
Effect of Tributyrin on Growth Performance and Pathway by which Tributyrin Regulates Oligopeptide Transporter 1 in Juvenile Grass Carp (Ctenopharyngodon idellus)
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diet
2.2. Feeding Experiment Design
2.3. Sample Collection
2.4. Grass Carp Intestinal Cell Culture
2.5. Intraperitoneal Injection Trials
2.6. RNA Extraction and cDNA Synthesis
2.7. Quantitative Real Time PCR
2.8. Statistical Analysis
2.9. Ethics Statement
3. Results
3.1. Appropriate Tributyrin Supplementation Promotes the Growth of Grass Carp
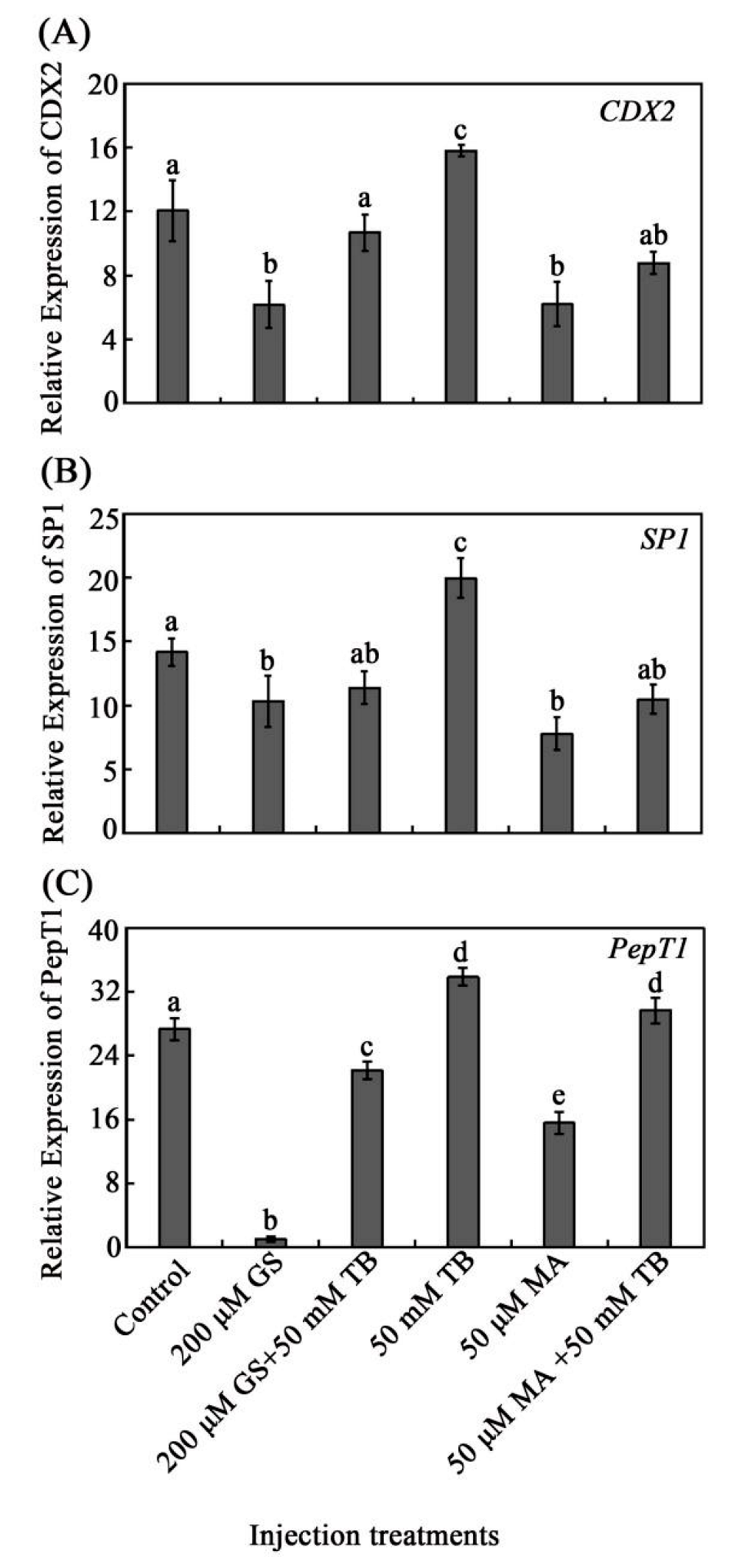
3.2. Tributyrin Regulates the Expressions of CDX2, SP1, and PepT1 In Vivo and In Vitro
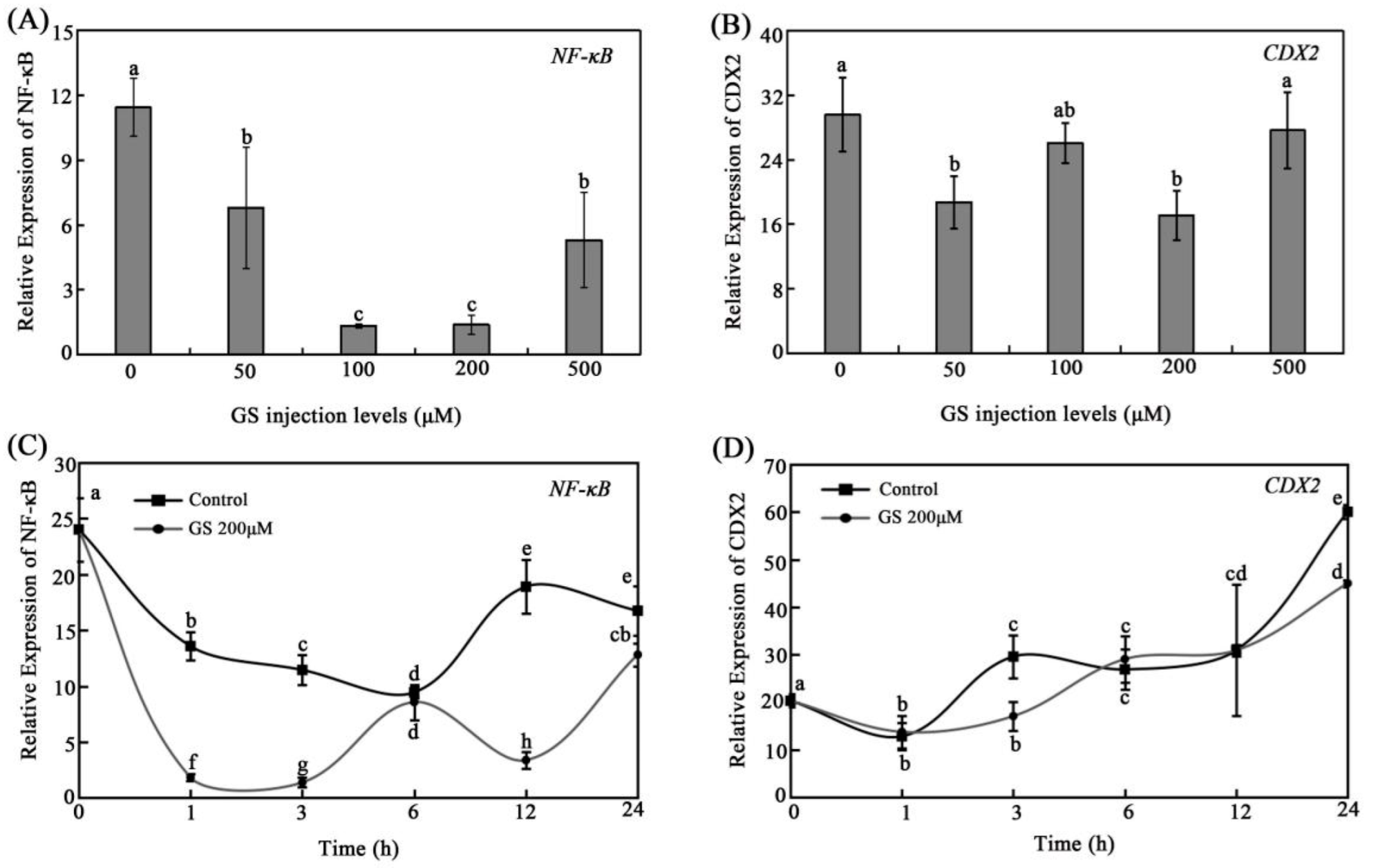
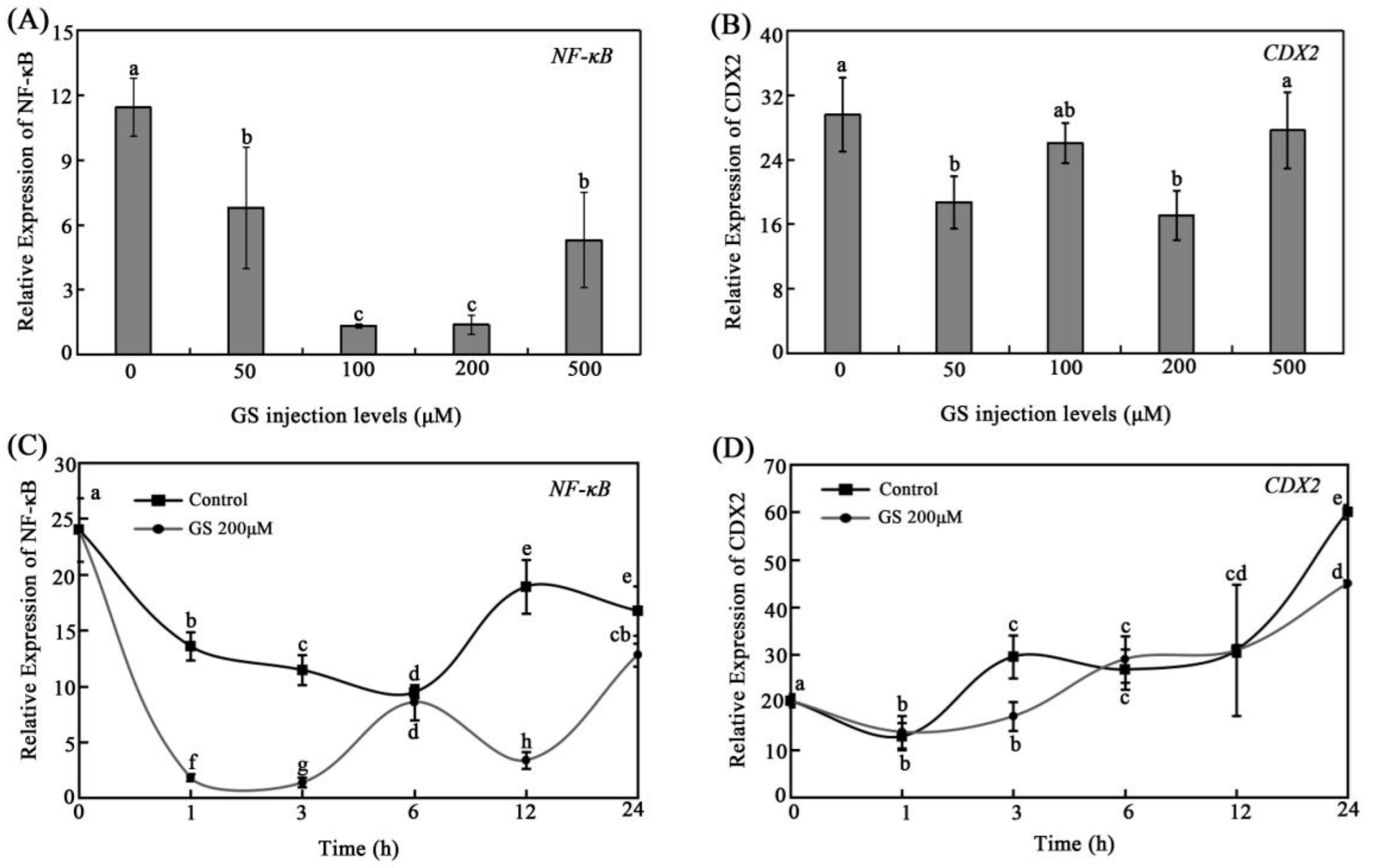
3.3. Effects of GS on Intestinal NF-KB and CDX2 Expression In Vivo
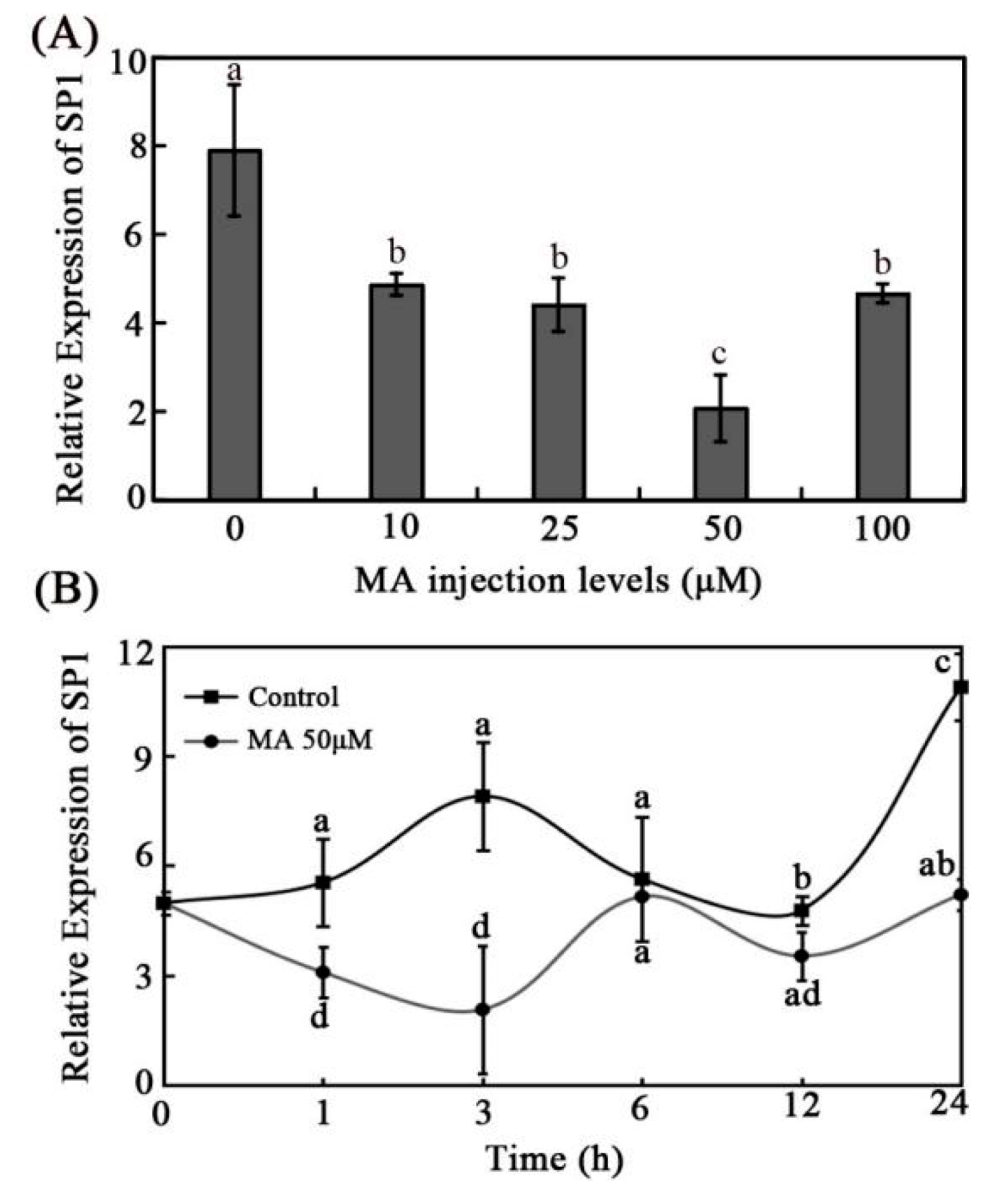
3.4. Effects of MA on Intestinal SP1 Expression In Vivo
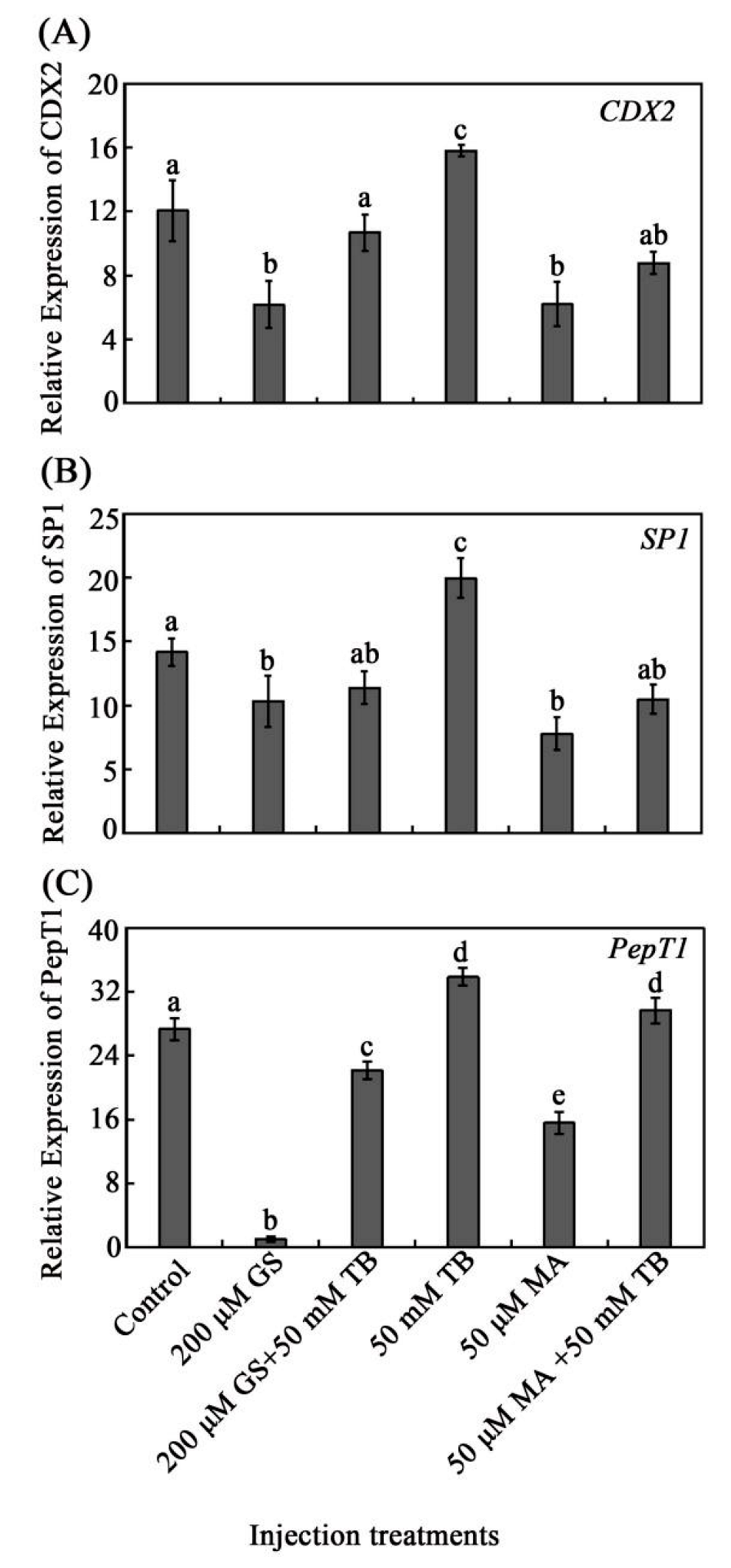
3.5. The Analysis of CDX2/SP1 Mediating the Regulation of Tributyrin on the Expression PepT1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sandri, S. Antiangiogenic Effects of the chemopreventive agent tributyrin, a butyric acid prodrug, during the promotion phase of hepatocarcinogenesis. Carcinogenesis 2019, 40, 979–988. [Google Scholar] [CrossRef]
- Schaefer, L.; Hernandez, H.; Coasts, R.A.; Yu, Z.; Pflugfelder, S.C.; Britton, R.A.; de Paiva, C.S. Gut-derived butyrate suppresses ocular surface inflammation. Sci. Rep. 2022, 12, 4512. [Google Scholar] [CrossRef]
- Su, J.; Zhang, N.; Ho, P.C. Determination of tributyrin and its metabolite butyrate in Wistar rat plasma samples by gas chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 2217–2222. [Google Scholar] [CrossRef] [PubMed]
- Jirsova, Z.; Heczkova, M.; Dankova, H.; Malinska, H.; Videnska, P.; Vespalcova, H.; Micenkova, L.; Bartonova, L.; Sticova, E.; Lodererova, A.; et al. The effect of butyrate-supplemented parenteral nutrition on intestinal defence mechanisms and the parenteral nutrition-induced shift in the gut microbiota in the rat model. Biomed Res. Int. 2019, 2019, 7084734. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, J.; Liu, Z.; Jiang, A.; Li, S.; Wu, D.; Zhang, Y.; Zhu, X.; Zhou, E.; Wei, Z.; et al. Sodium Butyrate Alleviates Lipopolysaccharide-Induced Inflammatory Responses by Down-Regulation of NF-kappaB, NLRP3 Signaling Pathway, and Activating Histone Acetylation in Bovine Macrophages. Front. Vet. Sci. 2020, 7, 579674. [Google Scholar] [CrossRef] [PubMed]
