local and landscape features of wooded streets influenced bird taxonomic and functional diversity. Urban For. Urban Green. 2021, 66, 127369. [Google Scholar] [CrossRef]
- Paker, Y.; Yom-Tov, Y.; Alon-Mozes, T.; Barnea, A. The effect of plant richness and urban garden structure on bird species richness, diversity and community structure. Landsc. Urban Plan. 2014, 122, 186–195. [Google Scholar] [CrossRef]
- Xu, W.; Fu, W.; Dong, J.; Yu, J.; Huang, P.; Zheng, D.; Chen, Z.; Zhu, Z.; Ding, G. Bird Communities Vary under Different Urbanization Types—A Case Study in Mountain Parks of Fuzhou, China. Diversity 2022, 14, 555. [Google Scholar] [CrossRef]
- Chen, J.; van den Bosch, C.C.K.; Lin, C.; Liu, F.; Huang, Y.; Huang, Q.; Wang, M.; Zhou, Q.; Dong, J. Effects of personality, health and mood on satisfaction and quality perception of urban mountain parks. Urban For. Urban Green. 2021, 63, 127210. [Google Scholar] [CrossRef]
- Cheng, B.; Gou, Z.; Zhang, F.; Feng, Q.; Huang, Z. Thermal comfort in urban mountain parks in the hot summer and cold winter climate. Sustain. Cities Soc. 2019, 51, 101756. [Google Scholar] [CrossRef]
- Xu, W.; Zheng, D.; Huang, P.; Yu, J.; Chen, Z.; Zhu, Z.; Dong, J.; Fu, W. Does Bird Diversity Affect Public Mental Health in Urban Mountain Parks?—A Case Study in Fuzhou City, China. Int. J. Environ. Res. Public Health 2022, 19, 7029. [Google Scholar] [CrossRef] [PubMed]
- Fuzhou Landscape Bureau. Available online: http://ylj.fuzhou.gov.cn/ (accessed on 6 March 2021).
- Fuzhou Urban and Rural Planning Bureau. Available online: http://ghj.fuzhou.gov.cn (accessed on 5 March 2021).
- Ossola, A.; Hahs, A.K.; Nash, M.A.; Livesley, S.J. Habitat complexity enhances comminution and decomposition processes in urban ecosystems. Ecosystems 2016, 19, 927–941. [Google Scholar] [CrossRef]
- Yang, G.; Xu, J.; Wang, Y.; Wang, X.; Pei, E.; Yuan, X.; Li, H.; Ding, Y.; Wang, Z. Evaluation of microhabitats for wild birds in a Shanghai urban area park. Urban For Urban Green. 2015, 14, 246–254. [Google Scholar] [CrossRef]
- Yapp, W.B. The theory of line transects. Bird Study 1956, 3, 93–104. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burgess, N.D.; Hillis, D.M.; Hill, D.A.; Mustoe, S. Bird Census Techniques; Elsevier: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Madow, W.G.; Madow, L.H. On the theory of systematic sampling, I. Ann. Math. Stat. 1944, 15, 1–24. [Google Scholar] [CrossRef]
- Goodall, D.W. Some considerations in the use of point quadrats for the analysis of vegetation. Aust. J. Biol. Sci. 1952, 5, 1–41. [Google Scholar] [CrossRef]
- Fiala, A.C.; Garman, S.L.; Gray, A.N. Comparison of five canopy cover estimation techniques in the western Oregon Cascades. For. Ecol. Manag. 2006, 232, 188–197. [Google Scholar] [CrossRef]
- Cao, Z.; Dou, L.; Yang, N.; Zhang, K.; Wang, B.; Xu, Y.; Ran, J. Evaluating the effectiveness of vegetation conservation on a sacred mountain in western China. Nat. Conserv. 2021, 45, 1–9. [Google Scholar] [CrossRef]
- Shimelis, A.; Bekele, A.; Asefa, A.; Wilhams, A.; Gove, S.; Thirgood, S. Structuring of the birds of the Bale Mountains National Park. Walia 2011, 2011, 15–27. [Google Scholar]
- Meffert, P.J.; Dziock, F. The influence of urbanisation on diversity and trait composition of birds. Landsc. Ecol. 2013, 28, 943–957. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Williams, N.S.; Hahs, A.K.; Livesley, S.J. Approaches to urban vegetation management and the impacts on urban bird and bat assemblages. Landsc. Urban Plan. 2016, 153, 28–39. [Google Scholar] [CrossRef]
- MacKinnon, J.R.; MacKinnon, J.; Phillipps, K.; He, F. A Field Guide to the Birds of China; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Anderson, D.R. Guidelines for Line Transect Sampling of Biological Populations; The Unit: Logan, UT, USA, 1976. [Google Scholar]
- Collias, N.E.; Collias, E.C. Nest Building and Bird Behavior; Princeton University Press: Princeton, NJ, USA, 2014; Volume 857. [Google Scholar]
- Chao, A.; Chazdon, R.L.; Colwell, R.K.; Shen, T.J. A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol. Lett. 2005, 8, 148–159. [Google Scholar] [CrossRef]
- Heip, C. A new index measuring evenness. J. Mar. Biol. Assoc. UK 1974, 54, 555–557. [Google Scholar] [CrossRef]
- Pla, L. Bootstrap confidence intervals for the Shannon biodiversity index: A simulation study. J. Agric. Biol. Environ. Stat. 2004, 9, 42–56. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Community Ecology Package, R Package Version 2. 2013. Available online: http://sortie-admin.readyhosting.com/lme/R%20Packages/vegan.pdf (accessed on 10 October 2021).
