Simple Summary
The origin and prevalence of Listeria monocytogenes was studied in dairy cattle farms in order to examine its diversity and determine its possible persistence in manure. The utilization of manure for agricultural purposes is common in many countries. While properly treated and managed manure is an effective and safe fertilizer, foodborne illness outbreaks can occur, as many of the most prominent foodborne pathogens are carried by healthy livestock. It is, therefore, necessary to study the origin and persistence of zoonotic agents in general and of L. monocytogenes in particular, in order to avoid recirculation in farms and reduce risk for human populations.
Abstract
Listeria monocytogenes is an opportunistic pathogen that is widely distributed in the environment. Here we show the prevalence and transmission of L. monocytogenes in dairy farms in the Cantabria region, on the northern coast of Spain. A total of 424 samples was collected from 14 dairy farms (5 organic and 9 conventional) and 211 L. monocytogenes isolates were recovered following conventional microbiological methods. There were no statistically significant differences in antimicrobial resistance ratios between organic and conventional farms. A clonal relationship among the isolates was assessed by pulsed field gel electrophoresis (PFGE) analysis and 64 different pulsotypes were obtained. Most isolates (89%, n = 187) were classified as PCR serogroup IVb by using a multiplex PCR assay. In this case, 45 isolates of PCR serogroup IVb were whole genome-sequenced to perform a further analysis at genomic level. In silico MLST analysis showed the presence of 12 sequence types (ST), of which ST1, ST54 and ST666 were the most common. Our data indicate that the environment of cattle farms retains a high incidence of L. monocytogenes, including subtypes involved in human listeriosis reports and outbreaks. This pathogen is shed in the feces and could easily colonize dairy products, as a result of fecal contamination. Effective herd and manure management are needed in order to prevent possible outbreaks.
1. Introduction
Listeria monocytogenes is a Gram-positive, non-spore-forming, motile, facultative anaerobic, rod-shaped bacterium that is an opportunistic pathogen. It causes listeriosis, a severe foodborne infection, leading to blood, brain or fetal infections in humans. Ruminants (cattle, sheep and goats), when infected, are mainly asymptomatic and excrete the bacteria in their feces, but infection can also lead to encephalitis or meningoencephalitis and abortion [1]. L. monocytogenes is resistant to cephalosporins, fosfomycin and fusidic acid [2]. Listeriosis is generally treated with ampicillin used alone or together with gentamicin [3]. Aminopenicillins can be substituted by cotrimoxazole, fluoroquinolones, rifampicin or linezolid. Vancomycin may occasionally be prescribed for non-meningeal infections, and erythromycin is used for listeriosis treatment during pregnancy [2]. Various recent reports have highlighted the elevation in the rate of resistance to one or more of the previously mentioned antibiotics, mainly in environmental and animal isolates and less frequently in clinical cases [4,5,6].
L. monocytogenes can be found in a variety of sources, including water, soil, food products, humans and animals [7,8]. At the farm level, the introduction sources of L. monocytogenes to livestock are not completely understood, but the route of L. monocytogenes infection in ruminants is considered to be contaminated silage, contaminated cattle bedding and contaminated water troughs [9]. The life cycle of this pathogen between the animals and their natural environment is fundamentally uninterrupted and hard to break [10]. As an environmental saprophyte, it is highly adapted to harsh conditions and competitive microbiota [11]. This is an important factor to take into consideration in silage production, as a pH below 6.0 should be achieved during silage fermentation in order to impede the growth of L. monocytogenes [12]. Many studies have reported the repeated detection of L. monocytogenes in samples collected from dairy farms [13,14,15]. The presence of L. monocytogenes in tank milk (TM) and tank milk filters (TMF) [16,17] was attributed to fecal contamination during the milking process and/or inefficient cleaning and sanitizing processes, possibly leading to biofilm formation in the milking equipment. Although on-farm eradication is highly unlikely, considering the ability of L. monocytogenes to survive and multiply in many habitats and hosts, the transmission and contamination rates could probably be decreased by implementing appropriate intervention strategies.
L. monocytogenes can be divided into four different phylogenetic lineages (I, II, III and IV), with each lineage containing different serotypes [18]. According to the somatic O antigen, it can be sub-classified in 14 serotypes. All serotypes could cause listeriosis, but serotypes 1/2a, 1/2b and 4b are more prevalent, causing over 95% of invasive infections. Serotype 4b isolates account for 50% of human listeriosis outbreaks [19] and are mostly prevalent in epidemic outbreaks. Serotypes 1/2a and 1/2b are mostly sporadic [20], with serotype 1/2a more adapted to environmental conditions [18]. Most studies suggest a higher prevalence of serotype 4b among cases of human listeriosis, but since this prevalence cannot be attributed to a higher prevalence of this serotype in food, it has been hypothesized that serotype 4b may have a higher pathogenicity compared to serotypes 1/2a and 1/2b [21].
The characterization and differentiation of foodborne pathogenic bacteria at the strain level have been achieved by the development of molecular methods. Macrorestriction with specific restriction enzymes combined with pulsed-field gel electrophoresis (PFGE) has been used since the 1990s as an effective tool for identifying bacterial subgroups. PFGE using restriction enzyme AscI and ApaI is well established as an effective molecular subtyping method for characterizing L. monocytogenes [22,23]. On the other hand, the multilocus sequence typing (MLST) method for analysis of L. monocytogenes isolates clonality is carried out by sequencing seven housekeeping genes: abcZ, bglA, cat, dapE, dat, ldh and lhkA [24], and has shown that L. monocytogenes forms a population structured by groups of genetically similar isolates, called clonal complexes (CCs) [24]. Currently, whole-genome sequencing (WGS) is the most powerful typing tool for population biology studies, including L. monocytogenes [25]. Core genome multilocus sequence typing (cgMLST, covering 1748 genes in L. monocytogenes) is a method with a high reproducibility rate that allows inter-laboratory strain comparison by using standardized allele and type nomenclatures [26,27].
The prevalence and diversity of L. monocytogenes isolated from dairy farms in the Cantabria region (north coast of Spain) are described in this study. Information on the genotypic characteristics is provided and possible public health implications are being discussed, bearing in mind that information on the molecular level of the foodborne bacteria can help in anticipating outbreaks and designing effective control strategies in the event of possible disease emergence. The PCR serogroups and pulsotypes of all isolates, as well as the WGS analysis and the antimicrobial susceptibility profiles of a selection of 45 isolates of PCR serogroup IVb were performed. According to the results of this study, the L. monocytogenes analyzed encode virulence factors associated with human disease, capable of surviving at least 35 days in livestock waste and 70 days on pasture. The information presented here helps to increase our understanding of the on-farm life cycle of L. monocytogenes.
2. Materials and Methods
2.1. Study Design
The sampling was performed during 2017—2019 and included 14 dairy cattle farms (5 organic and 9 conventional) from 8 different sites in the Cantabria region (north coast of Spain). Samples were taken from forage, water, raw tank milk, the tank milk filters, fresh feces, stored manure and soil. In total, 424 samples were collected and tested for L. monocytogenes.
2.2. Sample Collection, Processing and L. monocytogenes Isolation
The samples were collected using sterile material, transported to the laboratory on ice within 2 h from the collection and subjected to detection of L. monocytogenes immediately upon arrival using 10 g for solid samples; 3 L filtered through a 0.5 nm sterile filter for water; 100 mL centrifuged at 8500 rpm/10 min for milk. Rectal swabs and filters from the milk tank were placed directly in the selective media. Samples were treated according to the ISO11290-1:2017 method (https://www.iso.org/standard/60313.html, accessed on 1 January 2018) with modifications. Briefly, for an initial selective enrichment, each sample was inoculated in 100 mL half Fraser broth (HFB) (Oxoid, Basingstoke, UK) for 24 h at 30 °C. A second enrichment followed, by inoculation of 100 µL of the suspension of the first enrichment in 10 mL of Full-strength Fraser broth (FFB) (Oxoid, Basingstoke, UK) for 24 h at 37 °C. Next, 100 µL were spread onto agar Listeria according to Ottaviani and Agosti (ALOA) (bioMérieux, Marcy-l’Étoile, France) and modified Oxford agar (MOX) (Oxoid, Basingstoke, UK) and incubated for 24 h at 37 °C. Presumptive Listeria spp. colonies from each ALOA and MOX plates were re-streaked to tryptic soy agar supplemented with 0.6% yeast extract (TSA-YE) plates (bioMérieux, Marcy-l’Étoile, France), incubated for 24 h at 37 °C and subjected to microscopic examination, Gram staining, glucose fermentation, oxidase, catalase and API tests (bioMérieux, Marcy-l’Étoile, France), according to the manufacturer’s instructions.
2.3. Decline of L. monocytogenes in Manure
The experiment was performed as previously described [28] and included a tank with manure not inoculated with L. monocytogenes (used as negative control), a tank with manure inoculated with L. monocytogenes strain CIP103575 (https://www.pasteur.fr/en/cip-distribution, accessed on 1 January 2019; positive control) and samples inoculated with L. monocytogenes strain MS6507 isolated from cattle feces (this study), all repeated six times (n = 6). The L. monocytogenes strains used were grown at 37 °C in tryptic soy broth supplemented with 0.6% yeast extract (TSB-YE) (bioMérieux, Marcy-l’Étoile, France). Before manure inoculation, L. monocytogenes strains were centrifuged in order to remove growth media, washed and resuspended in 0.1% peptone buffer (Oxoid, Basingstoke, UK) to a density of 109 CFU/mL. Inoculants were providing 106 CFU/g of manure wet weight, a quantity that represents a very high pathogen load and represents the worst-case scenario in terms of pathogen load. The presence of L. monocytogenes was monitored by survival curves generated by plate counting in CHROMagarTM Listeria plates (https://www.chromagar.com/en/product/chromagar-listeria, accessed on 1 January 2019).
