
Laboratory and Field Assessments of Oral Vibrio Vaccine Indicate the Potential for Protection against Vibriosis in Cultured Marine Fishes

 , ,
, ,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain Selection
Preparation of Formalin-Killed Cells (FKC) of Vibrio Harveyi Strain VH1
2.2. Preparation of Feed Vaccine
2.3. Fish Vaccination and Sample Collection in Laboratory Trials
2.4. Determination of Specific Serum Antibody Production
2.5. Experimental Challenge of Vibrio sp.
2.6. Field Trial
2.6.1. Bacterial Isolation and Identification
2.6.2. Water Quality
2.7. Statistical Analysis
3. Results
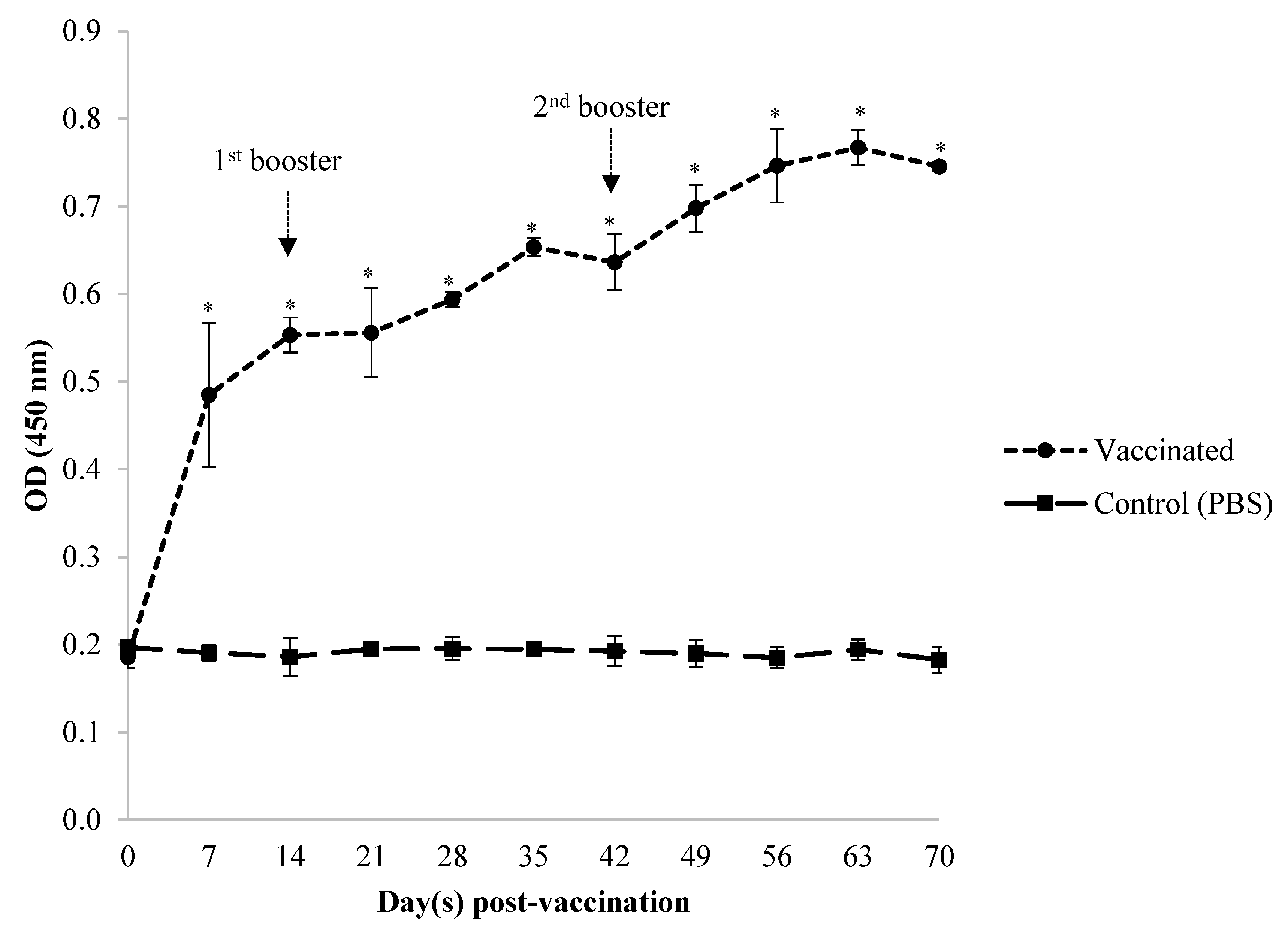
3.1. Serum Systemic Antibody Response
3.2. Protection against Pathogenic Challenge
3.3. Field Study
3.3.1. Weight Gain Effect and Feed Efficiency
3.3.2. Rate of Survival
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ina-Salwany, M.Y.; Al-Saari, N.; Mohamad, A.; Mursidi, F.A.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in Fish: A Review on Disease Development and Prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.; Suprapto, H.; Sulmartiwi, L.; Sudarno. Bacteria associated with mass mortality of hybrid grouper Epinephelus sp. in East Java Province Indonesia. Int. J. Fish. Aquat. Stud. 2016, 4, 439–441. [Google Scholar]
- Nor, N.M.; Yazid, S.H.M.; Daud, H.M.; Azmai, M.N.A.; Mohamad, N. Costs of management practices of Asian seabass (Lates calcarifer Bloch, 1790) cage culture in Malaysia using stochastic model that includes uncertainty in mortality. Aquaculture 2019, 510, 347–352. [Google Scholar] [CrossRef]
