

Nutritional Metabolites as Biomarkers of Previous Feed Intake in European Rabbit (Oryctolagus cuniculus): Applications on Conservation

 ,
,  ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statement
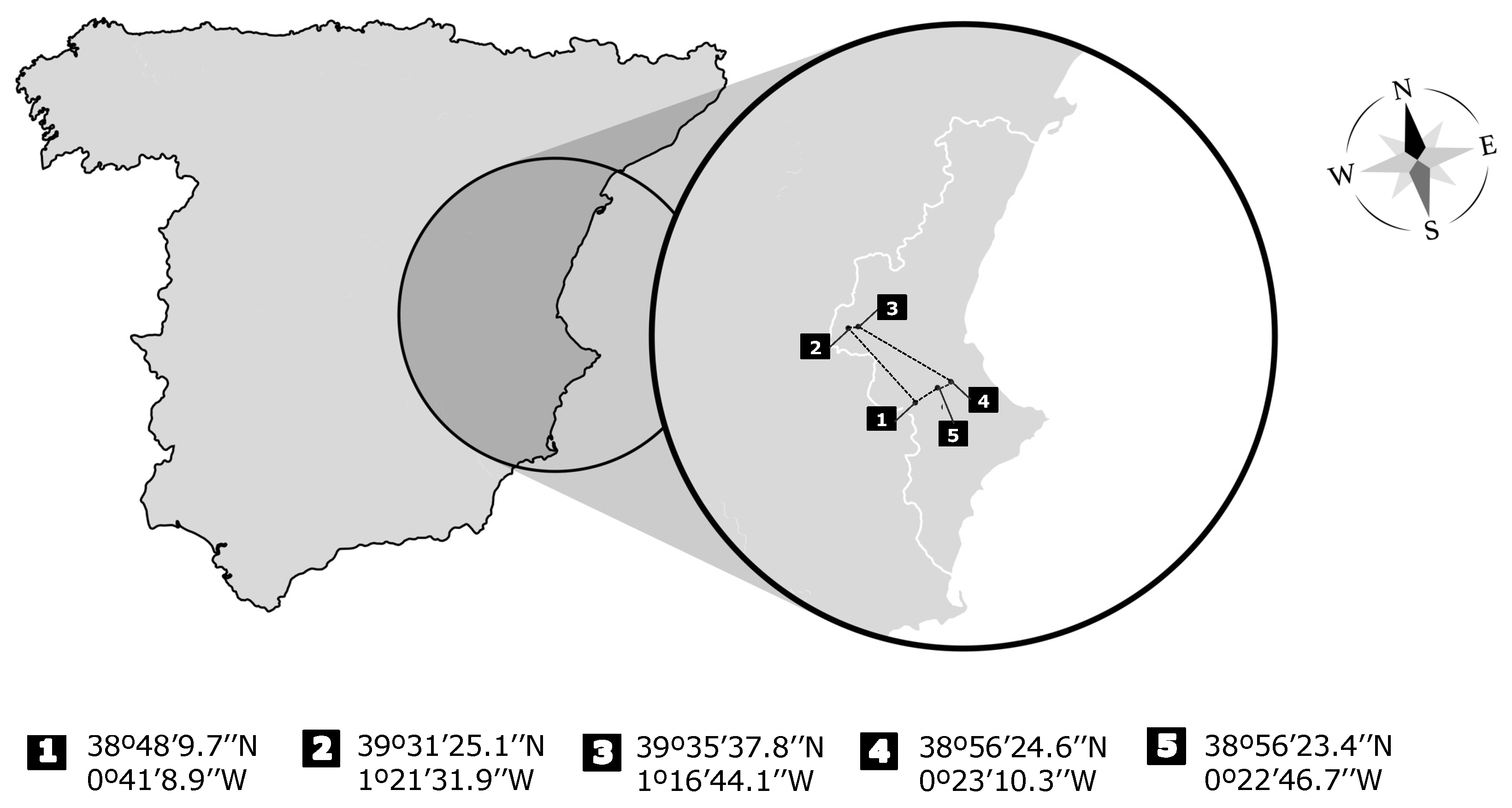
2.2. Animals and Sampling
2.3. Biochemical Analysis of Blood Nutritional Metabolites
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cortés-Avizanda, A.; Colomer, M.À.; Margalida, A.; Ceballos, O.; Donázar, J.A. Modeling the Consequences of the Demise and Potential Recovery of a Keystone-Species: Wild Rabbits and Avian Scavengers in Mediterranean Landscapes. Sci. Rep. 2015, 5, 17033. [Google Scholar] [CrossRef]
- Malo, J.E.; Jimenez, B.; Suarez, F. Seed Bank Build-up in Small Disturbances in a Mediterranean Pasture: The Contribution of Endozoochorous Dispersal by Rabbits. Ecography 1995, 18, 73–82. [Google Scholar] [CrossRef]
- Palomares, F.; Delibes, M.; Revilla, E.; Calzada, J.; Fedriani, J.M. Spatial Ecology of Iberian Lynx and Abundance of European Rabbits in Southwestern Spain. Wildl. Monogr. 2001, 148, 1–36. [Google Scholar]
- Osacar-Jimenez, J.J.; Lucientes-Curdi, J.; Calvete-Margolles, C. Abiotic Factors Influencing the Ecology of Wild Rabbit Fleas in North-Eastern Spain: Ecology of Wild Rabbit Fleas in Spain. Med. Vet. Entomol. 2001, 15, 157–166. [Google Scholar] [CrossRef]
- Revilla, E.; Palomares, F.; Fernández, N. Characteristics, Location and Selection of Diurnal Resting Dens by Eurasian Badgers (Meles Meles) in a Low Density Area. J. Zool. 2001, 255, 291–299. [Google Scholar] [CrossRef]