- Roh, S.; Kimura, N.; Sakamoto, K.; Nishihara, K.; Suzuki, K.; Katoh, K. Effects of butyrate supplementation in antibiotic-free milk replacer and starter on growth performance in suckling calves. Anim. Sci. J. 2018, 89, 1486–1491. [Google Scholar] [CrossRef]
- Yin, B.; Liu, H.; Tan, B.; Dong, X.; Chi, S.; Yang, Q.; Zhang, S. Dietary supplementation of beta-conglycinin, with or without sodium butyrate on the growth, immune response and intestinal health of hybrid grouper. Sci. Rep. 2021, 11, 17298. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, G.; Wang, H.; Mo, W.; Huang, Y.; Cao, J.; Li, P. Effects of dietary sodium butyrate on growth, digestive enzymes, body composition and nutrient retention-related gene expression of juvenile yellow catfish (Pelteobagrus fulvidraco). Anim. Nutr. 2021, 7, 539–547. [Google Scholar] [CrossRef]
- Alves Jesus, G.F.; Owatari, M.S.; Pereira, S.A.; Silva, B.C.; Syracuse, N.M.; Lopes, G.R.; Addam, K.; Cardoso, L.; Pedreira Mourino, J.L.; Martins, M.L. Effects of sodium butyrate and Lippia origanoides essential oil blend on growth, intestinal microbiota, histology, and haemato-immunological response of Nile tilapia. Fish Shellfish Immunol. 2021, 117, 62–69. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Hendam, B.M.; Shukry, M.; El-Shafai, N.M.; El-Mehasseb, I.M.; Dawood, M.A.O.; Abdel-Tawwab, M. Effects of sodium butyrate nanoparticles on the hemato-immunological indices, hepatic antioxidant capacity, and gene expression responses in Oreochromisniloticus. Fish Shellfish Immunol. 2021, 119, 516–523. [Google Scholar] [CrossRef]
- Tomasz, B.; Katarzyna, B.K.; Adam, K.; Anna, K.; Piotr, P.; Romuald, Z.; Piotr, K. Butyric acid in bowel inflammations. Gastroenterol. Rev. 2010, 5, 251–257. [Google Scholar] [CrossRef]
- Hebb, A.L.; Zacharko, R.M.; Dominguez, H.; Trudel, F.; Laforest, S.; Drolet, G. Odor-induced variation in anxiety-like behavior in mice is associated with discrete and differential effects on mesocorticolimbic cholecystokinin mRNA expression. Neuropsychopharmacology 2002, 27, 744–755. [Google Scholar] [CrossRef]
- Moquet, P.C.A.; Onrust, L.; Van Immerseel, F.; Ducatelle, R.; Hendriks, W.H.; Kwakkel, R.P. Importance of release location on the mode of action of butyrate derivatives in the avian gastrointestinal tract. World’s Poult. Sci. J. 2016, 72, 61–80. [Google Scholar] [CrossRef]