- Mohamad, N.; Amal, M.N.A.; Yasin, I.S.M.; Saad, M.Z.; Nasruddin, N.S.; Al-Saari, N.; Mino, S.; Sawabe, T. Vibriosis in cultured marine fishes: A review. Aquaculture 2019, 512, 734289. [Google Scholar] [CrossRef]
- Mohamad, N.; Roseli, F.A.M.; Azmai, M.N.A.; Saad, M.Z.; Yasin, I.S.M.; Zulkiply, N.A.; Nasruddin, N.S. Natural Concurrent Infection of Vibrio harveyi and V. alginolyticus in Cultured Hybrid Groupers in Malaysia. J. Aquat. Anim. Health 2018, 31, 88–96. [Google Scholar] [CrossRef]
- Amalina, N.Z.; Santha, S.; Zulperi, D.; Amal, M.N.A.; Yusof, M.T.; Zamri-Saad, M.; Ina-Salwany, Y. Prevalence, antimicrobial susceptibility and plasmid profiling of Vibrio spp. isolated from cultured groupers in Peninsular Malaysia. BMC Microbiol. 2019, 19, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Ge, M.; Zheng, X.; Tao, Z.; Zhou, S.; Wang, G. Investigation of Vibrio alginolyticus, V. harveyi, and V. parahaemolyticus in large yellow croaker, Pseudosciaena crocea (Richardson) reared in Xiangshan Bay, China. Aquac. Rep. 2016, 3, 220–224. [Google Scholar] [CrossRef] [Green Version]
- Bao, P.; Sun, X.; Liu, Q.; Zhang, Y.; Liu, X. Synergistic effect of a combined live Vibrio anguillarum and Edwardsiella piscicida vaccine in turbot. Fish Shellfish. Immunol. 2019, 88, 84–90. [Google Scholar] [CrossRef]
- Zhang, X.-H.; He, X.; Austin, B. Vibrio harveyi: A serious pathogen of fish and invertebrates in mariculture. Mar. Life Sci. Technol. 2020, 2, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Sudhagar, A.; Ferosekhan, S.; Linga Prabu, D. Fish vaccination: A health management tool for Aquaculture. Aquafind Aquac. Fish Data-Base 2016, 1–5. Available online: http://aquafind.com/articles/Vaccination.php (accessed on 1 January 2021).