- Gálvez, L.A.; López-Pintor, J.; De Miguel, G.; Alonso, M.; Rueda, S.; Rebollo, S.; Gómez-Sal, A. Ecosystem Engineering Effects of European Rabbits in a Mediterranean Habitat. In Lagomorph Biology: Evolution, Ecology and Conservation; Springer: Berlin/Heidelberg, Germany, 2008; pp. 125–140. [Google Scholar]
- Malo, J.E.; Surez, F. Herbivorous Mammals as Seed Dispersers in a Mediterranean Dehesa. Oecologia 1995, 104, 246–255. [Google Scholar] [CrossRef]
- Valkama, J.; Korpimäki, E.; Arroyo, B.; Beja, P.; Bretagnolle, V.; Bro, E.; Kenward, R.; Mañosa, S.; Redpath, S.M.; Thirgood, S.; et al. Birds of Prey as Limiting Factors of Gamebird Populations in Europe: A Review. Biol. Rev. 2005, 80, 171–203. [Google Scholar] [CrossRef]
- Nowell, K.; Jackson, P. Wild Cats: Status Survey and Conservation Action Plan; IUCN Publications, The Burlington Press: Cambridge, UK, 1996. [Google Scholar]
- Delibes-Mateos, M.; Ferreras, P.; Villafuerte, R. Rabbit Populations and Game Management: The Situation after 15 Years of Rabbit Haemorrhagic Disease in Central-Southern Spain. Biodivers. Conserv. 2008, 17, 559–574. [Google Scholar] [CrossRef]
- Moreno, S.; Villafuerte, R. Traditional Management of Scrubland for the Conservation of Rabbits Oryctolagus Cuniculus and Their Predators in Doñana National Park, Spain. Biological. Conserv. 1995, 73, 81–85. [Google Scholar] [CrossRef]
- Villafuerte, R.; Calvete, C.; Gortázar, C.; Moreno, S. First Epizootic of Rabbit Hemorrhagic Disease in Free Living Populations of Oryctolagus Cuniculus at Doñana National Park, Spain. J. Wildl. Dis. 1994, 30, 176–179. [Google Scholar] [CrossRef]
- Villafuerte, R.; Delibes-Mateos, M. Oryctolagus Cuniculus. IUCN 2019. 2019. Available online: https://doi.org/10.2305/IUCN.UK.2019-3.RLTS.T41291A45189779.en (accessed on 27 August 2022).