- Donovan, J.D.; Bauer, L.; Fahey, G.C., Jr.; Lee, Y. In Vitro Digestion and Fermentation of Microencapsulated Tributyrin for the Delivery of Butyrate. J. Food Sci. 2017, 82, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Gaschott, T.; Steinhilber, D.; Milovic, V.; Stein, J. Tributyrin, a stable and rapidly absorbed prodrug of butyric acid, enhances antiproliferative effects of dihydroxycholecalciferol in human colon cancer cells. J. Nutr. 2001, 131, 1839–1843. [Google Scholar] [CrossRef]
- Zhang, W.X.; Zhang, Y.; Zhang, X.W.; Deng, Z.X.; Liu, J.X.; He, M.L.; Wang, H.F. Effects of Dietary Supplementation with Combination of Tributyrin and Essential Oil on Gut Health and Microbiota of Weaned Piglets. Animals 2020, 10, 180. [Google Scholar] [CrossRef]
- Miragoli, F.; Patrone, V.; Prandini, A.; Sigolo, S.; Dell’Anno, M.; Rossi, L.; Senizza, A.; Morelli, L.; Callegari, M.L. Implications of Tributyrin on Gut Microbiota Shifts Related to Performances of Weaning Piglets. Microorganisms 2021, 9, 584. [Google Scholar] [CrossRef]
- Sotira, S.; Dell’Anno, M.; Caprarulo, V.; Hejna, M.; Pirrone, F.; Callegari, M.L.; Tucci, T.V.; Rossi, L. Effects of Tributyrin Supplementation on Growth Performance, Insulin, Blood Metabolites and Gut Microbiota in Weaned Piglets. Animals 2020, 10, 726. [Google Scholar] [CrossRef]
- Chen, G.; Zhuo, R.; Ding, H.; Yang, K.; Xue, J.; Zhang, S.; Chen, L.; Yin, Y.; Fang, R. Effects of dietary tributyrin and physterol ester supplementation on growth performance, intestinal morphology, microbiota and metabolites in weaned piglets. J. Appl. Microbiol. 2022, 132, 2293–2305. [Google Scholar] [CrossRef]
- Li, Y.; Wang, H.; Zhang, Y.; Li, X.; Jiang, X.; Ding, H. Effects of Dietary Supplementation with Glycerol Monolaurate (GML) or the Combination of GML and Tributyrin on Growth Performance and Rumen Microbiome of Weaned Lambs. Animals 2022, 12, 1309. [Google Scholar] [CrossRef]
- Hu, Q.; Yin, F.; Li, B.; Guo, Y.; Yin, Y. Dietary Tributyrin Administration Improves Intestinal Morphology and Selected Bacterial and Short-Chain Fatty Acid Profiles in Broilers Under an Isocaloric Feeding Regime. Front. Microbiol. 2021, 12, 715712. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Cao, S.; Shen, Z.; Hong, Q.; Feng, J.; Peng, Y.; Hu, C. Effects of dietary tributyrin on intestinal mucosa development, mitochondrial function and AMPK-mTOR pathway in weaned pigs. J. Anim. Sci. Biotechnol. 2019, 10, 93. [Google Scholar] [CrossRef]