2.4. Decline of L. monocytogenes in Pasture Crops
The experiment was performed as previously described [28] using compost infected with L. monocytogenes MS6507. The infected compost was applied to crops and the experiment was performed in a germination chamber under controlled conditions of temperature and light [28]. The presence of L. monocytogenes on the surface of plants was monitored on CHROMagarTM Listeria plates for the generation of the survival curve and the calculation of CFU/g of plant, using 3 g of pasture crops as described in [28].
2.5. PCR Serogroup Determination
L. monocytogenes isolates were initially subtyped by using a multiplex PCR serotyping to differentiate the four major L. monocytogenes serotypes (1/2a, 1/2b, 1/2c and 4b) into distinct PCR serogroups (IIa, IIb, IIc and IVb) [29,30].
2.6. Pulse-Field Gel Electrophoresis (PFGE) Typing
The clonal groups were identified by pulsed-field agarose gel electrophoresis (PFGE). The PFGE typing results were analyzed following an optimized PulseNet standardized protocol [31], as previously described [32]. L. monocytogenes strain H2446 was used as reference [33]. DNA extraction was carried out on agarose plugs slices in a conventional manner. DNA digestion was performed with two different restriction enzymes, AscI and ApaI (New England BioLabs, Ibswich, MA, USA), and the generated restriction fragments were separated by electrophoresis in a clamped homogeneous electric field (CHEF-DRII) system (BIO-RAD Laboratories, Hercules, CA, USA), with the following running parameters: gradient of 6 V/cm, angle 120°, temperature of 14 °C, initial switch time 4 s, final switch time 40 s and run time 24 h. Using the Bionumerics software (Version 4.5, Applied Maths, Kortrijk, Belgium), a database of the ApaI and AscI patterns obtained using PFGE was constructed with all the isolates of L. monocytogenes found in the different surveys conducted. These ApaI and AscI patterns were used to assign a new type of combined PFGE according to the standardized optimized PulseNet protocol for L. monocytogenes [31] and the PulseNet Bionumerics manual (http://www.pulsenetinternational.org/protocols/bionumerics/, accessed on 1 January 2019). The Dice correlation coefficient was applied to identify similarities between the PFGE types with a tolerance of 1.5% and an optimization of 0.5%, generating a single dendrogram using the Unweighted-Pair Group Matching Algorithm (UPGMA).
2.7. Antimicrobial Susceptibility Test
The antimicrobial susceptibility of the 45 whole-genome sequenced isolates was determined by broth microdilution according to the Clinical and Laboratory Standards Institute (CLSI) [34]. In this procedure, a panel of 8 antimicrobial agents was used: ampicillin (AMP), ciprofloxacin (CIP), erythromycin (ERY), gentamicin (GEN), tetracycline (TET), vancomycin (VA), meropenem (MEM) and cefoxitin (FOX). All antibiotics were purchased from Sigma-Aldrich, Darmstadt, Germany. Staphylococcus aureus ATCC29213, Enterococcus faecalis ATCC29212 and L. monocytogenes ATCCBAA-67 were used as quality control strains. MIC breakpoints were those established by the European Committee on Antimicrobial Susceptibility Testing (EUCAST) [35]. As in the case of FOX, CIP, GEN, TET and VA, there are no interpretive criteria available for L. monocytogenes, we applied the breakpoint defined by EUCAST for Staphylococcus spp. Isolates were considered multidrug-resistant (MDR) when they showed non-susceptibility to at least one agent in three or more antimicrobial categories [36].
2.8. DNA Extraction and Sequencing
The total DNA from the L. monocytogenes isolates was purified with the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) and sequenced on a MiSeq device using reagents kit v3 for 2 × 300 paired-end libraries (Illumina, San Diego, CA, USA) as previously described [37].
2.9. Bioinformatics Analysis
The raw reads were analyzed using the pipeline TORMES® version 1.0 [38]. Genome assembly was performed with SPAdes [39] and Quast [40] and genome annotation with Prokka [41]. Taxonomic confirmation was performed by using Kraken2 [42]. Additionally, 16S rRNA genes were extracted from each genome with Barrnap and used for taxonomic classification by using the RDP Classifier [43] at a confidence level of 0.8. Multilocus sequence typing was performed using an open source tool (MLST, T. Seemann, https://github.com/tseemann/mlst, accessed on 1 January 2020). Search of antibiotic resistance genes was performed by screening the genome against Resfinder [44], CARD [45] and ARG-ANNOT [46] databases by using ABRIcate (https://GitHub-tseemann/abricate: Mass screening of contigs for antimicrobial and virulence genes, accessed on 1 January 2020). Any hit with coverage and/or identity below 90% was removed. Pangenome was created with Roary [47] and FastTree [48]. The search of virulence genes was performed by screening the genome against the Virulence Factors Database (VFDB, [49]) by using ABRIcate. Any hit with coverage and/or identity below 90% was removed. Genes involved in conjugation, mobilization or genes known to be related to Pathogenicity Island LIPI-4, as well as ami and aut variants were detected using a custom blast database and using BLASTx version 2.12.0 [50]. The heatmap showing the virulence genes was made using the pheatmap package under R version 4.1.3 and RStudio RStudio 2022.02.0 (https://CRAN.R-project.org/package=pheatmap, accessed on 1 January 2020). The search of circular plasmids was performed with platon (https://github.com/oschwengers/platon). Further interrogation of integrons and conjugative plasmids was performed using Integron_Finder (https://github.com/gem-pasteur/Integron_Finder, accessed on 1 June 2022) and MacSyFinder (https://github.com/gem-pasteur/macsyfinder, accessed on 1 June 2022) (CONJscan_plasmids [51]), respectively.
2.10. Nucleotide Accession Numbers
The raw fastq files, annotated assembly and L. monocytogenes isolates information are all available under the BioProject ID PRJNA855628.
2.11. Statistical Analyses
All statistical analyses were carried out using GraphPad Prism version 9.4.0 (GraphPad Software, San Diego, CA, USA). To investigate the association between positive L. monocytogenes samples and source frequencies, the χ2 and Fisher´s exact test was performed. In linear regression analyses, the t-test was used to determine whether the slope of the regression line differs significantly from zero. In all cases, differences were considered statistically significant at p < 0.05.
3. Results
3.1. Environmental Sampling, Incidence and L. monocytogenes Isolation
A total of 424 samples were collected and processed from 14 different dairy cattle farms (5 organic and 9 conventional) in the Cantabria region (Northern Spain, Atlantic Coast). Samples were obtained from forage, silage, concentrate, water, raw tank milk and tank milk filters, fecal samples, manure and soil from within and around the stable. Fecal samples were collected from both unhealthy (diarrhea/infection/miscarriage) and apparently healthy dairy cows (without symptoms). The majority of samples (91%, n = 387) were positive for Listeria spp., of which 54% (n = 211) were positive for L. monocytogenes (Table 1). Strains were isolated by conventional microbiological plating methods and presumptive L. monocytogenes isolates were tested by multiplex PCR-based serogrouping, as described in Materials and Methods. Each of the 14 farms yielded at least one positive L. monocytogenes sample. Environmental samples obtained from sites around the farms related to feces (soil, slurry spreader and slurry drain) and fecal samples (stored manure and fresh feces) showed a higher percentage of L. monocytogenes and Listeria spp. in general in comparison with the feed samples and the raw dairy samples (χ2 and Fisher´s exact test p < 0.0001, Table 1). L. monocytogenes in particular, and Listeria spp. in general, were found in both organic and conventional farms with no statistical difference between them (χ2 and Fisher´s exact test p = 0.0960, Table 2). The full list of the L. monocytogenes isolates is shown in the Supplementary Materials (Table S1).

Table 1.
Prevalence of Listeria isolates from different samples from dairy cattle farms in the Cantabria region.
Table 2.
Prevalence of Listeria isolates from organic and conventional dairy cattle farms in the Cantabria region.
3.2. Transmission of L. monocytogenes
To assess the relationship of L. monocytogenes prevalence between sample categories, farm-level prevalence scatter plots were generated, and regression analyses were performed as described before [52]. Scatter plots were constructed for all possible relationships between samples, but only the environmental samples versus stored manure regression analysis had a slope significantly different from zero (p < 0.05) (Figure 1). Since the prevalence of L. monocytogenes in stored manure is higher than in environmental samples (Table 1) and assuming that transmission is expected to happen from high-contaminated to less-contaminated sites, these data indicate that L. monocytogenes is possibly transmitted through the manure to the farm ecosystem.
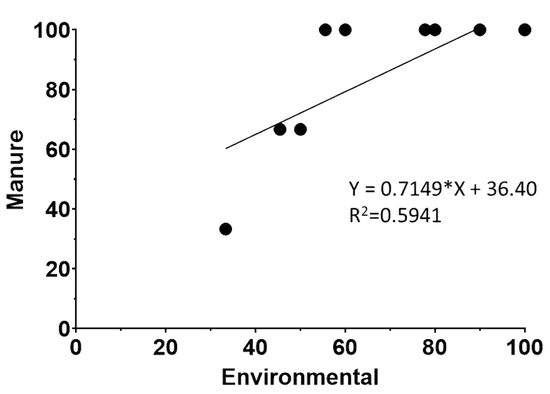
Figure 1.