- Aronson, J.; Floret, C.; Floćh, E.; Ovalle, C.; Pontanier, R. Restoration and Rehabilitation of Degraded Ecosystems in Arid and Semi-Arid Lands. I. A View from the South. Restor. Ecol. 1993, 1, 8–17. [Google Scholar] [CrossRef]
- Vogiatzakis, I.N.; Mannion, A.M.; Griffiths, G.H. Mediterranean Ecosystems: Problems and Tools for Conservation. Prog. Phys. Geogr. Earth Environ. 2006, 30, 175–200. [Google Scholar] [CrossRef]
- Llobat, L.; Marín-García, P.J. Application of Protein Nutrition in Natural Ecosystem Management for European Rabbit (Oryctolagus cuniculus) Conservation. Biodivers. Conserv. 2022, 31, 1435–1444. [Google Scholar] [CrossRef]
- Marín-García, P.J.; Llobat, L. What are the Keys to the Adaptive Success of European Wild Rabbit (Oryctolagus cuniculus) in the Iberian Peninsula? Animals 2021, 11, 2453. [Google Scholar] [CrossRef] [PubMed]
- Villafuerte, R.; Lazo, A.; Moreno, S. Influence of Food Abundance and Quality on Rabbit Fluctuations: Conservation and Management Implications in Doñana National Park (SW Spain). Rev. Ecol. 1997, 52, 345–356. [Google Scholar]
- Ulappa, A.C.; Kelsey, R.G.; Frye, G.G.; Rachlow, J.L.; Shipley, L.A.; Bond, L.; Pu, X.; Forbey, J.S. Plant Protein and Secondary Metabolites Influence Diet Selection in a Mammalian Specialist Herbivore. J. Mammal. 2014, 95, 834–842. [Google Scholar] [CrossRef]
- Calvete, C.; Villafuerte, R.; Lucientes, J.; Osacar, J.J. Effectiveness of Traditional Wild Rabbit Restocking in Spain. J. Zool. 1997, 241, 271–277. [Google Scholar] [CrossRef]
- Ferreira, C.; Alves, P.C. Influence of Habitat Management on the Abundance and Diet of Wild Rabbit (Oryctolagus Cuniculus Algirus) Populations in Mediterranean Ecosystems. Eur. J. Wildl. Res. 2009, 55, 487–496. [Google Scholar] [CrossRef]
- Whilde, J.; Martindale, M.Q.; Duffy, D.J. Precision Wildlife Medicine: Applications of the Human-Centred Precision Medicine Revolution to Species Conservation. Glob. Chang. Biol. 2017, 23, 1792–1805. [Google Scholar] [CrossRef]
- Chaousis, S.; Leusch, F.D.L.; van de Merwe, J.P. Charting a Path towards Non-Destructive Biomarkers in Threatened Wildlife: A Systematic Quantitative Literature Review. Environ. Pollut. 2018, 234, 59–70. [Google Scholar] [CrossRef]
- Rezzi, S.; Ramadan, Z.; Fay, L.B.; Kochhar, S. Nutritional Metabonomics: Applications and Perspectives. J. Proteome Res. 2007, 6, 513–525. [Google Scholar] [CrossRef]
- Beaulieu, M.; Costantini, D. Biomarkers of Oxidative Status: Missing Tools in Conservation Physiology. Conserv. Physiol. 2014, 2, cou014. [Google Scholar] [CrossRef]
- Beaulieu, M.; Thierry, A.-M.; Gonzalez-Acuna, D.; Polito, M.J. Integrating Oxidative Ecology into Conservation Physiology. Conserv. Physiol. 2013, 1, cot004. [Google Scholar] [CrossRef]
- Sylvie, G.; Marion, K.; Yvon, L.M.; Jean-Patrice, R.; Criscuolo, F. Of the Importance of Metabolic Phases in the Understanding of Oxidative Stress in Prolonged Fasting and Refeeding. Physiol. Biochem. Zool. 2012, 85, 415–420. [Google Scholar] [CrossRef]
- van de Crommenacker, J.; Komdeur, J.; Burke, T.; Richardson, D.S. Spatio-Temporal Variation in Territory Quality and Oxidative Status: A Natural Experiment in the Seychelles Warbler (Acrocephalus sechellensis): Territory Quality-Related Oxidative Costs in a Wild Passerine. J. Anim. Ecol. 2011, 80, 668–680. [Google Scholar] [CrossRef]
- Larsen, T.; Fernández, C. Enzymatic-Fluorometric Analyses for Glutamine, Glutamate and Free Amino Groups in Protein-Free Plasma and Milk. J. Dairy Res. 2017, 84, 32–35. [Google Scholar] [CrossRef]
- SAS. SAS/STAT ® 9.2 User’s Guide Version 9; SAS Institute Inc.: Cary, NC, USA, 2009. [Google Scholar]
- Calvete, C.; Angulo, E.; Estrada, R.; Moreno, S.; Villafuerte, R. Quarantine length and survival of translocated european wild rabbits. J. Wildl. Manag. 2005, 69, 1063–1072. [Google Scholar] [CrossRef]
- Bellier, R.; Gidenne, T.; Vernay, M.; Colin, M. In Vivo Study of Circadian Variations of the Cecal Fermentation Pattern in Postweaned and Adult Rabbits. J. Anim. Sci. 1995, 73, 128. [Google Scholar] [CrossRef]