- Gong, L.; Xiao, G.; Zheng, L.; Yan, X.; Qi, Q.; Zhu, C.; Feng, X.; Huang, W.; Zhang, H. Effects of Dietary Tributyrin on Growth Performance, Biochemical Indices, and Intestinal Microbiota of Yellow-Feathered Broilers. Animals 2021, 11, 3425. [Google Scholar] [CrossRef]
- Hansen, V.L.; Kahl, S.; Proszkowiec-Weglarz, M.S.; Jimenez, C.; Vaessen, S.F.C.; Schreier, L.L.; Jenkins, M.C.; Russell, B.; Miska, K.B. The effects of tributyrin supplementation on weight gain and intestinal gene expression in broiler chickens during Eimeria maxima-induced coccidiosis. Poult. Sci. 2021, 100, 100984. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Lin, X.; Gou, Z.; Fan, Q.; Jiang, S. Effects of Clostridium butyricum, Sodium Butyrate, and Butyric Acid Glycerides on the Reproductive Performance, Egg Quality, Intestinal Health, and Offspring Performance of Yellow-Feathered Breeder Hens. Front. Microbiol. 2021, 12, 657542. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, H.; Bai, S.; Zeng, Q.; Su, Z.; Zhuo, Y.; Mao, X.; Yin, H.; Feng, B.; Liu, J.; et al. Dietary tributyrin improves reproductive performance, antioxidant capacity, and ovary function of broiler breeders. Poult. Sci. 2021, 100, 101429. [Google Scholar] [CrossRef]
- Wang, J.; Fu, S.; Zou, X.; Luo, C.; Shu, D.; Qu, H. The Impact of Tributyrin on Performance and Intestinal Health of Broiler Chickens Post Coccidiosis Vaccination. Avian Dis. 2021, 65, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Ji, K.; Ge, X.; Xi, B.; Ren, M.; Zhang, L.; Chen, X. Tributyrin Plays an Important Role in Regulating the Growth and Health Status of Juvenile Blunt Snout Bream (Megalobrama amblycephala), as Evidenced by Pathological Examination. Front. Immunol. 2021, 12, 652294. [Google Scholar] [CrossRef] [PubMed]
- Domon-Dell, C.; Wang, Q.; Kim, S.; Kedinger, M.; Evers, B.M.; Freund, J.N. Stimulation of the intestinal Cdx2 homeobox gene by butyrate in colon cancer cells. Gut 2002, 50, 525–529. [Google Scholar] [CrossRef]
- Dalmasso, G.; Nguyen, H.T.; Yan, Y.; Charrier-Hisamuddin, L.; Sitaraman, S.V.; Merlin, D. Butyrate transcriptionally enhances peptide transporter PepT1 expression and activity. PLoS ONE 2008, 3, e2476. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhou, Y.; Liu, S.; Zhao, Q.; Feng, J.; Lu, S.; Xiong, G.; Xie, D.; Zhang, J.; Liu, Y. Characterization and dietary regulation of oligopeptide transporter (PepT1) in different ploidy fishes. Peptides 2014, 52, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Donde, H.; Ghare, S.; Joshi-Barve, S.; Zhang, J.; Vadhanam, M.V.; Gobejishivili, L.; Lorkiewicz, P.; Srivastava, S.; McClain, C.J.; Barve, S. Tributyrin Inhibits Ethanol-Induced Epigenetic Repression of CPT-1A and Attenuates Hepatic Steatosis and Injury. Cell Mol. Gastroenterol. Hepatol. 2020, 9, 569–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Wang, P.Y.; Wang, X.; Wan, Y.L.; Liu, Y.C. Butyrate enhances intestinal epithelial barrier function via up-regulation of tight junction protein Claudin-1 transcription. Dig. Dis. Sci. 2012, 57, 3126–3135. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kawai, Y.; Hanson, R.W.; Arinze, I.J. Sodium butyrate induces transcription from the G alpha(i2) gene promoter through multiple Sp1 sites in the promoter and by activating the MEK-ERK signal transduction pathway. J. Biol. Chem. 2001, 276, 25742–25752. [Google Scholar] [CrossRef]
- Yu, D.C.; Waby, J.S.; Chirakkal, H.; Staton, C.A.; Corfe, B.M. Butyrate suppresses expression of neuropilin I in colorectal cell lines through inhibition of Sp1 transactivation. Mol. Cancer 2010, 9, 276. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, Y.; Feng, J.; Lu, S.; Zhao, Q.; Zhang, J. Characterization of oligopeptide transporter (PepT1) in grass carp (Ctenopharyngodon idella). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2013, 164, 194–200. [Google Scholar] [CrossRef]
- Shimakura, J.; Terada, T.; Katsura, T.; Inui, K. Characterization of the human peptide transporter PEPT1 promoter: Sp1 functions as a basal transcriptional regulator of human PEPT1. American journal of physiology. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G471–G477. [Google Scholar] [CrossRef]
- He, Z.M.; Cai, Y.Y.; Yang, M.; Liu, N.; Zeng, Z.H.; Li, X.J.; Zhou, X.M.; Liu, S.C.; Liu, Z. Transcription factor CDX2 directly regulates the expression of Ctenopharyngodon idellus intestinal PepT1 to mediate the transportation of oligopeptide. Aquacult. Rep. 2022, 24, 1011148. [Google Scholar] [CrossRef]
- Luo, W.J.; Song, P.; He, Z.M.; Cao, S.P.; Tang, J.Z.; Xu, W.Q.; Xiong, D.; Qu, F.F.; Zhao, D.F.; Liu, Z.; et al. Jak2 mediates the regulation of pept1 expression by leptin in the grass carp (ctenopharyngodon idella) intestine. Front. Physiol. 2020, 11, 79. [Google Scholar] [CrossRef]
- Yamada, T.; Osawa, S.; Ikuma, M.; Kajimura, M.; Sugimoto, M.; Furuta, T.; Iwaizumi, M.; Sugimoto, K. Guggulsterone, a plant-derived inhibitor of NF-TB, suppresses CDX2 and COX-2 expression and reduces the viability of esophageal adenocarcinoma cells. Digestion 2014, 90, 208–217. [Google Scholar] [CrossRef]
