
Detection and Molecular Characterization of a Novel Species of Circovirus in a Tawny Owl (Strix aluco) in Southern Italy


 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. PCR Assays
2.3. Phylogenetic and Genomic Analyses
3. Results
3.1. PCR Assays
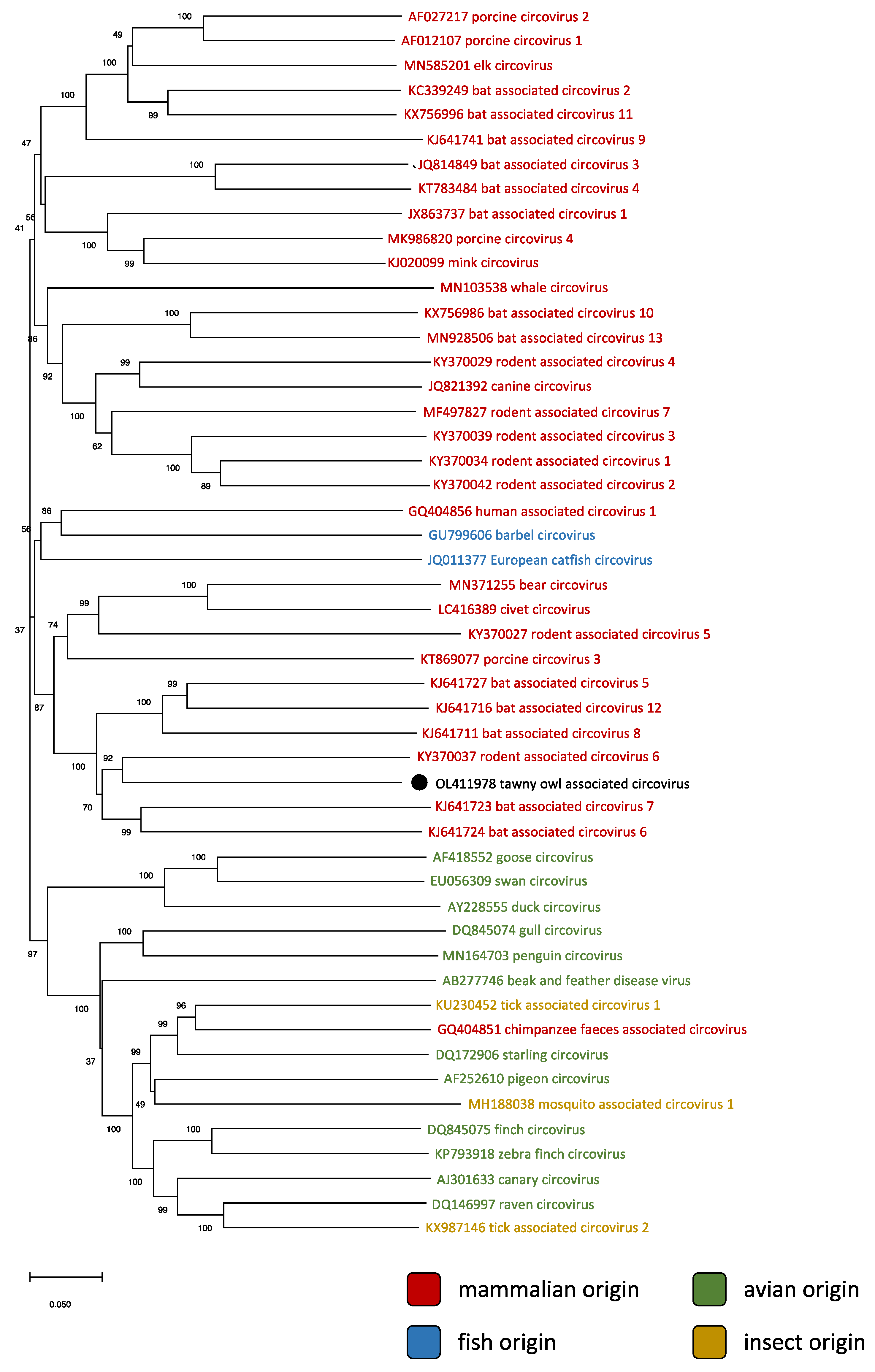
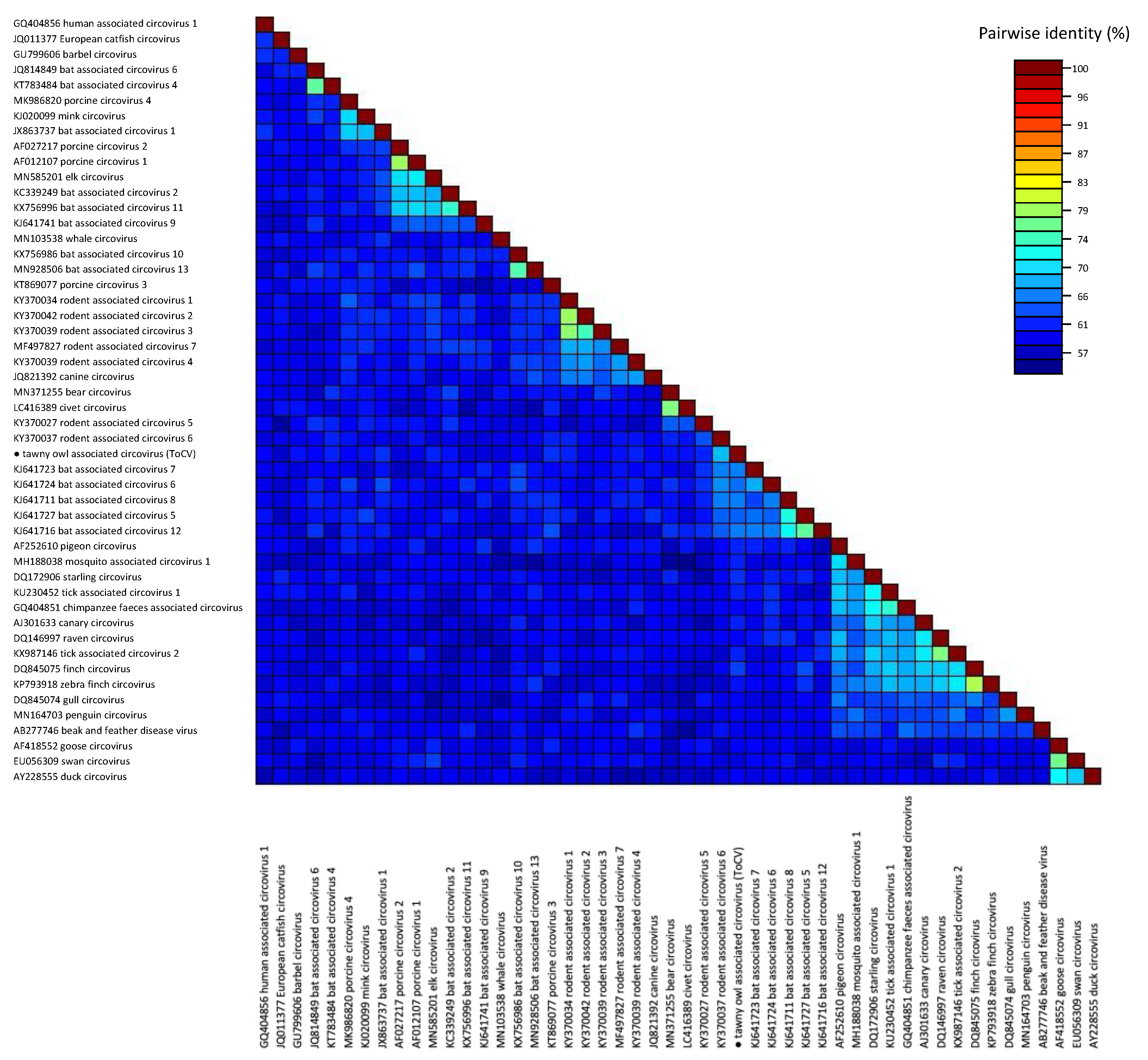
3.2. Phylogenetic and Genomic Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Plyusnin, I.; Kant, R.; Jääskeläinen, A.J.; Sironen, T.; Holm, L.; Vapalahti, O.; Smura, T. Novel NGS pipeline for virus discovery from a wide spectrum of hosts and sample types. Virus Evol. 2020, 6, veaa091. [Google Scholar] [CrossRef] [PubMed]
- Dance, A. Beyond coronavirus: The virus discoveries transforming biology. Nature 2021, 595, 22–25. [Google Scholar] [CrossRef]
- Zhao, L.; Rosario, K.; Breitbart, M.; Duffy, S. Eukaryotic Circular Rep-Encoding Single-Stranded DNA (CRESS DNA) Viruses: Ubiquitous Viruses with Small Genomes and a Diverse Host Range. Adv. Virus. Res. 2019, 103, 71–133. [Google Scholar] [PubMed]
- Shulman, L.; Davidson, I. Viruses with Circular Single-Stranded DNA Genomes Are Everywhere! Annu. Rev. Virol. 2017, 4, 159–180. [Google Scholar] [CrossRef]
- Kazlauskas, D.; Varsani, A.; Koonin, E.V.; Krupovic, M. Multiple origins of prokaryotic and eukaryotic single-stranded DNA viruses from bacterial and archaeal plasmids. Nat. Commun. 2019, 10, 3425. [Google Scholar] [CrossRef] [Green Version]
- Rosario, K.; Duffy, S.; Breitbart, M. Diverse circovirus-like genome architectures revealed by environmental metagenomics. J. Gen. Virol. 2009, 90, 2418–2424. [Google Scholar] [CrossRef]