- Prud’Hon, M.; Chérubin, M.; Goussopoulos, J.; Carles, Y. Évolution, au cours de la croissance, des caractéristiques de la consommation d’aliments solide et liquide du lapin domestique nourri ad libitum. Ann. Zootech. 1975, 24, 289–298. [Google Scholar] [CrossRef]
- de Blas, C.; Wiseman, J. (Eds.) Nutrition of the Rabbit, 2nd ed.; CABI: Wallingford, UK, 2010; ISBN 978-1-84593-669-3. [Google Scholar]
- Marín-García, P.J.; Ródenas, L.; Martínez-Paredes, E.; Cambra-López, M.; Blas, E.; Pascual, J.J. A Moderate Protein Diet Does Not Cover the Requirements of Growing Rabbits with High Growth Rate. Anim. Feed. Sci. Technol. 2020, 264, 114495. [Google Scholar] [CrossRef]
- Joseph, D.R. The relation of the weight of the contents of stomach and cecum to the body-weight in rabbits. J. Exp. Med. 1909, 11, 36–40. [Google Scholar] [CrossRef]
- Brecchia, G.; Bonanno, A.; Galeati, G.; Federici, C.; Maranesi, M.; Gobbetti, A.; Zerani, M.; Boiti, C. Hormonal and Metabolic Adaptation to Fasting: Effects on the Hypothalamic–Pituitary–Ovarian Axis and Reproductive Performance of Rabbit Does. Domest. Anim. Endocrinol. 2006, 31, 105–122. [Google Scholar] [CrossRef]
- Ebeid, T.; Tůmová, E.; Volek, Z. Effects of a one week intensive feed restriction in the growing rabbit: Part 1-Performance and blood biochemical parameters. In Proceedings of the 10th World Rabbit Congress, Sharm El-Sheikh, Egypt, 3–6 September 2012; pp. 607–611. [Google Scholar]
- van Harten, S.; Cardoso, L.A. Feed Restriction and Genetic Selection on the Expression and Activity of Metabolism Regulatory Enzymes in Rabbits. Animal 2010, 4, 1873–1883. [Google Scholar] [CrossRef]
- Roth-Maier, D.A.; Ott, H.; Roth, F.X.; Paulicks, B.R. Effects of the Level of Dietary Valine Supply on Amino Acids and Urea Concentration in Milk and Blood Plasma of Lactating Sows: Valine for Lactating Sows. J. Anim. Physiol. Anim. Nutr. 2004, 88, 39–45. [Google Scholar] [CrossRef]
- Nieto, R.; Barea, R.; Lara, L.; Palma-Granados, P.; Aguilera, J.F. Lysine Requirement Relative to Total Dietary Protein for Optimum Performance and Carcass Protein Deposition of Iberian Piglets. Anim. Feed. Sci. Technol. 2015, 206, 48–56. [Google Scholar] [CrossRef]
- Donsbough, A.L.; Powell, S.; Waguespack, A.; Bidner, T.D.; Southern, L.L. Uric Acid, Urea, and Ammonia Concentrations in Serum and Uric Acid Concentration in Excreta as Indicators of Amino Acid Utilization in Diets for Broilers. Poult. Sci. 2010, 89, 287–294. [Google Scholar] [CrossRef]
- Marín-García, P.J.; López-Luján, M.d.C.; Ródenas, L.; Martínez-Paredes, E.M.; Blas, E.; Pascual, J.J. Plasma Urea Nitrogen as an Indicator of Amino Acid Imbalance in Rabbit Diets. World Rabbit Sci. 2020, 28, 63. [Google Scholar] [CrossRef]
- Marín-García, P.J.; López-Luján, M.C.; Ródenas, L.; Martínez-Paredes, E.M.; Blas, E.; Pascual, J.J. Plasmatic Urea Nitrogen in Growing Rabbits with Different Combinations of Dietary Levels of Lysine, Sulphur Amino Acids and Threonine. Animals 2020, 10, 946. [Google Scholar] [CrossRef]
- Lu, J.; Shen, Y.; He, Z.; Dai, X.; Wang, D.; Zhang, J.; Li, H. Effects of a Short-Term Feed Restriction on Growth Performance, Blood Metabolites and Hepatic IGF-1 Levels in Growing Rabbits. World Rabbit Sci. 2017, 25, 233. [Google Scholar] [CrossRef]
- Parker, K.L. Advances in the Nutritional Ecology of Cervids at Different Scales. Écoscience 2003, 10, 395–411. [Google Scholar] [CrossRef]
- Raubenheimer, D.; Simpson, S.J.; Mayntz, D. Nutrition, Ecology and Nutritional Ecology: Toward an Integrated Framework. Funct. Ecol. 2009, 23, 4–16. [Google Scholar] [CrossRef]
- Marín-García, P.J.; López-Luján, M.C.; Ródenas, L.; Martínez-Paredes, E.; Cambra-López, M.; Blas, E.; Pascual, J.J. Do growing rabbits with a high growth rate require diets with high levels of essential amino acids? A choice-feeding trial. Animals 2021, 11, 824. [Google Scholar] [CrossRef] [PubMed]
 = normal;
= normal;  = lower) on the energetic metabolites: Glucose (a) and NEFA (b) of Oryctolagus cuniculus (n = 99). Least square means ± standard errors. NEFA: Non-esterified fatty acids.