Scatter plot of percentage of L. monocytogenes positive manure samples versus L. monocytogenes positive environmental samples. Regression equation and R2 values are determined by regression analysis as described in Material and Methods. The (*) in the equation corresponds to the multiplication sign.
3.3. Decline in L. monocytogenes in Livestock Waste
One of the L. monocytogenes isolates (MS6507, isolated from rectal swab sample from cow) was selected to assess its ability to survive in manure. L. monocytogenes CIP103575 was used as a control strain. Liquid manure samples were inoculated, and survival curves were created. L. monocytogenes was detected until 28 days without enrichment. After that point, enrichment was performed before plating to confirm the presence/absence of L. monocytogenes, which was detected for one more week, until 35 days had passed. Negative control tanks always yielded negative results for L. monocytogenes. The results are shown in Figure 2A, expressed as the average ± standard deviation (n = 6).
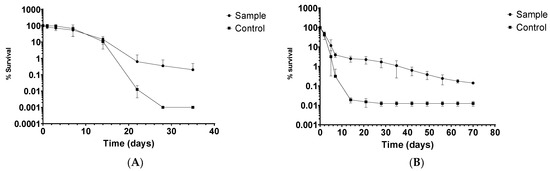
Figure 2.
Death curve of L. monocytogenes in (A) livestock waste and (B) crops fertilized with infected manure.
3.4. Decline in L. monocytogenes in Pasture Crops
The stored manure was inoculated with L. monocytogenes and applied to pasture crops in a controlled laboratory setting. L. monocytogenes was detected until 15 days without enrichment. After that point, enrichment was performed before plating to confirm the presence/absence of L. monocytogenes. The results are shown in Figure 2B, expressed as the average ± standard deviation (n = 4). Maximum survival time on crops was surprisingly high, reaching 70 days.
3.5. PCR Serogroups
The 89% of the isolates (n = 187 out of 211) were identified as PCR serogroup IVb, 5% (n = 11 out of 211) as IIa and 6% (n = 13 out of 211) as serotype IIb. All three PCR serogroups were found in both organic and conventional farms with no statistical difference (χ2 and Fisher´s exact test p = 0.2479). Farms had at least two different PCR serogroups, except one organic farm in which only IVb was found.
3.6. Pulsed-Field Gel Electrophoresis (PFGE)
A total of 211 L. monocytogenes isolates were characterized using PFGE in order to elucidate the genetic relationship among them. The PFGE profiles were analyzed and compared using BioNumerics. The PFGE analysis using ApaI and AscI yielded 62 and 40 restriction profiles, respectively (representative PFGE profiles are shown in Figure 3A). The combination of the two restriction enzymes resulted in 64 restriction PFGE types or pulsotypes with 53% of the pulsotypes (34 out of 64) isolated from unique sites and not repeated among different farms (Figure 3B). In addition, the total of the farms showed a high diversity of pulsotypes, as more than one pulsotype was found within the same farm. On the other hand, isolates with the same pulsotype were isolated from geographically distant farms (Supplementary Materials Table S1).
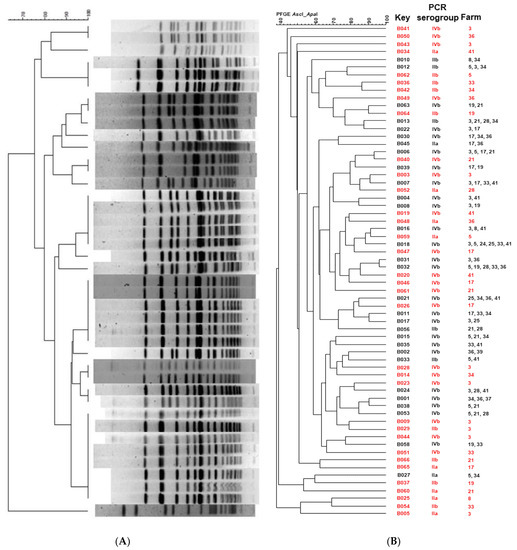
Figure 3.
(A) Representative L. monocytogenes PFGE profiles obtained with ApaI. (B) Dendrogram showing similarities among the total number of the L. monocytogenes PFGE profiles obtained with ApaI and AscI. In red are shown pulsotypes found in unique sites (not repeated between farms).
3.7. Genome Analysis of the L. monocytogenes Isolates and Multilocus Sequence Typing (MLST)
The molecular basis of the clonal relatedness and the virulence genes repertoire of 45 L. monocytogenes isolates was examined by whole genome sequencing (WGS). For the selection of the samples to be sequenced, the following criteria were employed: isolates should be of PCR serogroup IV, should represent different pulsotypes and should have been isolated from farms with information available about antibiotics use. Summary and statistics for the genome sequencing of the L. monocytogenes isolates are detailed in the Supplementary Materials (Table S2). The pangenome (the set of all genes that are present in the analyzed dataset) consisted of 4240 genes, with a core-genome (the pool of conserved genes, which are represented in all genomes included in the analysis) of 2545 (60%) genes; softcore-genome (the set of genes present in at least 95% of genomes analyzed) of 83 (3%); shell-genome (the pool of genes moderately common in the pangenome; more than 15% but less than 95%) of 434 (10%) and cloud-genome (the genes present in less than 15% of the genomes analyzed) of 1178 (27%). Plasmids were not detected in any of the isolates. Nevertheless, genes involved on conjugation and/or mobilization of plasmids were detected in 25 of the 45 isolates, using a custom blast database in BLASTx [50] and MacSyFinder [51]. The results are given in Supplementary Materials (Tables S3 and S4).
In silico MLST determination was performed, and showed that the 45 sequenced L. monocytogenes isolates belonged to one of the following 12 different sequence types (STs): ST1, ST2, ST4, ST6, ST54, ST59, ST217, ST388, ST389, ST489, ST666 and one novel sequence type ST2921 with the following housekeeping allele combination: abcZ(1), bglA(1), cat(12), dapE(659), dat(2), ldh(1), lhkA(3). ST1 was the most abundant (13 out of 45 isolates, followed by ST666 (7 out of 45 strains), ST54 (7 out of 45 strains), ST388 (5 out of 45 strains), ST6 (4 out of 45 strains), ST217 (3 out of 45 strains) and ST2, ST4, ST59, ST389, ST489 and ST2921 (1 out of 45 strains, each) (Table 3). In the approximately-maximum-likelihood phylogenetic trees from the alignments of the accessory and the core genes (Figure 4A,B, respectively), the isolates are grouped according to the CCs that they belong.
Table 3.
In silico multilocus sequence typing (MLST) of the 45 sequenced L. monocytogenes isolates. In bold, isolates from organic farms.
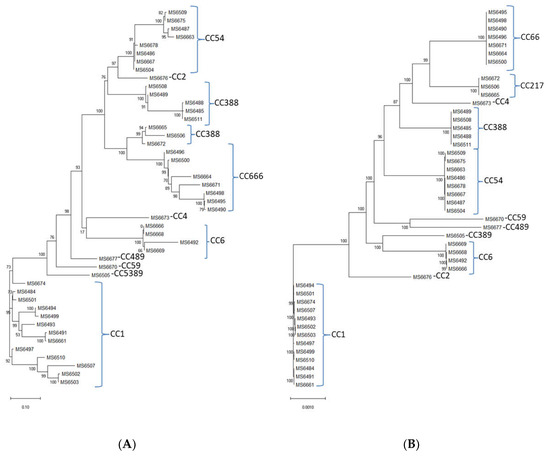
Figure 4.
The approximately-maximum-likelihood phylogenetic trees from the alignments of the (A) accessory and the (B) core genes of the L. monocytogenes isolates.
3.8. In Silico Analysis of Antimicrobial Resistance Genes and In Vitro Antimicrobial Susceptibility Testing
Only four antimicrobial resistance genes, present in all of the 45 L. monocytogenes isolates, were detected: fosX (involved in fosfomycin resistance and detected with ResFinder and CARD database), lin (involved in lincomycin resistance and detected with CARD and ARG-ANNOT databases), norB (conferring quinolone resistance and detected with CARD database) and mprF (which protects against cationic peptides and detected with CARD database).
The 45 isolates used for the WGS were subjected to antimicrobial susceptibility testing by using a panel of eight antimicrobial agents: ampicillin (AMP), ciprofloxacin (CIP), erythromycin (ERY), gentamicin (GEN), tetracycline (TET), vancomycin (VAN), meropenem (MEM) and cefoxitin (FOX). All isolates were susceptible to GEN, but resistant to FOX. The percentages of resistance to the rest of antimicrobial agents were as follows: AMP (15.5%, n = 7), TET (28.9%, n = 13), MEM (11%, n = 5), CIP (11%, n = 5), ERY (6.7%, n = 3) and VAN (4.4%, n = 2). Six isolates were found to be multidrug-resistant (MDR), as displayed resistance to at least 3 drugs in different antimicrobial categories (Table 4 and Supplementary Materials Table S5).
Table 4.
Antimicrobial susceptibility of L. monocytogenes isolates characterized as multi-drug resistant (MDR). Values correspond to minimum inhibitory concentrations (MIC) and expressed in mg/mL.