- Quarni, W.; Dutta, R.; Green, R.; Katiri, S.; Patel, B.; Mohapatra, S.S.; Mohapatra, S. Mithramycin A Inhibits Colorectal Cancer Growth by Targeting Cancer Stem Cells. Sci. Rep. 2019, 9, 15202. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Liang, H.; Ren, M.; Ge, X.; Ji, K.; Yu, H.; Maulu, S. Effects of dietary lysine levels on growth performance, whole body composition and gene expression related to glycometabolism and lipid metabolism in grass carp, Ctenopharyngodon idellus fry. Aquaculture 2021, 530, 735806. [Google Scholar] [CrossRef]
- Liu, S.; Wang, J.; Feng, Y.; Ye, Q.; Wen, L.; Xu, G.; Zou, J. Effects of compound antimicrobial peptides on the growth performance, antioxidant and immune responses and disease resistance of grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 2020, 107, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Ming, J.; Ye, J.; Zhang, Y.; Xu, Q.; Yang, X.; Shao, X.; Qiang, J.; Xu, P. Optimal dietary curcumin improved growth performance, and modulated innate immunity, antioxidant capacity and related genes expression of NF-kappaB and Nrf2 signaling pathways in grass carp (Ctenopharyngodon idella) after infection with Aeromonas hydrophila. Fish Shellfish Immunol. 2020, 97, 540–553. [Google Scholar] [CrossRef]
- Ma, P.; Hu, Z.; Li, L.; Li, D.; Tang, R. Dietary selenium promotes the growth performance through growth hormone-insulin-like growth factor and hypothalamic-pituitary-thyroid axes in grass carp (Ctenopharyngodon idella). Fish Physiol. Biochem. 2021, 47, 1313–1327. [Google Scholar] [CrossRef]
- Li, J.; Liu, L.; Liang, X.F.; Yuan, X.; Li, J.; Li, B.; Wang, Q.; Fang, J.; Xue, M.; Wang, J. Modulation of appetite, lipid and glucose metabolism of juvenile grass carp (Ctenopharyngodon idellus) by different dietary protein levels. Fish Physiol. Biochem. 2017, 43, 297–307. [Google Scholar] [CrossRef]
- Li, Z.H.; Li, P.; Wu, Y. Effects of temperature fluctuation on endocrine disturbance of grass carp Ctenopharyngodon idella under mercury chloride stress. Chemosphere 2021, 263, 128137. [Google Scholar] [CrossRef]
- Jinendiran, S.; Archana, R.; Sathishkumar, R.; Kannan, R.; Selvakumar, G.; Sivakumar, N. Dietary Administration of Probiotic Aeromonas veronii V03 on the Modulation of Innate Immunity, Expression of Immune-Related Genes and Disease Resistance Against Aeromonas hydrophila Infection in Common Carp (Cyprinus carpio). Probiotics Antimicrob. Proteins 2021, 13, 1709–1722. [Google Scholar] [CrossRef]