= normal; = lower) on the energetic metabolites: Glucose (a) and NEFA (b) of Oryctolagus cuniculus (n = 99). Least square means ± standard errors. NEFA: Non-esterified fatty acids.
= lower) on the energetic metabolites: Glucose (a) and NEFA (b) of Oryctolagus cuniculus (n = 99). Least square means ± standard errors. NEFA: Non-esterified fatty acids.
= normal; = lower) on the energetic metabolites: Glucose (a) and NEFA (b) of Oryctolagus cuniculus (n = 99). Least square means ± standard errors. NEFA: Non-esterified fatty acids. = normal; = lower) on the protein metabolites (Plasmatic urea nitrogen (a), Albumin (b), Glutamate (c) and Total protein (d)) of Oryctolagus cuniculus (n = 99). Least square means ± standard errors. PUN: Plasmatic urea nitrogen.
= normal; = lower) on the protein metabolites (Plasmatic urea nitrogen (a), Albumin (b), Glutamate (c) and Total protein (d)) of Oryctolagus cuniculus (n = 99). Least square means ± standard errors. PUN: Plasmatic urea nitrogen.
= normal; = lower) on the protein metabolites (Plasmatic urea nitrogen (a), Albumin (b), Glutamate (c) and Total protein (d)) of Oryctolagus cuniculus (n = 99). Least square means ± standard errors. PUN: Plasmatic urea nitrogen.
= normal; = lower) on the protein metabolites (Plasmatic urea nitrogen (a), Albumin (b), Glutamate (c) and Total protein (d)) of Oryctolagus cuniculus (n = 99). Least square means ± standard errors. PUN: Plasmatic urea nitrogen.

{kind=link}
{kind=link}
{kind=link}
| Metabolites | Range | Values | Coefficient of Variation (%) |
|---|---|---|---|
| PUN | 12.95–73.00 | 24.1 ± 1.32 | 49.3 |
| NEFA | 105–3600 | 1172 ± 91.8 | 69.6 |
| Glucose | 2.16–30.5 | 9.07 ± 0.648 | 62.3 |
| Albumin | 25.2–57.1 | 40.9 ± 1.17 | 17.1 |
| Total protein | 40.9–71.3 | 56.3 ± 8.36 | 14.8 |
| Glutamate | 74–589 | 329 ± 22.1 | 37.5 |
| Gastric content weight (g) | 2.17–109.8 | 22.39 ± 2.50 | 59.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marín-García, P.J.; Llobat, L.; Rouco, C.; Aguayo-Adán, J.A.; Larsen, T.; Cambra-López, M.; Blas, E.; Pascual, J.J. Nutritional Metabolites as Biomarkers of Previous Feed Intake in European Rabbit (Oryctolagus cuniculus): Applications on Conservation. Animals 2022, 12, 2608. https://doi.org/10.3390/ani12192608
Marín-García PJ, Llobat L, Rouco C, Aguayo-Adán JA, Larsen T, Cambra-López M, Blas E, Pascual JJ. Nutritional Metabolites as Biomarkers of Previous Feed Intake in European Rabbit (Oryctolagus cuniculus): Applications on Conservation. Animals. 2022; 12(19):2608. https://doi.org/10.3390/ani12192608
Chicago/Turabian StyleMarín-García, Pablo Jesús, Lola Llobat, Carlos Rouco, Juan Antonio Aguayo-Adán, Torben Larsen, María Cambra-López, Enrique Blas, and Juan José Pascual. 2022. "Nutritional Metabolites as Biomarkers of Previous Feed Intake in European Rabbit (Oryctolagus cuniculus): Applications on Conservation" Animals 12, no. 19: 2608. https://doi.org/10.3390/ani12192608
APA StyleMarín-García, P. J., Llobat, L., Rouco, C., Aguayo-Adán, J. A., Larsen, T., Cambra-López, M., Blas, E., & Pascual, J. J. (2022). Nutritional Metabolites as Biomarkers of Previous Feed Intake in European Rabbit (Oryctolagus cuniculus): Applications on Conservation. Animals, 12(19), 2608. https://doi.org/10.3390/ani12192608