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (H ill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Thode, H.C. Testing for Normality; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Gehan, E.A. A generalized Wilcoxon test for comparing arbitrarily singly-censored samples. Biometrika 1965, 52, 203–224. [Google Scholar] [CrossRef]
- McHugh, M.L. The chi-square test of independence. Biochem. Med. 2013, 23, 143–149. [Google Scholar] [CrossRef]
- Holland, S.M. Non-Metric Multidimensional Scaling (MDS). 2008, pp. 30602–32501. Available online: http://strata.uga.edu/8370/handouts/mdsTutorial.pdf (accessed on 10 October 2021).
- Abdi, H.; Williams, L.J. Principal component analysis. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Benesty, J.; Chen, J.; Huang, Y.; Cohen, I. Pearson Correlation Coefficient; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–4. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Lemon, S.C.; Roy, J.; Clark, M.A.; Friedmann, P.D.; Rakowski, W. Classification and regression tree analysis in public health: Methodological review and comparison with logistic regression. Ann. Behav. Med. 2003, 26, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Therneau, T.; Atkinson, B.; Ripley, B.; Ripley, M.B. Package ‘Rpart’. Available online: https://cran.r-project.org/web/packages/rpart/rpart.pdf (accessed on 20 April 2016).
- Germaine, S.S.; Rosenstock, S.S.; Schweinsburg, R.E.; Richardson, W.S. Relationships among breeding birds, habitat, and residential development in greater Tucson, Arizona. Ecol. Appl. 1998, 8, 680–691. [Google Scholar] [CrossRef]
- Fernandez-Juricic, E.; Jokimäki, J. A habitat island approach to conserving birds in urban landscapes: Case studies from southern and northern Europe. Biodivers. Conserv. 2001, 10, 2023–2043. [Google Scholar] [CrossRef]
- Erdelen, M. Bird communities and vegetation structure: I. Correlations and comparisons of simple and diversity indices. Oecologia 1984, 61, 277–284. [Google Scholar] [CrossRef]
- Amaya-Espinel, J.D.; Hostetler, M.E. The value of small forest fragments and urban tree canopy for Neotropical migrant birds during winter and migration seasons in Latin American countries: A systematic review. Landsc. Urban Plan. 2019, 190, 103592. [Google Scholar] [CrossRef]
- Fontana, S.; Sattler, T.; Bontadina, F.; Moretti, M. How to manage the urban green to improve bird diversity and community structure. Landsc. Urban Plan. 2011, 101, 278–285. [Google Scholar] [CrossRef]
- Bergner, A.; Avcı, M.; Eryiğit, H.; Jansson, N.; Niklasson, M.; Westerberg, L.; Milberg, P. Influences of forest type and habitat structure on bird assemblages of oak (Quercus spp.) and pine (Pinus spp.) stands in southwestern Turkey. For. Ecol. Manag. 2015, 336, 137–147. [Google Scholar] [CrossRef]
- Matsuba, M.; Nishijima, S.; Katoh, K. Effectiveness of corridor vegetation depends on urbanization tolerance of forest birds in central Tokyo, Japan. Urban For. Urban Green. 2016, 18, 173–181. [Google Scholar] [CrossRef]
- Prowse, T.A.; Collard, S.J.; Blackwood, A.; O’Connor, P.J.; Delean, S.; Barnes, M.; Cassey, P.; Possingham, H.P. Prescribed burning impacts avian diversity and disadvantages woodland-specialist birds unless long-unburnt habitat is retained. Biol. Conserv. 2017, 215, 268–276. [Google Scholar] [CrossRef]
- You, M.; Guan, C.; Lai, R. Spatial Structure of an Urban Park System Based on Fractal Theory: A Case Study of Fuzhou, China. Remote Sens. 2022, 14, 2144. [Google Scholar] [CrossRef]
- Bradfer Lawrence, T.; Gardner, N.; Dent, D.H. Canopy bird assemblages are less influenced by habitat age and isolation than understory bird assemblages in Neotropical secondary forest. Ecol. Evol. 2018, 8, 5586–5597. [Google Scholar] [CrossRef] [PubMed]