There was no statistically significant difference in antimicrobial resistance ratios between organic and conventional farms (χ2 and Fisher´s exact test p = 0.2337). The results are given in Supplementary Materials (Table S3). Of the 6 MDR isolates, only one [MS6499(CC1)] was isolated from an organic farm; the other five [MS6484(CC1), MS6488(CC388), MS6490(CC666), MS6485(CC388) and MS6501(CC1)] were isolated from conventional farms.
3.9. In Silico Analysis of Virulence Genes
Virulence genes were detected in silico using TORMES® version 1.0 by screening the L. monocytogenes isolates genomes against the Virulence Factors Data Base (VFDB) [49] and using the L. monocytogenes strain EGD-e as reference. For Listeria pathogenicity island 4 (LIPI-4) the L. monocytogenes strain CLIP80459 was used as reference and was detected using a custom blast database and using BLASTx version 2.12.0 [50]. The majority of virulence genes were shared across the 45 sequenced L. monocytogenes genomes (Figure 5). All the sequenced isolates harbored inlA, inlB, inlC, inlJ and inlK, encoding proteins of the “internalin” family of L. monocytogenes, which interact with distinct host receptors to promote infection of human cells [53]. An additional gene encoding a member of the internalin family, inlF, was also present in most isolates (39 out of 45). All sequenced isolates contained the pathogenicity island LIPI-1, a 9 kb DNA fragment composed of six genes (prfA, mpl, plcA, plcB, actA and hly), the products of which are required for the intracellular life cycle of L. monocytogenes [54]. Most isolates (38 out of 45) harbored a 6 kb DNA fragment corresponding to the pathogenicity cluster LIPI-3 composed of eight genes (llsA, llsB, llsD, llsG, llsH, llsP, llsX and llsY) which encodes listeriolysin O, a pore-forming toxin involved in virulence [55]. On the other hand, the pathogenicity island LIP-4, a system strongly associated with the central nervous system and placental infections, was only found in 9 isolates. Other genes involved in virulence, present in all sequenced isolates are prsA2 gene, coding a critical post-translocation secretion chaperone [56] and loci encoding the Clp stress tolerance mediators, ClpC, ClpE and ClpP, involved in intraphagosomal survival [57,58,59].
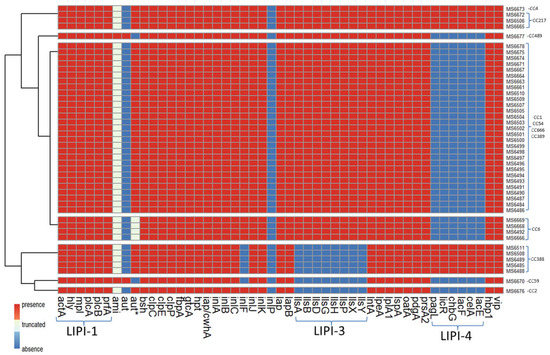
Figure 5.
Presence/absence of virulence genes of L. monocytogenes isolates. aut* refers to the short variant of aut; LIPI-1: Listeria pathogenicity island 1; LIPI-3: Listeria pathogenicity island 3; LIPI-4: Listeria pathogenicity island 4.
Only two isolates [MS6670 (CC59) and MS6677 (CC489)] harbored ami gene, which codes for an autolysin required for adhesion to eukaryotic cells [60]. However, the rest of the isolates (43 out of 45) contained truncated versions of ami gene leading to a truncated protein with 772 or 607 amino acids instead of 934 as in the canonical Ami sequence (Figure 6A). The same two isolates [MS6670 (CC59) and MS6677 (CC489)] that harbored ami gene, contained aut gene also, which codes for another autolysin, required for entry into eukaryotic cells [61]. The shorter aut variant LMOF2365_RS00075 [62] was found in all the rest of the sequenced isolates. Four isolates belonging to CC6 (MS6492, MS6666, MS6668 and MS6669) showed a deletion of six amino acids in the shorter Aut variant (Figure 6B).

Figure 6.
Sequence alignment of (A) Ami and (B) Aut with the respective variants found in the L. monocytogenes isolates. The figure shows a CLUSTALW alignment. Colour code: red on yellow background=identical residues; Black on green background = similar residues; Green on white background=weakly conserved residues; Blue on light blue background: strongly conserved residues; Black on white background = non-conserved residues. Accession Numbers are: Ami = NP_466081.1; Aut (short variant) = WP_003734189.1.
4. Discussion
Listeria monocytogenes is a facultative foodborne pathogen and several listeriosis outbreaks have been linked to its presence in the farm environment, either because of fresh vegetables fertilized with infected manure or because of the consumption of contaminated fresh dairy products [21,63]. The results of the present study demonstrate that samples from sites related to feces (slurry spreader, stable floor, slurry drain, manure and fresh feces) had a higher prevalence of L. monocytogenes than the ones found in feed and raw dairy samples (Table 1). These data support the hypothesis that on-farm transmission appears to be due to ingestion of feed contaminated with L. monocytogenes and, afterward, fecal shedding of L. monocytogenes in the environment by the bovine hosts, both with and without clinical disease [8]. In a way, livestock contributes to the increase and spread of L. monocytogenes into the farm environment, a hypothesis that is consistent with previous studies [52,64].
Several studies conducted under controlled laboratory conditions confirmed that L. monocytogenes can grow on plants [65,66,67]. The main drawback of most of those studies is the absence of other microorganisms. As a result, these experiments are best-case scenarios demonstrating that L. monocytogenes can colonize the surface of most plants. Whether or not L. monocytogenes colonizes plants internally is still a matter of debate, and conflicting reports are available [67,68,69]. Nevertheless, data available so far favor the hypothesis that L. monocytogenes can utilize nutrients from the plants to multiply and survive on plant surfaces. This hypothesis could explain the extended survival time of L. monocytogenes in crops, as shown in Figure 2B. Even though the experiment was performed in controlled laboratory conditions and the presence of rhizosphere and phyllosphere microbiome was limited, the maximum survival time of L. monocytogenes on crops was surprisingly high (70 days). The results of the decline of L. monocytogenes in pasture crops in combination with its decline in manure (Figure 2A) demonstrate that the life cycle of this pathogen between dairy animals and their surroundings is continuous. In order to reduce the contamination risk, effective herd management aiming the reduction of intestinal carriage, and manure treatment and management, when used as fertilizer in grassland and cropland, should be applied [8].
In this study, three PCR serogroups were isolated: IIa (lineage II) and IIb and IVb (lineage I). Comparatively few studies have isolates from natural environments and/or animals characterized to PCR serogroup and PFGE subtype. In general, according to previous studies, L. monocytogenes lineage I and lineage II isolates seem to be similarly prevalent [18,52,70], though contamination patterns of lineage I and II may differ [23]. Nevertheless, in this study, 95% of the isolates belonged to lineage I. Usually, linage I isolates display higher tolerance to low pH [71], which can be advantageous for the survival of L. monocytogenes in the gastric acidic environment and may account for their association with the fecal samples. Taking into account the prevalence of lineage I isolates in this study, it can be an additional indicator that the presence of L. monocytogenes in the farm environment is due to fecal contamination. Similar results were observed in poultry carcasses [23].
L. monocytogenes isolates were characterized using PFGE in order to elucidate their genetic resemblance. A total of 211 isolates were subtyped by PFGE, using ApaI and AscI endonucleases resulting in 64 PFGE types or pulsotypes (Figure 3). The pulsotypes were separated by their PCR serogroup, results that are in accordance with previous studies [72,73] that confirmed relationships between serotype and PFGE patterns and reveal that the L. monocytogenes population of dairy farms in the Cantabria region is genetically diverse. From these 64 pulsotypes, 45 isolates were selected for whole genome sequencing, employing the following criteria: isolates should be of PCR serogroup IVb, represent different pulsotypes and be isolates from farms with information available about the antibiotics use. The reason why the PCR serogroup IVb was selected instead of IIa and IIb, which were also isolated from the dairy farms, is because the PCR serogroup IVb strains are commonly implicated in outbreaks and clinical human listeriosis cases. Previous studies showed that serotype 4b displays the highest pathogenicity compared to other serotypes in zebrafish embryos [74] and highest virulence than other serotypes according to reports that used mouse and Galleria mellonella as a model for identifying virulence determinants [75,76]. These features make the PCR serogroup IVb strains a very interesting candidate for further analysis at the genomic level.
A diversity of STs was found, as the 45 sequenced isolates were grouped in 12 different STs. Except for a new ST [ST2921(CC54)], isolated from an organic dairy farm, the rest of the L. monocytogenes isolates sequenced in this study included STs already reported. The most prevalent ST (Table 3) was ST1 (29% of all the sequenced isolates), followed by ST6 (15% of all the sequenced isolates), both of them related to outbreaks and clinical cases [77,78]. The prevalence of ST1/CC1 in ruminant-associated L. monocytogenes isolates has also been reported in another recent study conducted in Spain [79]. Interestingly, clones ST1/CC1, ST2/CC2, ST4/CC4, and ST6/CC6 constitute almost half of the sequenced L. monocytogenes isolates of this study (18 out of 45). The predominance of those clones in dairy products and ruminant feces has been already demonstrated and as they invade more efficiently the gut, they probably could lead to a higher fecal shedding and release of L. moonocytogenes in the environment [80]. Of particular concern is the isolation of ST388 (CC388), as it has been identified as the cause of a severe outbreak that took place in 2019 in Spain (data from the Spanish Ministry of Health). These findings highlight the importance of surveillance programs in farm animals.