- Colquhoun, D.J.; Lillehaug, A. Vaccination against Vibriosis. In Fish Vaccination; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 172–184. [Google Scholar]
- Li, C.; Ye, Z.; Wen, L.; Chen, R.; Tian, L.; Zhao, F.; Pan, J. Identification of a novel vaccine candidate by immunogenic screening of Vibrio parahaemolyticus outer membrane proteins. Vaccine 2014, 32, 6115–6121. [Google Scholar] [CrossRef]
- Pang, H.; Chen, L.; Hoare, R.; Huang, Y.; Wu, Z.; Jian, J. Identification of DLD, by immunoproteomic analysis and evaluation as a potential vaccine antigen against three Vibrio species in Epinephelus coioides. Vaccine 2016, 34, 1225–1231. [Google Scholar] [CrossRef]
- Lun, J.; Xia, C.; Yuan, C.; Zhang, Y.; Zhong, M.; Huang, T.; Hu, Z. The outer membrane protein, LamB (maltoporin), is a versatile vaccine candidate among the Vibrio species. Vaccine 2014, 32, 809–815. [Google Scholar] [CrossRef]
- Mursidi, F. Antigenic Analysis of Outer Membrane Protein of Vibrio Species and Development of Versatile Recombinant vhDnaJ Vaccine against Vibriosis; Universiti Putra Malaysia: Serdang, Malaysia, 2018. [Google Scholar]
- Shoemaker, C.A.; LaFrentz, B.R.; Klesius, P.H. Bivalent vaccination of sex reversed hybrid tilapia against Streptococcus iniae and Vibrio vulnificus. Aquaculture 2012, 354–355, 45–49. [Google Scholar] [CrossRef]
- Soltani, M.; Lymbery, A.; Song, S.K.; Shekarabi, P.H. Adjuvant effects of medicinal herbs and probiotics for fish vaccines. Rev. Aquac. 2018, 11, 1325–1341. [Google Scholar] [CrossRef]
- Ismail, M.S.; Siti-Zahrah, A.; Syafiq, M.R.M.; Amal, M.N.A.; Firdaus-Nawi, M.; Zamri-Saad, M. Feed-based vaccination regime against streptococcosis in red tilapia, Oreochromis niloticus × Oreochromis mossambicus. BMC Veter-Res. 2016, 12, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoare, R.; Jung, S.-J.; Ngo, T.P.H.; Bartie, K.; Bailey, J.; Thompson, K.; Adams, A. Efficacy and safety of a non-mineral oil adjuvanted injectable vaccine for the protection of Atlantic salmon (Salmo salar L.) against Flavobacterium psychrophilum. Fish Shellfish. Immunol. 2019, 85, 44–51. [Google Scholar] [CrossRef]
- Mohamad, A.; Zamri-Saad, M.; Amal, M.; Al-Saari, N.; Monir, S.; Chin, Y.; Yasin, I.-S.M. Vaccine Efficacy of a Newly Developed Feed-Based Whole-Cell Polyvalent Vaccine against Vibriosis, Streptococcosis and Motile Aeromonad Septicemia in Asian Seabass. Lates calcarifer. Vaccines 2021, 9, 368. [Google Scholar] [CrossRef]
- Wohlsen, T.; Bates, J.; Vesey, G.; Robinson, W.; Katouli, M. Evaluation of the methods for enumerating coliform bacteria from water samples using precise reference standards. Lett. Appl. Microbiol. 2006, 42, 350–356. [Google Scholar] [CrossRef] [Green Version]
- Sulaiman, M.A.; Kamarudin, M.S.; Romano, N.; Syukri, F. Effects of increasing dietary carbohydrate level on feed utilisation, body composition, liver glycogen, and intestinal short chain fatty acids of hybrid lemon fin barb (Barbonymus gonionotus ♀ × Hypsibarbus wetmorei male ♂). Aquac. Rep. 2020, 16, 100250. [Google Scholar] [CrossRef]
- Firdaus-Nawi, M.; Yusoff, S.M.; Yusof, H.; Abdullah, S.-Z.; Zamri-Saad, M. Efficacy of feed-based adjuvant vaccine against Streptococcus agalactiae in Oreochromis spp. in Malaysia. Aquac. Res. 2012, 45, 87–96. [Google Scholar] [CrossRef]
- Aris, A.M.; Saad, M.-Z.; Daud, H.M.; Yusof, M.T.; Yasin, I.S.M. Vibrio harveyi protease deletion mutant as a live attenuated vaccine candidate against vibriosis and transcriptome profiling following vaccination for Epinephelus fuscoguttatus. Aquac. Int. 2019, 27, 125–140. [Google Scholar] [CrossRef]