- Sun, Y.; Xiang, Y.; He, M.; Zhang, X.; Wang, S.; Guo, W.; Liu, C.; Cao, Z.; Zhou, Y. Evaluation of Lactococcus lactis HNL12 combined with Schizochytrium limacinum algal meal in diets for humpback grouper (Cromileptes altivelis). Fish Shellfish Immunol. 2019, 94, 880–888. [Google Scholar] [CrossRef]
- Mansour, A.T.; Mial, L.; Espinosa, C.; Garcia-Beltran, J.M.; Ceballos Franscisco, D.C.; Esteban, M.A. Effects of dietary inclusion of Moringa oleifera leaves on growth and some systemic and mucosal immune parameters of seabream. Fish Physiol. Biochem. 2018, 44, 1223–1240. [Google Scholar] [CrossRef]
- Vacca, F.; Barca, A.; Gomes, A.S.; Mazzei, A.; Piccinni, B.; Clinquetti, R.; Del Vecchio, G.; Romano, A.; Ronnestad, I.; Bossi, E.; et al. The peptide transporter 1a of the zebrafish Danio rerio, an emerging model in nutrigenomics and nutrition research: Molecular characterization, functional properties, and expression analysis. Genes Nutr. 2019, 14, 33. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, E.R.; Wong, E.A.; Webb, K.E., Jr. Board-invited review: Peptide absorption and utilization: Implications for animal nutrition and health. J. Anim. Sci. 2008, 86, 2135–2155. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Beguet-Crespel, F.; Marinelli, L.; Jamet, A.; Ledue, F.; Blottiere, H.M.; Lapaque, N. Butyrate produced by gut commensal bacteria activates TGF-beta1 expression through the transcription factor SP1 in human intestinal epithelial cells. Sci. Rep. 2018, 8, 9742. [Google Scholar] [CrossRef] [PubMed]
- Shimakura, J.; Terada, T.; Shimada, Y.; Katsura, T.; Inui, K. The transcription factor Cdx2 regulates the intestine-specific expression of human peptide transporter 1 through functional interaction with Sp1. Biochem. Pharmacol. 2006, 71, 1581–1588. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Percentage Dry Weight (%) |
|---|---|
| Wheat flour | 8.00 |
| Starch | 30.00 |
| Soybean meal | 32.00 |
| Fish meal 1 | 18.50 |
| Soybean oil | 2.50 |
| Fish oil | 3.00 |
| Choline chloride | 0.5 |
| Monocalcium phosphate [Ca(H2PO4)2] | 1.0 |
| Chromium hemitrioxide (Cr2O3) | 0.5 |
| Methyl cellulose | 2.00 |
| Mineral Premix 2 | 2.00 |
| Total | 100.00 |
| Proximate composition (% dry matter) | |
| Crude protein | 30.26 |
| Crude lipid | 8.00 |
| Ash | 5.35 |