Antibiotic resistance is one of the major concerns of public health since the percentage of infections due to resistant bacteria is increasing [81]. From the 45 isolates tested for their minimum inhibitory concentration (MIC), only six were characterized as multidrug-resistant. All 45 isolates were resistant to cefoxitin, an expected result, since Listeria spp. are naturally resistant to cephalosporins [2,82]. Overall, our results confirm the susceptibility of L. monocytogenes. However, some isolates were found resistant to antimicrobials used to treat human cases of listeriosis, such as ampicillin and tetracycline, and even though the antimicrobial resistance among L. monocytogenes isolates is still low, there are reports of increasing resistance. This is an important issue since it is known that L. monocytogenes can acquire and/or transfer resistance genes through horizontal transfer processes in the intestinal microbiota [83]. In a recent study, 61% of L. monocytogenes isolates from a meat facility contained conjugative and/or mobilizable plasmids [84]. In contrast, no plasmids were detected in the isolates described here. Nevertheless, genes responsible for plasmid conjugation and/or mobilization were detected by MacSyFinder, with protein profiles mostly belonging to the MOBP1 Hidden Markov Models (HMMs) profile (Table S4). The MOBP1 HMMs profile targets relaxases from the MOBP conjugative transfer system [85], a diverse superfamily divided in seven subfamilies (MOBP1-MOBP7) [86]. The conjugative transfer systems are classified in six MOB families: MOBF, MOBH, MOBQ, MOBC, MOBP and MOBV, with MOBP being the most diverse as it can be detected in plasmids from various incompatibility groups [86]. Since proteins from the MOBP conjugative transfer system is present in many conjugative and mobilizable plasmids [87] we can only hypothesize that conjugative and/or mobilizable plasmids might be present in the isolates studied, but it was not possible to detect them with the tools used.
The presence of core genome virulence factors and pathogenicity islands was evaluated for the 45 sequenced isolates in this study (Figure 5). Four Listeria pathogenicity islands (LIPI) have been verified thus far in the literature, involved in the invasion, survival and colonization of Listeria in host tissues. Three of them have been described for L. monocytogenes (LIPI-1, LIPI-3 and LIPI-4), whereas LIPI-2 has only been detected in L. ivanovii [88]. LIPI-1 contains six genes: hly, prfA, plcA, plcB, mpl and actaA [89,90]. It encodes virulence factors that enable bacteria to escape from the vacuole (endosome or phagosome), proliferate in the cytosol and spread to the adjacent cells. LIPI-3 is composed of eight genes: llsA, llsB, llsD, llsG, llsH, llsP, and llsX. It encodes listeriolysin S (LLS), a haemolysin acting as a bacteriocin able to alter the host intestinal microbiota [91]. LIPI-4 encodes a cellobiose-family phosphotransfer system (PTS) and is involved in neural and placental infection [76]. Each of the 45 isolates sequenced in this study had at least one LIPI, with LIPI-1 being the one present in all strains. LIPI-3 was absent in ST2/CC2, as reported before [74], and also in CC59 and CC388. Isolates from CC4 and CC217 had all three LIPI, indicating the hypervirulence nature of those isolates. Originally the presence of LIPI-4 was reported only in strains from CC4 [76] and later other strains such as ST217/CC217 and ST388/CC388 were reported for carrying LIPI-4 [92,93], confirming the results of the present study.
In addition to the three pathogenicity islands described above, L. monocytogenes expresses a variety of virulence factors that are essential for its survival and persistence in the gastrointestinal tract and sequenced isolates contained various genes associated with virulence. For example, all isolates harbored the virulence genes encoding for internalins A, B, C, and J (inlA, inlB, inlC and inlJ), some cover surface proteins important in bacterial adhesion and invasion [94]. The inlK gene was also present in all of the sequenced isolates, encoding for a protein, InlK, which is involved in the autophagic recognition escape of L. monocytogenes [95]. Another gene encoding for a member of the internalin family of surface proteins, inlF, was found in 39 out of 45 isolates; its translational product, InlF, was reported of playing a role in L. monocytogenes colonization of the brain in vivo [96]. The mechanism of pathogenesis in L. monocytogenes involves a diversity of virulence proteins and the mechanisms leading to virulence are not completely clear yet. However, the presence of inlA, inlB, inlC, inlF, inlJ and inlK genes suggests the virulence potential of the L. monocytogenes isolates described in this study. Surprisingly, only two of the sequenced isolates harbored the ami gene, coding the Ami protein, involved in attachment to host cells and bacterial colonization of hepatocytes [60,97]. After closer inspection, all of the remaining 43 sequenced isolates not containing the ami gene, contained a truncated ami gene, leading to either a truncated 772- or a 607-amino acid Ami variants instead of the 934-amino acid wild type Ami protein (Figure 6A). Since the initial search of virulence genes was performed by screening the genome against Virulence Factors DataBase (VFDB, [49]) by using ABRIcate and accepting only hits with coverage and/or identity above 90%, the truncated version of ami was not originally detected but revealed after using a custom blast database and using BLASTx [50]. This deletion has been already reported by other authors [74]. The same pattern was observed for the aut, a gene that encodes a surface protein with autolytic activity required for invasion into eucaryotic cells and as a consequence for virulence in vitro [61]. After the first analysis, aut gene was found absent in all sequenced isolates, except in two isolates that also harbored the full-length ami gene (Figure 5). After closer inspection, it was revealed that the rest of the sequenced isolates harbored the shorter aut variant LMOF2365_RS00075 already reported by other authors [62]. A new variation of the aut allele was found in four isolates from CC6 (MS6492, MS6666, MS6668 and MS6669) showing a deletion of six amino acids (Figure 6B). Whether or not the truncated ami gene and aut allele found in the isolates described in this study lead to non-virulent or less virulent strains in vivo, remains to be tested and was beyond the objectives of the present study. In any case, our data demonstrate that dairy farms constitute a reservoir for possibly hypervirulent L. monocytogenes, which are shed in the feces and could easily colonize dairy products, as a result of fecal contamination.
5. Conclusions
An analysis of L. monocytogenes isolated from environmental sites of livestock and livestock farms has shown that several L. monocytogenes, responsible for human infection, circulate in the biosphere and agricultural systems and might contribute to the spread of these pathogens throughout the food chain, thereby posing a major health challenge. A significant portion of isolates recovered are from the same CCs as those frequently isolated from human clinical cases and outbreaks on a global scale and many of those strains encode virulence factors associated with serious illness. As the SARS-CoV2 outbreak has demonstrated, the spread of infectious diseases in humans from animal reservoirs represents a major public health risk and it is expected that zoonotic diseases will occur more often due to climate change. This is the main pillar of the “One Health” approach, which accepts that human health is tightly connected with animal health and the environment. Considering the ability of L. monocytogenes to survive and adapt in different ecosystems and hosts, which makes it an important archetype of the “One Health” axis, effective herd and manure management and respect of the animal welfare are needed to prevent foodborne illnesses. Farmers should pay attention especially to the transmission from animal to animal through fecal-oral routes, usually via manure contamination of the pasture or silage with the microorganism. Feeding the animals with good quality silage, avoiding any other rotten vegetation and isolation of sick animals following good hygiene and sanitation on the farm is also important. Thereafter, food safety programs throughout the food production chain (from farm to fork) are needed to prevent foodborne illnesses.
Supplementary Materials
The following supporting information is available on line and can be downloaded at: https://www.mdpi.com/article/10.3390/ani12182477/s1: Table S1: The complete list of L. moncytogenes isolates; Table S2: Assembly statistics of the L. moncytogenes isolates; Table S3: Genes involved in conjugation/mobilization of plasmids detected in the L. monocytogenes isolates using a custom blast database in BLASTx; Table S4: Genes involved in conjugation/mobilization of plasmids detected in the L. monocytogenes isolates using MacSyFinder; Table S5: Antimicrobial susceptibility of L. monocytogenes isolates.
Author Contributions
A.V.: performed most experiments, conceived the study, acquired funds, analyzed the results, wrote, review and edited the last version of the manuscript; S.O.: performed experiments, reviewed and edited last version of the manuscript; P.S.: Isolated the strains, conceived the study, acquired funds, reviewed and edited the last version of the manuscript; P.L.: performed experiments, reviewed and edited last version of the manuscript; V.L.-A.: performed experiments, reviewed and edited last version of the manuscript; M.H.: performed the sequencing experiments, reviewed and edited the last version of the manuscript; D.A.: performed genome assembly and bioinformatics analysis, reviewed and edited last version of the manuscript; J.R.-G.: performed bioinformatics analysis, reviewed and edited last version of the manuscript; A.A.O.-S.: performed susceptibility tests, reviewed and edited the last version of the manuscript; J.V.M.-S.: acquired funds, conceived the study, reviewed and edited the last version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Research Project grants RTA08-099, RTA2008-00080-C02, RTA2014-00045-C03-01 (INIA and FEDER) from the Spanish Ministry of Economy and Competitiveness and RTI2018-098267-R-C31 (INIA and FEDER) from the Spanish Ministry of Science and Innovation.
Institutional Review Board Statement
No laboratory animals were used during this study. Samples were collected from 14 commercial dairy farms in the Cantabria region (Spain), after the permission of the farmers/owners and in accordance with the EU directive UE 2016/679 on “General data protection regulation”. Samples collected from rectal swabs were from Holstein-Friesian dairy cows, housed free and sampling procedures were in accordance with the EU Directive 2010/63/EU “on the protection of animals used for scientific purposes”. The study was approved by the Ministry of Livestock, Fisheries and Rural Development, Government of Cantabria (Spain) and by The Official College of Veterinarians, according to the European Veterinary Code of Conduct.