- Haldar, S.C.A.S. Vibrio Related Diseases in Aquaculture and Development of Rapid and Accurate Identification Methods. J. Mar. Sci. Res. Dev. 2012, s1, 2–7. [Google Scholar] [CrossRef]
- Tookwinas, S. Review of grow-out techniques under tropical conditions: Experience of Thailand on Seabass. Adv. Trop. Aquac. 1989, 9, 737–750. [Google Scholar]
- Plaza, N.; Castillo, D.; Pérez-Reytor, D.; Higuera, G.; García, K.; Bastías, R. Bacteriophages in the control of pathogenic Vibrios. Electron. J. Biotechnol. 2018, 31, 24–33. [Google Scholar] [CrossRef]
- Bin Park, S.; Nho, S.W.; Bin Jang, H.; Cha, I.S.; Kim, M.S.; Lee, W.-J.; Jung, T.S. Development of three-valent vaccine against streptococcal infections in olive flounder, Paralichthys olivaceus. Aquaculture 2016, 461, 25–31. [Google Scholar] [CrossRef]
- Mutoloki, S.; Munang’Andu, H.M.; Evensen, Ø. Oral Vaccination of Fish—Antigen Preparations, Uptake, and Immune Induction. Front. Immunol. 2015, 6, 519. [Google Scholar] [CrossRef] [Green Version]
- Pereira, G.D.V.; Da Silva, B.C.; Vieira, F.D.N.; Seiffert, W.Q.; Ushizima, T.T.; Mouriño, J.L.P.; Martins, M.L. Vaccination strategies with oral booster for surubim hybrid (Pseudoplatystoma corruscans × P. reticulatum) against haemorrhagic septicaemia. Aquac. Res. 2015, 46, 1831–1841. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, C.; Chi, H.; Liu, X.; Gong, H.; Xie, A.; Zheng, W.; Chen, J.; Zhang, N.; Wu, Y. Early immune response in large yellow croaker (Larimichthys crocea) after immunization with oral vaccine. Mol. Cell. Probes 2021, 56, 101708. [Google Scholar] [CrossRef]
- Han, B.; Xu, K.; Liu, Z.; Ge, W.; Shao, S.; Li, P.; Yan, N.; Li, X.; Zhang, Z. Oral yeast-based DNA vaccine confers effective protection from Aeromonas hydrophila infection on Carassius auratus. Fish Shellfish. Immunol. 2019, 84, 948–954. [Google Scholar] [CrossRef]
- Giri, S.S.; Chi, C.; Jun, J.W.; Park, S.C. Use of bacterial subcellular components as immunostimulants in fish aquaculture. Rev. Aquac. 2016, 10, 474–492. [Google Scholar] [CrossRef]
- Fraser, T.; Hansen, T.; Mayer, I.; Skjæraasen, J.E.; Glover, K.A.; Sambraus, F.; Fjelldal, P.G. The effect of triploidy on vaccine side-effects in Atlantic salmon. Aquaculture 2014, 433, 481–490. [Google Scholar] [CrossRef]
- Amar, E.C.; Faisan, J.P.; Gapasin, R.S. Field efficacy evaluation of a formalin-inactivated white spot syndrome virus (WSSV) vaccine for the preventive management of WSSV infection in shrimp grow-out ponds. Aquaculture 2021, 531, 735907. [Google Scholar] [CrossRef]
- Effect of incorporating different concentrations of palm oil as adjuvant in fish vaccine. Int. J. Biosci. (IJB) 2018, 12, 35–41. [CrossRef]
- Wanasawaeng, W.; Tawatsin, A.; Sasipreeyajan, J.; Poomvises, P.; Chansiripornchai, N. Development of inactivated New-castle disease vaccine using palm oil as an adjuvant. Thai J. Vet. Med. 2009, 39, 9–16. [Google Scholar]
- Roslindawani, M.N.; Syafiqah, A.S.; Jesse, F.F.A.; Effendy, A.W.; Zamri-Saad, M. Recombinant Caseous Lymphadenitis Vaccine with Palm Oil as Adjuvant Enhances the Humoral and Cell-Mediated Immune Responses in Rat Model. J. Anim. Health Prod. 2016, 4, 22–25. [Google Scholar] [CrossRef]
- Monir, S.; Yusoff, S.B.M.; Zulperi, Z.B.M.; Abu Hassim, H.B.; Mohamad, A.; Ngoo, M.S.B.M.H.; Ina-Salwany, Y. Haemato-immunological responses and effectiveness of feed-based bivalent vaccine against Streptococcus iniae and Aeromonas hydrophila infections in hybrid red tilapia (Oreochromis mossambicus × O. niloticus). BMC Veter-Res. 2020, 16, 1–14. [Google Scholar] [CrossRef]