- Van Regenmortel, M.H.V.; Fauquet, C.M.; Bishop, D.H.L.; Carstens, E.B.; Estes, M.K.; Lemon, S.M.; Maniloff, J.; Mayo, M.A.; McGeoch, D.J.; Pringle, C.R.; et al. Virus Taxonomy. Seventh Report of the International Committee on Taxonomy of Viruses; Academic Press: San Diego, CA, USA, 2000; p. 1162. [Google Scholar]
- Biagini, P.; Bendinelli, M.; Hino, S.; Kakkola, L.; Mankertz, A.; Niel, C.; Okamoto, H.; Raidal, S.; Teo, C.G.; Todd, D. Family Circoviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Academic Press: London, UK, 2012; pp. 343–349. [Google Scholar]
- Virus Taxonomy. Available online: https://talk.ictvonline.org/ictv-reports (accessed on 6 November 2021).
- Rosario, K.; Breitbart, M.; Harrach, B.; Segalés, J.; Delwart, E.; Biagini, P.; Varsani, A. Revisiting the taxonomy of the family Circoviridae: Establishment of the genus Cyclovirus and removal of the genus Gyrovirus. Arch. Virol. 2017, 162, 1447–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Kapoor, A.; Slikas, B.; Bamidele, O.S.; Wang, C.; Shaukat, S.; Alam Masroor, M.; Wilson, M.L.; Ndjango, J.-B.N.; Peeters, M.; et al. Multiple diverse circoviruses infect farm animals and are commonly found in human and chimpanzee feces. J. Virol. 2010, 84, 1674–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, D. Circoviruses: Immunosuppressive threats to avian species: A review. Avian Pathol. 2000, 29, 373–394. [Google Scholar] [CrossRef] [PubMed]
- Cruz, T.F.; Araujo, J.P., Jr. Cultivation of PCV2 in swine testicle cells using the shell vial technique and monitoring of viral replication by qPCR and RT-qPCR. J. Virol. Methods 2014, 196, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Nath, B.K.; Das, S.; Roby, J.A.; Sarker, S.; Luque, D.; Raidal, S.R.; Forwood, J.K. Structural Perspectives of Beak and Feather Disease Virus and Porcine Circovirus Proteins. Viral Immunol. 