| Moisture | 6.15 |
| Carbohydrate | 50.24 |
| Gross energy (MJ/kg) | 19.00 |
| Primer | Accession Number | Sequence (5’ to 3’) |
|---|---|---|
| PepT1-qPCR-F | JN088166 | TGCTCTTGTTGTGTTCATCG |
| PepT1-qPCR-R | CTCTCTCTTGGGGTATTGCTT | |
| CDX2-qPCR-F | KC748025 | TTTGTAACCGCACCTCC |
| CDX2-qPCR-R | AGTTCCTGGCCCATAAGT | |
| Sp1-qPCR-F | KY081668 | AGTGACCCCAGTAAGAAGAAGCA |
| Sp1-qPCR-R | CAAGTGTGCCCGCAGATG | |
| NF-κB-qPCR-F | KY129991 | GCGTCTATGCTTCCAGATTTACC |
| NF-κB-qPCR-R | ACTGCCACTGTTCTTGTTCACC | |
| β-Actin-F | M25013 | CCTTCTTGGGTATGGAGTCTTG |
| β-Actin-R | AGAGTATTTACGCTCAGGTGGG |
| Tributyrin Levels | IBW 1 (g) | FBW 2 (g) | SR 3 (%) | WGR 4 (%) | SGR 5 (%/d) |
|---|---|---|---|---|---|
| 0 g/kg | 43.20 ± 0.088 | 61.70 ± 3.217 a | 100.0 ± 0 | 42.82 ± 0.072 a | 0.64 ± 0.001 a |
| 0.5 g/kg | 43.30 ± 0.018 | 67.80 ± 1.096 b | 100.0 ± 0 | 56.58 ± 0.026 b | 0.80 ± 0 b |
| 1.0 g/kg | 43.20 ± 0.088 | 75.30 ± 16.29 c | 100.0 ± 0 | 74.31 ± 0.151 c | 0.99 ± 0.002 b |
| 1.5 g/kg | 43.60 ± 0.194 | 60.20 ± 4.278 a | 100.0 ± 0 | 38.07 ± 0.105 a | 0.58 ± 0.001 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.; Liu, N.; Cai, Y.; Yang, N.; Li, G.; Xiao, Y.; Zhou, X.; Cao, S.; Qu, F.; Tang, J.; et al. Effect of Tributyrin on Growth Performance and Pathway by which Tributyrin Regulates Oligopeptide Transporter 1 in Juvenile Grass Carp (Ctenopharyngodon idellus). Animals 2022, 12, 2498. https://doi.org/10.3390/ani12192498
He Z, Liu N, Cai Y, Yang N, Li G, Xiao Y, Zhou X, Cao S, Qu F, Tang J, et al. Effect of Tributyrin on Growth Performance and Pathway by which Tributyrin Regulates Oligopeptide Transporter 1 in Juvenile Grass Carp (Ctenopharyngodon idellus). Animals. 2022; 12(19):2498. https://doi.org/10.3390/ani12192498
Chicago/Turabian StyleHe, Zhimin, Na Liu, Yuyang Cai, Na Yang, Gen Li, Yang Xiao, Xiaomei Zhou, Shenping Cao, Fufa Qu, Jianzhou Tang, and et al. 2022. "Effect of Tributyrin on Growth Performance and Pathway by which Tributyrin Regulates Oligopeptide Transporter 1 in Juvenile Grass Carp (Ctenopharyngodon idellus)" Animals 12, no. 19: 2498. https://doi.org/10.3390/ani12192498
APA StyleHe, Z., Liu, N., Cai, Y., Yang, N., Li, G., Xiao, Y., Zhou, X., Cao, S., Qu, F., Tang, J., Liu, S., & Liu, Z. (2022). Effect of Tributyrin on Growth Performance and Pathway by which Tributyrin Regulates Oligopeptide Transporter 1 in Juvenile Grass Carp (Ctenopharyngodon idellus). Animals, 12(19), 2498. https://doi.org/10.3390/ani12192498