- Heyman, E. Clearance of understory in urban woodlands: Assessing impact on bird abundance and diversity. For. Ecol. Manag. 2010, 260, 125–131. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| DIVt | 0.490 | 0.108 | 0.085 | 0.123 |
| DIVs | 0.243 | −0.454 | 0.168 | −0.305 |
| RICt | 0.465 | 0.054 | 0.171 | 0.246 |
| RICs | 0.185 | −0.382 | −0.295 | −0.344 |
| RICg | −0.220 | −0.265 | 0.002 | 0.180 |
| COVt | 0.131 | 0.115 | −0.430 | 0.209 |
| COVs | 0.326 | −0.066 | −0.287 | −0.109 |
| COVg | 0.046 | 0.383 | 0.420 | −0.117 |
| ABUt | 0.398 | 0.008 | 0.179 | 0.371 |
| MAX_HEIt | 0.294 | 0.223 | −0.351 | −0.325 |
| AVE_HBt | 0.021 | 0.349 | −0.001 | −0.451 |
| AVE_HEIs | 0.099 | −0.420 | 0.059 | 0.154 |
| AVE_DBH | −0.047 | 0.216 | −0.450 | 0.323 |
| AVE_HEIt | −0.138 | −0.068 | −0.208 | 0.192 |
| Standard deviation | 1.834 | 1.453 | 1.321 | 1.239 |
| Proportion of Variance | 0.240 | 0.151 | 0.125 | 0.110 |
| Cumulative Proportion | 0.240 | 0.391 | 0.516 | 0.625 |
| NO. | Model | Intercept | df | Loglik | AICc | ΔAICc | wi |
|---|---|---|---|---|---|---|---|
| Richness (Chao1) | |||||||
| 1 | PC1 | 19.917 | 3 | −83.785 | 174.712 | 0.000 | 0.181 |
| 2 | PC1 + PC2 | 19.917 | 4 | −83.154 | 176.309 | 1.596 | 0.081 |
| 3 | NoT + PC1 | 21.353 | 4 | −83.579 | 177.157 | 2.445 | 0.053 |
| 4 | PC1 + PC4 | 19.917 | 4 | −83.743 | 177.487 | 2.774 | 0.045 |
| 5 | NoP + PC1 | 20.176 | 4 | −83.764 | 177.529 | 2.817 | 0.044 |
| Abundance | |||||||
| 4 | PC1 + PC4 | 75.160 | 4 | −115.353 | 240.706 | 0.000 | 0.144 |
| 5 | PC1 | 75.160 | 3 | −116.828 | 240.800 | 0.093 | 0.137 |
| 6 | PC1 + PC2 | 75.160 | 4 | −116.606 | 243.213 | 2.507 | 0.041 |
| 7 | NoT + PC1 + PC4 | 80.976 | 5 | −115.083 | 243.325 | 2.619 | 0.039 |
| 8 | NoT + PC1 | 79.901 | 4 | −116.669 | 243.338 | 2.632 | 0.039 |
| Shannon diversity | |||||||
| 9 | PC1 | 2.549 | 3 | −6.692 | 20.528 | 0.000 | 0.208 |
| 10 | NoT + PC1 | 2.649 | 4 | −6.208 | 22.416 | 1.888 | 0.081 |
| 11 | NoP + PC1 | 2.594 | 4 | −6.401 | 22.802 | 2.274 | 0.067 |
| 12 | PC1 + PC4 | 2.549 | 4 | −6.488 | 22.975 | 2.447 | 0.061 |
| 13 | PC1 + PC3 | 2.549 | 4 | −6.506 | 23.011 | 2.484 | 0.060 |
| Pielou index | |||||||
| 14 | PC4 | 0.619 | 3 | 46.284 | −85.425 | 0.020 | 0.065 |
| 15 | PC3 | 0.619 | 3 | 45.957 | −84.772 | 0.673 | 0.047 |
| 16 | PC3 + PC4 | 0.619 | 4 | 47.355 | −84.710 | 0.735 | 0.045 |
| 17 | PC1 | 0.619 | 3 | 45.839 | −84.535 | 0.910 | 0.041 |
| 18 | PC1 + PC4 | 0.619 | 4 | 47.223 | −84.446 | 0.999 | 0.040 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Yu, J.; Huang, P.; Zheng, D.; Lin, Y.; Huang, Z.; Zhao, Y.; Dong, J.; Zhu, Z.; Fu, W. Relationship between Vegetation Habitats and Bird Communities in Urban Mountain Parks. Animals 2022, 12, 2470. https://doi.org/10.3390/ani12182470
Xu W, Yu J, Huang P, Zheng D, Lin Y, Huang Z, Zhao Y, Dong J, Zhu Z, Fu W. Relationship between Vegetation Habitats and Bird Communities in Urban Mountain Parks. Animals. 2022; 12(18):2470. https://doi.org/10.3390/ani12182470
Chicago/Turabian StyleXu, Weizhen, Jiao Yu, Peilin Huang, Dulai Zheng, Yuxin Lin, Ziluo Huang, Yujie Zhao, Jiaying Dong, Zhipeng Zhu, and Weicong Fu. 2022. "Relationship between Vegetation Habitats and Bird Communities in Urban Mountain Parks" Animals 12, no. 18: 2470. https://doi.org/10.3390/ani12182470
APA StyleXu, W., Yu, J., Huang, P., Zheng, D., Lin, Y., Huang, Z., Zhao, Y., Dong, J., Zhu, Z., & Fu, W. (2022). Relationship between Vegetation Habitats and Bird Communities in Urban Mountain Parks. Animals, 12(18), 2470. https://doi.org/10.3390/ani12182470