2021, 34, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Todd, D.; Gortázar, C. Circovirus Infections. In Infectious Diseases of Wild Mammals and Birds in Europe; Gavier-Widén, D., Duff, P.J., Meredith, A., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2012; pp. 67–72. [Google Scholar]
- Todd, D. Avian circovirus diseases: Lessons for the study of PMWS. Vet. Microbiol. 2004, 98, 169–174. [Google Scholar] [CrossRef]
- Stewart, M.E.; Perry, R.; Raidal, S.R. Identification of a novel circovirus in Australian ravens (Corvus coronoides) with feather disease. Avian Pathol. 2006, 35, 86–92. [Google Scholar] [CrossRef]
- Rinder, M.; Schmitz, A.; Peschel, A.; Wörle, B.; Gerlach, H.; Korbel, R. Molecular characterization of a recently identified circovirus in zebra finches (Taeniopygia guttata) associated with immunosuppression and opportunistic infections. Avian Pathol. 2017, 46, 106–116. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.T.; Kang, M.; Jang, H.K. Pathogenesis of duck circovirus genotype 1 in experimentally infected Pekin ducks. Poult. Sci. 2018, 97, 3050–3057. [Google Scholar] [CrossRef]
- Sheykhi, A.; Sheikhi, N.; Charkhkar, S.; Brujeni, G.N. Detection and characterization of circovirus in canary flocks. Avian Dis. 2018, 62, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Malmberg, J.L.; White, L.A.; VandeWoude, S. Bioaccumulation of pathogen exposure in top predators. Trends Ecol. Evol. 2021, 36, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar]
- ORF Finder. Available online: https://www.ncbi.nlm.nih.gov/orffinder/ (accessed on 8 November 2021).