Informed Consent Statement
Not applicable.
Data Availability Statement
Genome sequences information is available in http://www.ncbi.nlm.nih.gov/bioproject/, (accessed on 1 June 2022) under the BioProject ID PRJNA855628.
Acknowledgments
The authors want to thank the Government of Cantabria for supporting the work performed, the local farmers for their collaboration and the curators of the Listeria MLST system at the Pasteur Institute, Paris, France (http:://bigsdb.pasteur.fr, accessed on 17 March 2022) for assignment of novel ST.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Roberts, A.J.; Wiedmann, M. Pathogen, Host and Environmental Factors Contributing to the Pathogenesis of Listeriosis. Cell. Mol. Life Sci. 2003, 60, 904–918. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Lanza, V.F.; Duval, M.; Coque, T.M. Ecogenetics of Antibiotic Resistance in Listeria monocytogenes. Mol. Microbiol. 2020, 113, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, E.; Courvalin, P. Antibiotic Resistance in Listeria spp. Antimicrob. Agents Chemother. 1999, 43, 2103. [Google Scholar] [CrossRef] [PubMed]
- Noll, M.; Kleta, S.; Al Dahouk, S. Antibiotic Susceptibility of 259 Listeria monocytogenes Strains Isolated from Food, Food-Processing Plants and Human Samples in Germany. J. Infect. Public Health 2018, 11, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Kayode, A.J.; Okoh, A.I. Assessment of Multidrug-Resistant Listeria monocytogenes in Milk and Milk Product and One Health Perspective. PLoS ONE 2022, 17, e0270993. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.A.C.; Carvalho, F.T.; Vallim, D.C.; Pereira, R.C.L.; Neto, A.C.; Vieira, B.S.; Carvalho, R.C.T.; Figueiredo, E.E.S. Listeria monocytogenes in Export-Approved Beef from Mato Grosso, Brazil: Prevalence, Molecular Characterization and Resistance to Antibiotics and Disinfectants. Microorganisms 2019, 8, 18. [Google Scholar] [CrossRef]
- Rabiey, S.; Hosseini, H.; Rezaei, M. The Hurdle Effect of Bunium Persicum Essential Oil, Smoke and NaCl for Controlling the Listeria monocytogenes Growth in Fish Model Systems. J. Food Saf. 2013, 33, 137–144. [Google Scholar] [CrossRef]
- Santorum, P.; Garcia, R.; Lopez, V.; Martinez-Suarez, J.V. Dairy Farm Management and Production Practices Associated with the Presence of Listeria monocytogenes in Raw Milk and Beef. Span. J. Agric. Res. 2012, 10, 360–371. [Google Scholar] [CrossRef]
- Oevermann, A.; Zurbriggen, A.; Vandevelde, M. Rhombencephalitis Caused by Listeria monocytogenes in Humans and Ruminants: A Zoonosis on the Rise? Interdiscip. Perspect. Infect. Dis. 2010, 2010, 632513. [Google Scholar] [CrossRef]
- Van Kessel, J.S.; Santin-Duran, M.; Karns, J.S.; Schukken, Y. Tracing Zoonotic Pathogens in Dairy Production. Tracing zoonotic pathogens in dairy production. In Tracing Pathogens in the Food Chain; Brul, S., Fratamico, P.M., McMeekin, T.A., Eds.; Woodhead Publishing Limited: Philadelphia, PA, USA, 2011; pp. 503–526. [Google Scholar] [CrossRef]
- NicAogáin, K.; O’Byrne, C.P. The Role of Stress and Stress Adaptations in Determining the Fate of the Bacterial Pathogen Listeria monocytogenes in the Food Chain. Front. Microbiol. 2016, 7, 1865. [Google Scholar] [CrossRef]
- Fenlon, D.R. Wild Birds and Silage as Reservoirs of Listeria in the Agricultural Environment. J. Appl. Bacteriol. 1985, 59, 537–543. [Google Scholar] [CrossRef]
- Hasegawa, M.; Iwabuchi, E.; Yamamoto, S.; Muramatsu, M.; Takashima, I.; Hirai, K. Prevalence and Characteristics of Listeria monocytogenes in Feces of Black Beef Cattle Reared in Three Geographically Distant Areas in Japan. Foodborne Pathog. Dis. 2014, 11, 96–103. [Google Scholar] [CrossRef]
- Fox, E.; O’Mahony, T.; Clancy, M.; Dempsey, R.; O’Brien, M.; Jordan, K. Listeria monocytogenes in the Irish Dairy Farm Environment. J. Food Prot. 2009, 72, 1450–1456. [Google Scholar] [CrossRef]
- Haley, B.J.; Sonnier, J.; Schukken, Y.H.; Karns, J.S.; Van Kessel, J.A.S. Diversity of Listeria monocytogenes within a U.S. Dairy Herd, 2004–2010. Foodborne Pathog. Dis. 2015, 12, 844–850. [Google Scholar] [CrossRef]
- Latorre, A.A.; Van Kessel, J.S.; Karns, J.S.; Zurakowski, M.J.; Pradhan, A.K.; Boor, K.J.; Jayarao, B.M.; Houser, B.A.; Daugherty, C.S.; Schukken, Y.H. Biofilm in Milking Equipment on a Dairy Farm as a Potential Source of Bulk Tank Milk Contamination with Listeria monocytogenes. J. Dairy Sci. 2010, 93, 2792–2802. [Google Scholar] [CrossRef]
- Van Kessel, J.S.; Karns, J.S.; Gorski, L.; McCluskey, B.J.; Perdue, M.L. Prevalence of Salmonellae, Listeria monocytogenes, and Fecal Coliforms in Bulk Tank Milk on US Dairies. J. Dairy Sci. 2004, 87, 2822–2830. [Google Scholar] [CrossRef]
- Orsi, R.H.; de Bakker, H.C.; Wiedmann, M. Listeria monocytogenes Lineages: Genomics, Evolution, Ecology, and Phenotypic Characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef]
- Burall, L.S.; Grim, C.J.; Datta, A.R. A Clade of Listeria monocytogenes Serotype 4b Variant Strains Linked to Recent Listeriosis Outbreaks Associated with Produce from a Defined Geographic Region in the US. PLoS ONE 2017, 12, e0176912. [Google Scholar] [CrossRef]
- Jeršek, B.; Gilot, P.; Gubina, M.; Klun, N.; Mehle, J.; Tcherneva, E.; Rijpens, N.; Herman, L. Typing of Listeria Monocytogenes Strains by Repetitive Element Sequence-Based PCR. J. Clin. Microbiol. 1999, 37, 103–109. [Google Scholar] [CrossRef]
- Swaminathan, B.; Gerner-Smidt, P. The Epidemiology of Human Listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef]
- Martin, P.; Jacquet, C.; Goulet, V.; Vaillant, V.; De Valk, H. Pulsed-Field Gel Electrophoresis of Listeria monocytogenes Strains: The PulseNet Europe Feasibility Study. Foodbourne Pathog. Dis. 2006, 3, 303–308. [Google Scholar] [CrossRef]
- López, V.; Ortiz, S.; Corujo, A.; López, P.; Poza, D.; Navas, J.; Moreno, R.; Martínez-Suárez, J.V. Different Contamination Patterns of Lineage I and II Strains of Listeria monocytogenes in a Spanish Broiler Abattoir. Poult. Sci. 2008, 87, 1874–1882. [Google Scholar] [CrossRef]
- Ragon, M.; Wirth, T.; Hollandt, F.; Lavenir, R.; Lecuit, M. A New Perspective on Listeria monocytogenes Evolution. PLoS Pathog. 2008, 4, 1000146. [Google Scholar] [CrossRef]
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Björkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V.; et al. Whole Genome-Based Population Biology and Epidemiological Surveillance of Listeria monocytogenes. Nat. Microbiol. 2016, 2, 16185. [Google Scholar] [CrossRef]
- Maiden, M.C.J.; Van Rensburg, M.J.J.; Bray, J.E.; Earle, S.G.; Ford, S.A.; Jolley, K.A.; McCarthy, N.D. MLST Revisited: The Gene-by-Gene Approach to Bacterial Genomics. Nat. Rev. Microbiol. 2013, 11, 728–736. [Google Scholar] [CrossRef]
- Van Walle, I.; Björkman, J.T.; Cormican, M.; Dallman, T.; Mossong, J.; Moura, A.; Pietzka, A.; Ruppitsch, W.; Takkinen, J.; Mattheus, W.; et al. Retrospective Validation of Whole Genome Sequencingenhanced Surveillance of Listeriosis in Europe, 2010 to 2015. Eurosurveillance 2018, 23, 1700798. [Google Scholar] [CrossRef] [PubMed]
- Varsaki, A.; Ortiz, S.; Santorum, P.; López, P.; López-Alonso, V.; Martínez-Suárez, J.V. Genetic Diversity, Antimicrobial Resistance and Survival upon Manure Storage of Campylobacter jejuni Isolated from Dairy Cattle Farms in the Cantabric Coast of Spain. Zoonotic Dis. 2022, 2, 82–94. [Google Scholar] [CrossRef]
- Doumith, M.; Buchrieser, C.; Glaser, P.; Jacquet, C.; Martin, P. Differentiation of the Major Listeria monocytogenes Serovars by Multiplex PCR. J. Clin. Microbiol. 2004, 42, 3819. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, A.; Chenal-Francisque, V.; Dieye, H.; Cantinelli, T.; Drali, R.; Brisse, S.; Lecuit, M. Characterization of the Novel Listeria monocytogenes PCR Serogrouping Profile IVb-V1. Int. J. Food Microbiol. 2011, 147, 74–77. [Google Scholar] [CrossRef]
- Halpin, J.L.; Garrett, N.M.; Ribot, E.M.; Graves, L.M.; Cooper, K.L. Re-Evaluation, Optimization, and Multilaboratory Validation of the PulseNet-Standardized Pulsed-Field Gel Electrophoresis Protocol for Listeria monocytogenes. Foodborne Pathog. Dis. 2010, 7, 293–298. [Google Scholar] [CrossRef]
- Ortiz, S.; López, V.; Villatoro, D.; López, P.; Dávila, J.C.; Martínez-Suárez, J.V. A 3-Year Surveillance of the Genetic Diversity and Persistence of Listeria monocytogenes in an Iberian Pig Slaughterhouse and Processing Plant. Foodborne Pathog. Dis. 2010, 7, 1177–1184. [Google Scholar] [CrossRef]
- Graves, L.M.; Swaminathan, B. PulseNet Standardized Protocol for Subtyping Listeria monocytogenes by Macrorestriction and Pulsed-Field Gel Electrophoresis. Int. J. Food Microbiol. 2001, 65, 55–62. [Google Scholar] [CrossRef]
- CLSI Clinical and Laboratory Standards Institude. Performance Standards for Antimicrobial Susceptibility Testing. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 15 January 2022).