- Chin, Y.K.; Al-Saari, N.; Zulperi, Z.; Mohd-Aris, A.; Salleh, A.; Silvaraj, S.; Mohamad, A.; Lee, J.; Zamri-Saad, M.; Ina-Salwany, Y. Efficacy of bath vaccination with a live attenuated Vibrio harveyi against vibriosis in Asian seabass fingerling, Lates calcarifer. Aquac. Res. 2019, 51, 389–399. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Organ of Isolation | Host Species |
|---|---|---|
| Vibrio harveyi strain VH1 | Skin, kidney, liver | Epinephelus fuscoguttus |
| Vibrio alginolyticus strain VA2 | Liver, kidney | Epinephelus fuscoguttus |
| Vibrio parahaemolyticus strain VPK1 | Liver | Epinephelus fuscoguttus |
| Primers | Primer Sequence (5′-3′) | Tm (°C) | Expected Size (bp) | Reference |
|---|---|---|---|---|
| gyrB_F | GAGAACCCGACAGAAGCGAAG | 50.0 | 314 | [25] |
| gyrB_R | CCTAGTGCGGTGATCAGTGTTG |
| Parameter | Mean ± SD | * Range |
|---|---|---|
| Ammonia–nitrogen (mg/L) | 0.03 ± 0.02 | less than 0.02 |
| pH (1–14) | 8.00 ± 0.15 | 7.5–8.3 |
| Temperature (°C) | 30.45 ± 0.62 | 26–32 |
| Salinity (ppt) | 30.12 ± 1.33 | 10–31 |
| Dissolved oxygen(mg/L) | 4.71 ± 0.39 | 4.0–8.0 |
| Group | Bacterial Challenge | Number of Challenged Fish | Mortality (%) | RPS (%) |
|---|---|---|---|---|
| Control (unvaccinated) | PBS | 20 | 0 ± 0.0 | - |
| Vibrio harveyi (Vh) | 20 | 100 ± 0.0 | - | |
| V. alginolyticus (Va) | 20 | 100 ± 0.0 | - | |
| V. parahaemolyticus (Vp) | 20 | 100 ± 0.0 | - | |
| Vh-Vp-Va | 20 | 100 ± 0.0 | - | |
| Vaccinated | PBS | 20 | 0 ± 0.0 | - |
| Vibrio harveyi (Vh) | 20 | 25 ± 7.1 | 75 | |
| V. alginolyticus (Va) | 20 | 30 ± 0.0 | 70 | |
| V. parahaemolyticus (Vp) | 20 | 15 ± 7.1 | 85 | |
| Vh-Vp-Va | 20 | 30 ± 0.0 | 70 |
| Initial weight (g) | Control | 31.00 ± 0.97 |
| Vaccinated | 30.73 ± 0.92 | |
| Final weight (g) | Control | 208 ± 5.54 |
| Vaccinated | 248 ± 9.32 | |
| Average weight gain | Control | 177.3 g/fish |
| Vaccinated | 217.3 g/fish | |
| Survival (%) | Control | 65.1 ± 0.14% |
| Vaccinated | 79.75 ± 0.07% | |
| Total amount of feed given (kg) | Control | 688.8 ± 13.22 kg |
| Vaccinated | 890.4 ± 18.99 kg | |
| Feed efficiency | Control | 7.39 ± 0.08 g feed/g growth |
| Vaccinated | 3.10 ± 0.09 g feed/g growth |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamad, A.; Mursidi, F.-A.; Zamri-Saad, M.; Amal, M.N.A.; Annas, S.; Monir, M.S.; Loqman, M.; Hairudin, F.; Al-saari, N.; Ina-Salwany, M.Y. Laboratory and Field Assessments of Oral Vibrio Vaccine Indicate the Potential for Protection against Vibriosis in Cultured Marine Fishes. Animals 2022, 12, 133. https://doi.org/10.3390/ani12020133
Mohamad A, Mursidi F-A, Zamri-Saad M, Amal MNA, Annas S, Monir MS, Loqman M, Hairudin F, Al-saari N, Ina-Salwany MY. Laboratory and Field Assessments of Oral Vibrio Vaccine Indicate the Potential for Protection against Vibriosis in Cultured Marine Fishes. Animals. 2022; 12(2):133. https://doi.org/10.3390/ani12020133
Chicago/Turabian StyleMohamad, Aslah, Fathin-Amirah Mursidi, Mohd Zamri-Saad, Mohammad Noor Azmai Amal, Salleh Annas, Md Shirajum Monir, Mohd Loqman, Fahmie Hairudin, Nurhidayu Al-saari, and Md Yasin Ina-Salwany. 2022. "Laboratory and Field Assessments of Oral Vibrio Vaccine Indicate the Potential for Protection against Vibriosis in Cultured Marine Fishes" Animals 12, no. 2: 133. https://doi.org/10.3390/ani12020133
APA StyleMohamad, A., Mursidi, F.-A., Zamri-Saad, M., Amal, M. N. A., Annas, S., Monir, M. S., Loqman, M., Hairudin, F., Al-saari, N., & Ina-Salwany, M. Y. (2022). Laboratory and Field Assessments of Oral Vibrio Vaccine Indicate the Potential for Protection against Vibriosis in Cultured Marine Fishes. Animals, 12(2), 133. https://doi.org/10.3390/ani12020133