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, F.A.; Portela, S.J.; Nogueira, T.; Carvalho, C.L.; de Sousa, R.; Duarte, M.D. Harmless or Threatening? Interpreting the Results of Molecular Diagnosis in the Context of Virus-Host Relationships. Front. Microbiol. 2021, 12, 1257. [Google Scholar] [CrossRef] [PubMed]
- Ilyina, T.V.; Koonin, E.V. Conserved sequence motifs in the initiator proteins for rolling circle DNA replication encoded by diverse replicons from eubacteria, eucaryotes and archaebacteria. Nucleic Acids Res. 1992, 20, 3279–3285. [Google Scholar] [CrossRef] [PubMed]
- Rosario, K.; Duffy, S.; Breitbart, M. A field guide to eukaryotic circular single-stranded DNA viruses: Insights gained from metagenomics. Arch. Virol. 2012, 157, 1851–1871. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Koonin, E.V. Helicases: Amino acid sequence comparisons and structure-function relationships. Curr. Opin. Struct. Biol. 1993, 3, 419–429. [Google Scholar] [CrossRef]
- Stothard, P.; Grant, J.R.; Van Domselaar, G. Visualizing and comparing circular genomes using the CGView family of tools. Brief. Bioinform. 2019, 20, 1576–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, S.; Terrón, M.C.; Khandokar, Y.; Aragão, D.; Hardy, J.M.; Radjainia, M.; Jiménez-Zaragoza, M.; De Pablo, P.J.; Coulibaly, F.; Luque, D.; et al. Structural insights into the assembly and regulation of distinct viral capsid complexes. Nat. Commun. 2016, 7, 13014. [Google Scholar] [CrossRef] [PubMed]
- Heath, L.; Williamson, A.-L.; Rybicki, E.P. The Capsid Protein of Beak and Feather Disease Virus Binds to the Viral DNA and Is Responsible for Transporting the Replication-Associated Protein into the Nucleus. J. Virol. 2006, 80, 7219–7225. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.K.; Hsiao, C.; Lo, A.R.; Wang, C.Y. Characterization of the nuclear localization sequence of beak and feather disease virus capsid proteins and their assembly into virus-like particles. Virus Res. 2020, 289, 198144. [Google Scholar] [CrossRef] [PubMed]
- Steinfeldt, T.; Finsterbusch, T.; Mankertz, A. Demonstration of nicking/joining activity at the origin of dna replication associated with the rep and rep’ proteins of porcine circovirus type 1. J. Virol. 2006, 80, 6225–6234. [Google Scholar] [CrossRef] [Green Version]
- Mankertz, A.; Hattermann, K.; Ehlers, B.; Soike, D. Cloningand sequencing of columbid circovirus (CoCV), a new circovirus from pigeons. Arch. Virol. 2000, 145, 2469–2479. [Google Scholar] [CrossRef] [PubMed]
- Phenix, K.V.; Weston, J.H.; Ypelaar, I.; Lavazza, A.; Smyth, J.A.; Todd, D.; Wilcox, G.E.; Raidal, S.R. Nucleotide sequence analysis of a novel circovirus of canaries and its relationship to other members of the genus Circovirus of the family Circoviridae. J. Gen. Virol. 2001, 82, 2805–2809. [Google Scholar] [CrossRef] [PubMed]
- Todd, D.; Weston, J.H.; Soike, D.; Smyth, J.A. Genome Sequence Determinations and Analyses of Novel Circoviruses from Goose and Pigeon. Virology 2001, 286, 354–362. [Google Scholar] [CrossRef]