- EUCAST. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 28 March 2022).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Hernández, M.; Iglesias, M.R.; Rodríguez-Lázaro, D.; Gallardo, A.; Quijada, N.M.; Miguela-Villoldo, P.; Campos, M.J.; Píriz, S.; López-Orozco, G.; de Frutos, C.; et al. Co-Occurrence of Colistin-Resistance Genes Mcr-1 and Mcr-3 among Multidrug-Resistant Escherichia coli Isolated from Cattle, Spain, September 2015. Eurosurveillance 2017, 22, 30586. [Google Scholar] [CrossRef]
- Quijada, N.M.; Rodríguez-Lázaro, D.; Eiros, J.M.; Hernández, M. TORMES: An Automated Pipeline for Whole Bacterial Genome Analysis. Bioinformatics 2019, 35, 4207–4212. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved Metagenomic Analysis with Kraken2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The Comprehensive Antibiotic Resistance Database. Antimicrob. Agents Chemother. 2013, 57, 3348. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a New Bioinformatic Tool to Discover Antibiotic Resistance Genes in Bacterial Genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid Large-Scale Prokaryote Pan Genome Analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing Large Minimum Evolution Trees with Profiles Instead of a Distance Matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A Reference Database for Bacterial Virulence Factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Cury, J.; Abby, S.S.; Doppelt-Azeroual, O.; Néron, B.; Rocha, E.P.C. Identifying Conjugative Plasmids and Integrative Conjugative Elements with CONJscan. Methods Mol. Biol. 2020, 2075, 265–283. [Google Scholar] [CrossRef]
- Nightingale, K.K.; Schukken, Y.H.; Nightingale, C.R.; Fortes, E.D.; Ho, A.J.; Her, Z.; Grohn, Y.T.; McDonough, P.L.; Wiedmann, M. Ecology and Transmission of Listeria monocytogenes Infecting Ruminants and in the Farm Environment. Appl. Environ. Microbiol. 2004, 70, 4458–4467. [Google Scholar] [CrossRef]
- Ireton, K.; Mortuza, R.; Gyanwali, G.C.; Gianfelice, A.; Hussain, M. Role of Internalin Proteins in the Pathogenesis of Listeria monocytogenes. Mol. Microbiol. 2021, 116, 1407–1419. [Google Scholar] [CrossRef]
- Vázquez-Boland, J.A.; Domínguez-Bernal, G.; González-Zorn, B.; Kreft, J.; Goebel, W. Pathogenicity Islands and Virulence Evolution in Listeria. Microbes Infect. 2001, 3, 571–584. [Google Scholar] [CrossRef]
- Cotter, P.D.; Draper, L.A.; Lawton, E.M.; Daly, K.M.; Groeger, D.S.; Casey, P.G.; Ross, R.P.; Hill, C. Listeriolysin S, a Novel Peptide Haemolysin Associated with a Subset of Lineage I Listeria monocytogenes. PLoS Pathog. 2008, 4, e1000144. [Google Scholar] [CrossRef] [PubMed]
- Cahoon, L.A.; Freitag, N.E. Listeria monocytogenes Virulence Factor Secretion: Don’t Leave the Cell without a Chaperone. Front. Cell. Infect. Microbiol. 2014, 4, 13. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Milohanic, E.; Berche, P. ClpC ATPase Is Required for Cell Adhesion and Invasion of Listeria monocytogenes. Infect. Immun. 2000, 68, 7061–7068. [Google Scholar] [CrossRef]
- Nair, S.; Frehel, C.; Nguyen, L.; Escuyer, V.; Berche, P. ClpE, a Novel Member of the HSP100 Family, Is Involved in Cell Division and Virulence of Listeria monocytogenes. Mol. Microbiol. 1999, 31, 185–196. [Google Scholar] [CrossRef]
- Gaillot, O.; Bregenholt, S.; Jaubert, F.; Di Santo, J.P.; Berche, P. Stress-Induced ClpP Serine Protease of Listeria monocytogenes Is Essential for Induction of Listeriolysin O-Dependent Protective Immunity. Infect. Immun. 2001, 69, 4938. [Google Scholar] [CrossRef]
- Milohanic, E.; Jonquières, R.; Cossart, P.; Berche, P.; Gaillard, J.-L. The Autolysin Ami Contributes to the Adhesion of Listeria monocytogenes to Eukaryotic Cells via Its Cell Wall Anchor. Mol. Microbiol. 2001, 39, 1212–1224. [Google Scholar] [CrossRef]
- Cabanes, D.; Dussurget, O.; Dehoux, P.; Cossart, P. Auto, a Surface Associated Autolysin of Listeria Monocytogenes Required for Entry into Eukaryotic Cells and Virulence. Mol. Microbiol. 2004, 51, 1601–1614. [Google Scholar] [CrossRef]
- Painset, A.; Björkman, J.T.; Kiil, K.; Guillier, L.; Mariet, J.F.; Felix, B.; Amar, C.; Rotariu, O.; Roussel, S.; Perez-Reche, F.; et al. Liseq—Whole-Genome Sequencing of a Cross-Sectional Survey of Listeria monocytogenes in Ready-to-Eat Foods and Human Clinical Cases in Europe. Microb. Genomics 2019, 5, e000257. [Google Scholar] [CrossRef]
- Shenoy, A.G.; Oliver, H.F.; Deering, A.J. Listeria monocytogenes Internalizes in Romaine Lettuce Grown in Greenhouse Conditions. J. Food Prot. 2017, 80, 573–581. [Google Scholar] [CrossRef]
- Fenlon, D.R.; Wilson, J.; Donachie, W. The Incidence and Level of Listeria monocytogenes Contamination of Food Sources at Primary Production and Initial Processing. J. Appl. Bacteriol. 1996, 81, 641–650. [Google Scholar] [CrossRef]
- Bardsley, C.A.; Boyer, R.R.; Rideout, S.L.; Strawn, L.K. Survival of Listeria monocytogenes on the Surface of Basil, Cilantro, Dill, and Parsley Plants. Food Control 2019, 95, 90–94. [Google Scholar] [CrossRef]
- Kljujev, I.; Raicevic, V.; Jovicic-Petrovic, J.; Vujovic, B.; Mirkovic, M.; Rothballer, M. Listeria monocytogenes—Danger for Health Safety Vegetable Production. Microb. Pathog. 2018, 120, 23–31. [Google Scholar] [CrossRef]
- Truong, H.N.; Garmyn, D.; Gal, L.; Fournier, C.; Sevellec, Y.; Jeandroz, S.; Piveteau, P. Plants as a Realized Niche for Listeria monocytogenes. Microbiologyopen 2021, 10, e1255. [Google Scholar] [CrossRef]
- Kõiv, V.; Arbo, K.; Maiväli, Ü.; Kisand, V.; Roosaare, M.; Remm, M.; Tenson, T. Endophytic Bacterial Communities in Peels and Pulp of Five Root Vegetables. PLoS ONE 2019, 14, e0210542. [Google Scholar] [CrossRef]
- Kutter, S.; Hartmann, A.; Schmid, M. Colonization of Barley (Hordeum Vulgare) with Salmonella enterica and Listeria spp. FEMS Microbiol. Ecol. 2006, 56, 262–271. [Google Scholar] [CrossRef]
- Lyautey, E.; Lapen, D.R.; Wilkes, G.; McCleary, K.; Pagotto, F.; Tyler, K.; Hartmann, A.; Piveteau, P.; Rieu, A.; Robertson, W.J.; et al. Distribution and Characteristics of Listeria monocytogenes Isolates from Surface Waters of the South Nation River Watershed, Ontario, Canada. Appl. Environ. Microbiol. 2007, 73, 5401–5410. [Google Scholar] [CrossRef]
- Hingston, P.; Chen, J.; Dhillon, B.K.; Laing, C.; Bertelli, C.; Gannon, V.; Tasara, T.; Allen, K.; Brinkman, F.S.L.; Hansen, L.T.; et al. Genotypes Associated with Listeria monocytogenes Isolates Displaying Impaired or Enhanced Tolerances to Cold, Salt, Acid, or Desiccation Stress. Front. Microbiol. 2017, 8, 369. [Google Scholar] [CrossRef]
- Lukinmaa, S.; Aarnisalo, K.; Suihko, M.L.; Siitonen, A. Diversity of Listeria monocytogenes Isolates of Human and Food Origin Studied by Serotyping, Automated Ribotyping and Pulsed-Field Gel Electrophoresis. Clin. Microbiol. Infect. 2004, 10, 562–568. [Google Scholar] [CrossRef][Green Version]