- Wu, Z.; Lu, L.; Du, J.; Yang, L.; Ren, X.; Liu, B.; Jiang, J.; Yang, J.; Dong, J.; Sun, L.; et al. Comparative analysis of rodent and small mammal viromes to better understand the wildlife origin of emerging infectious diseases. Microbiome 2018, 6, 178. [Google Scholar] [CrossRef]
- Stenzel, T.; Dziewulska, D.; Tykałowski, B.; Koncicki, A. The Clinical Infection with Pigeon Circovirus (PiCV) Leads to Lymphocyte B Apoptosis But Has No Effect on Lymphocyte T Subpopulation. Pathogens 2020, 9, 632. [Google Scholar] [CrossRef]
- Schubbert, R.; Renz, D.; Schmitz, B.; Doerfler, W. Foreign (M13) DNA ingested by mice reaches peripheral leukocytes, spleen, and liver via the intestinal wall mucosa and can be covalently linked to mouse DNA. Proc. Natl. Acad. Sci. USA 1997, 94, 961–966. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| rep | cap | ||
|---|---|---|---|
| p-Distance | Species | p-Distance | Species |
| 0.270 | KJ370037 rodent associated circovirus 6 | 0.400 | KJ641716 bat associated circovirus 12 |
| 0.361 | KJ641727 bat associated circovirus 5 | 0.422 | KY370037 rodent associated circovirus 6 |
| 0.363 | KJ641723 bat associated circovirus 7 | 0.422 | KJ641724 bat associated circovirus 6 |
| 0.365 | KJ641711 bat associated circovirus 8 | 0.437 | JQ814849 bat associated circovirus 3 |
| 0.373 | KJ641724 bat associated circovirus 6 | 0.437 | KX756996 bat associated circovirus 11 |
| 0.438 | AB277746 beak and feather disease virus | 0.489 | AY228555 duck circovirus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legnardi, M.; Grassi, L.; Franzo, G.; Menandro, M.L.; Tucciarone, C.M.; Minichino, A.; Dipineto, L.; Borrelli, L.; Fioretti, A.; Cecchinato, M. Detection and Molecular Characterization of a Novel Species of Circovirus in a Tawny Owl (Strix aluco) in Southern Italy. Animals 2022, 12, 135. https://doi.org/10.3390/ani12020135
Legnardi M, Grassi L, Franzo G, Menandro ML, Tucciarone CM, Minichino A, Dipineto L, Borrelli L, Fioretti A, Cecchinato M. Detection and Molecular Characterization of a Novel Species of Circovirus in a Tawny Owl (Strix aluco) in Southern Italy. Animals. 2022; 12(2):135. https://doi.org/10.3390/ani12020135
Chicago/Turabian StyleLegnardi, Matteo, Laura Grassi, Giovanni Franzo, Maria Luisa Menandro, Claudia Maria Tucciarone, Adriano Minichino, Ludovico Dipineto, Luca Borrelli, Alessandro Fioretti, and Mattia Cecchinato. 2022. "Detection and Molecular Characterization of a Novel Species of Circovirus in a Tawny Owl (Strix aluco) in Southern Italy" Animals 12, no. 2: 135. https://doi.org/10.3390/ani12020135
APA StyleLegnardi, M., Grassi, L., Franzo, G., Menandro, M. L., Tucciarone, C. M., Minichino, A., Dipineto, L., Borrelli, L., Fioretti, A., & Cecchinato, M. (2022). Detection and Molecular Characterization of a Novel Species of Circovirus in a Tawny Owl (Strix aluco) in Southern Italy. Animals, 12(2), 135. https://doi.org/10.3390/ani12020135