- Esteban, J.I.; Oporto, B.; Aduriz, G.; Juste, R.A.; Hurtado, A. Faecal Shedding and Strain Diversity of Listeria monocytogenes in Healthy Ruminants and Swine in Northern Spain. BMC Vet. Res. 2009, 5, 2. [Google Scholar] [CrossRef]
- Muchaamba, F.; Eshwar, A.K.; Stevens, M.J.A.; Stephan, R.; Tasara, T. Different Shades of Listeria monocytogenes: Strain, Serotype, and Lineage-Based Variability in Virulence and Stress Tolerance Profiles. Front. Microbiol. 2022, 12, 792162. [Google Scholar] [CrossRef]
- Lee, B.H.; Garmyn, D.; Gal, L.; Guérin, C.; Guillier, L.; Rico, A.; Rotter, B.; Nicolas, P.; Piveteau, P. Exploring Listeria monocytogenes Transcriptomes in Correlation with Divergence of Lineages and Virulence as Measured in Galleria mellonella. Appl. Environ. Microbiol. 2019, 85, e01370-19. [Google Scholar] [CrossRef]
- Maury, M.M.; Tsai, Y.H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, A.; et al. Uncovering Listeria monocytogenes Hypervirulence by Harnessing Its Biodiversity. Nat. Genet. 2016, 48, 308. [Google Scholar] [CrossRef]
- Cabal, A.; Pietzka, A.; Huhulescu, S.; Allerberger, F.; Ruppitsch, W.; Schmid, D. Isolate-Based Surveillance of Listeria monocytogenes by Whole Genome Sequencing in Austria. Front. Microbiol. 2019, 10, 2282. [Google Scholar] [CrossRef]
- Halbedel, S.; Wilking, H.; Holzer, A.; Kleta, S.; Fischer, M.A.; Lüth, S.; Pietzka, A.; Huhulescu, S.; Lachmann, R.; Krings, A.; et al. Large Nationwide Outbreak of Invasive Listeriosis Associated with Blood Sausage, Germany, 2018–2019. Emerg. Infect. Dis. 2020, 26, 1456–1464. [Google Scholar] [CrossRef]
- Palacios-Gorba, C.; Moura, A.; Gomis, J.; Leclercq, A.; Gómez-Martín, Á.; Bracq-Dieye, H.; Mocé, M.L.; Tessaud-Rita, N.; Jiménez-Trigos, E.; Vales, G.; et al. Ruminant-Associated Listeria monocytogenes Isolates Belong Preferentially to Dairy-Associated Hypervirulent Clones: A Longitudinal Study in 19 Farms. Environ. Microbiol. 2021, 23, 7617–7631. [Google Scholar] [CrossRef]
- Maury, M.M.; Bracq-Dieye, H.; Huang, L.; Vales, G.; Lavina, M.; Thouvenot, P.; Disson, O.; Leclercq, A.; Brisse, S.; Lecuit, M. Hypervirulent Listeria Monocytogenes Clones’ Adaption to Mammalian Gut Accounts for Their Association with Dairy Products. Nat. Commun. 2019, 10, 2488. [Google Scholar] [CrossRef]
- Huemer, M.; Mairpady Shambat, S.; Brugger, S.D.; Zinkernagel, A.S. Antibiotic resistance and persistence-Implications for human health and treatment perspectives. EMBO Rep. 2020, 21, e51034. [Google Scholar] [CrossRef]
- Haubert, L.; Mendonça, M.; Lopes, G.V.; de Itapema Cardoso, M.R.; da Silva, W.P. Listeria monocytogenes Isolates from Food and Food Environment Harbouring TetM and ErmB Resistance Genes. Lett. Appl. Microbiol. 2016, 62, 23–29. [Google Scholar] [CrossRef]
- Neil, K.; Allard, N.; Rodrigue, S. Molecular Mechanisms Influencing Bacterial Conjugation in the Intestinal Microbiota. Front. Microbiol. 2021, 12, 1415. [Google Scholar] [CrossRef]
- Alvarez-Molina, A.; Cobo-Díaz, J.F.; López, M.; Prieto, M.; de Toro, M.; Alvarez-Ordóñez, A. Unraveling the Emergence and Population Diversity of Listeria monocytogenes in a Newly Built Meat Facility through Whole Genome Sequencing. Int. J. Food Microbiol. 2021, 340, 109043. [Google Scholar] [CrossRef] [PubMed]
- Garcillán-Barcia, M.P.; Redondo-Salvo, S.; Vielva, L.; de la Cruz, F. MOBscan: Automated annotation of MOB relaxases. Methods Mol. Biol. 2020, 2075, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Garcillán-Barcia, M.P.; Francia, M.V.; de la Cruz, F. The diversity of conjugative relaxases and its application in plasmid classification. FEMS Microbiol Rev. 2009, 33, 657–687. [Google Scholar] [CrossRef] [PubMed]
- Coluzzi, C.; Garcillán-Barcia, M.D.P.; de la Cruz, F.; Rocha, E.P.C. Evolution of Plasmid Mobility: Origin and Fate of Non-Conjugative Plasmids. bioRxiv 2021. [Google Scholar] [CrossRef]
- Disson, O.; Moura, A.; Lecuit, M. Making sense of the biodiversity and virulence of Listeria monocytogenes. Trends Microbiol. 2021, 29, 811–822. [Google Scholar] [CrossRef]
- Hadjilouka, A.; Paramithiotis, S.; Drosinos, E.H. Genetic Analysis of the Listeria Pathogenicity Island 1 of Listeria monocytogenes 1/2a and 4b Isolates. Curr. Microbiol. 2018, 75, 857–865. [Google Scholar] [CrossRef]
- Poimenidou, S.V.; Dalmasso, M.; Papadimitriou, K.; Fox, E.M.; Skandamis, P.N.; Jordan, K. Virulence Gene Sequencing Highlights Similarities and Differences in Sequences in Listeria monocytogenes Serotype 1/2a and 4b Strains of Clinical and Food Origin from 3 Different Geographic Locations. Front. Microbiol. 2018, 9, 1103. [Google Scholar] [CrossRef]
- Quereda, J.J.; Dussurget, O.; Nahori, M.A.; Ghozlane, A.; Volant, S.; Dillies, M.A.; Regnault, B.; Kennedy, S.; Mondot, S.; Villoing, B.; et al. Bacteriocin from Epidemic Listeria Strains Alters the Host Intestinal Microbiota to Favor Infection. Proc. Natl. Acad. Sci. USA 2016, 113, 5706–5711. [Google Scholar] [CrossRef]
- Chen, M.; Cheng, J.; Wu, Q.; Zhang, J.; Chen, Y.; Xue, L.; Lei, T.; Zeng, H.; Wu, S.; Ye, Q.; et al. Occurrence, Antibiotic Resistance, and Population Diversity of Listeria monocytogenes isolated from Fresh Aquatic Products in China. Front. Microbiol. 2018, 9, 2215. [Google Scholar] [CrossRef]
- Lee, S.; Chen, Y.; Gorski, L.; Ward, T.J.; Osborne, J.; Kathariou, S. Listeria monocytogenes Source Distribution Analysis Indicates Regional Heterogeneity and Ecological Niche Preference among Serotype 4b Clones. mBio 2018, 9, e00396-18. [Google Scholar] [CrossRef]
- Quereda, J.J.; Rodríguez-Gómez, I.M.; Meza-Torres, J.; Gómez-Laguna, J.; Nahori, M.A.; Dussurget, O.; Carrasco, L.; Cossart, P.; Pizarro-Cerdá, J. Reassessing the Role of Internalin B in Listeria monocytogenes Virulence Using the Epidemic Strain F2365. Clin. Microbiol. Infect. 2019, 25, 252.e1–252.e4. [Google Scholar] [CrossRef]
- Dortet, L.; Mostowy, S.; Louaka, A.S.; Gouin, E.; Nahori, M.A.; Wiemer, E.A.C.; Dussurget, O.; Cossart, P. Recruitment of the Major Vault Protein by InlK: A Listeria monocytogenes Strategy to Avoid Autophagy. PLoS Pathog. 2011, 7, e1002168. [Google Scholar] [CrossRef]
- Ghosh, P.; Halvorsen, E.M.; Ammendolia, D.A.; Mor-Vaknin, N.; O’Riordan, M.X.D.; Brumell, J.H.; Markovitz, D.M.; Higgins, D.E. Invasion of the Brain by Listeria monocytogenes Is Mediated by InlF and Host Cell Vimentin. mBio 2018, 9, e00160-18. [Google Scholar] [CrossRef]
- Asano, K.; Sashinami, H.; Osanai, A.; Asano, Y.; Nakane, A. Autolysin Amidase of Listeria monocytogenes Promotes Efficient Colonization of Mouse Hepatocytes and Enhances Host Immune Response. Int. J. Med. Microbiol. 2011, 301, 480–487. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).