
Identification of Copy Number Variations and Genetic Diversity in Italian Insular Sheep Breeds

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Genotyping
2.2. CNV and CNVR Detection
2.3. Comparison of CNVRs between Breeds
2.4. Gene Content and Functional Annotation
3. Results
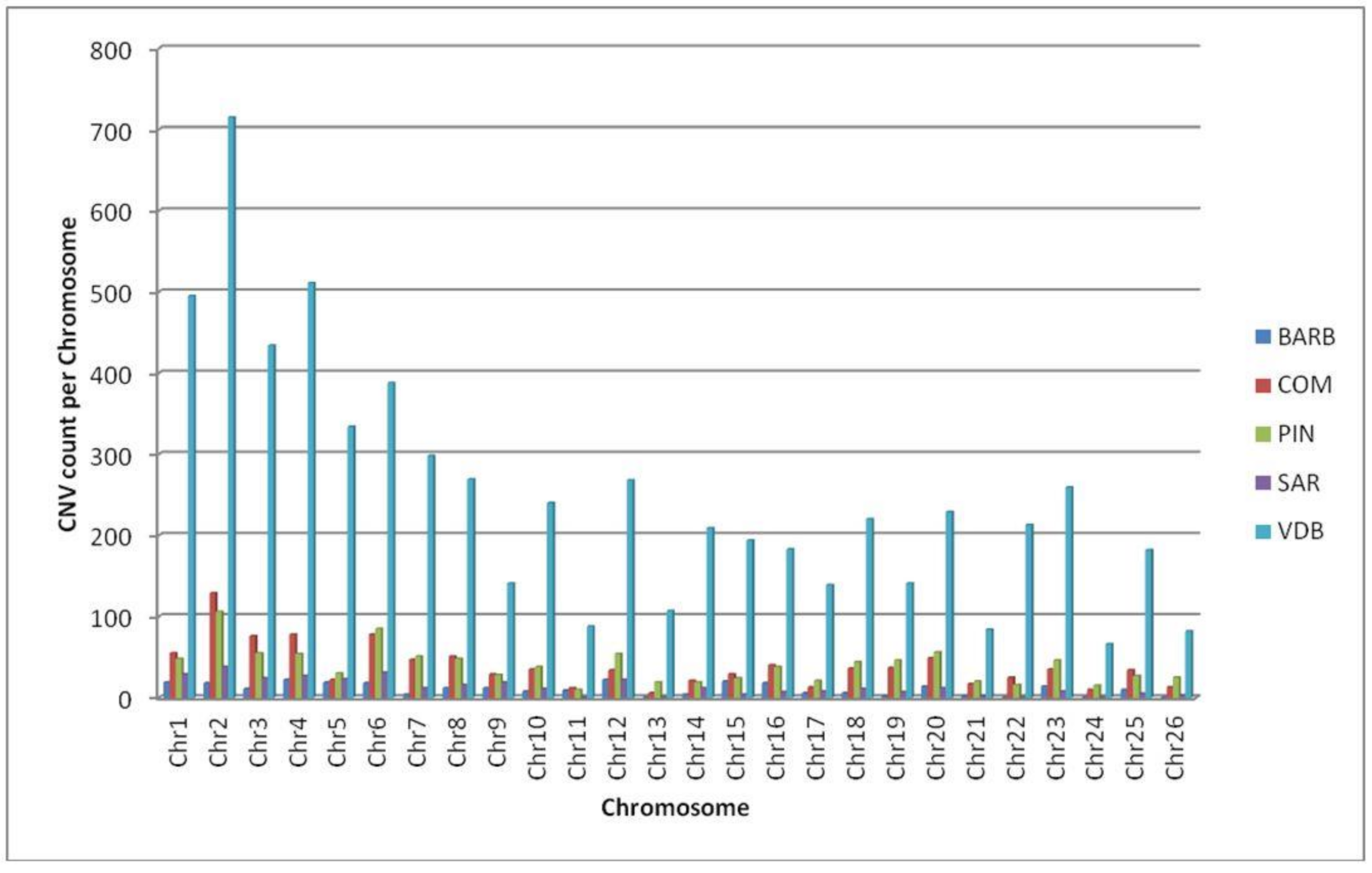
3.1. CNVs and CNVRs Detection
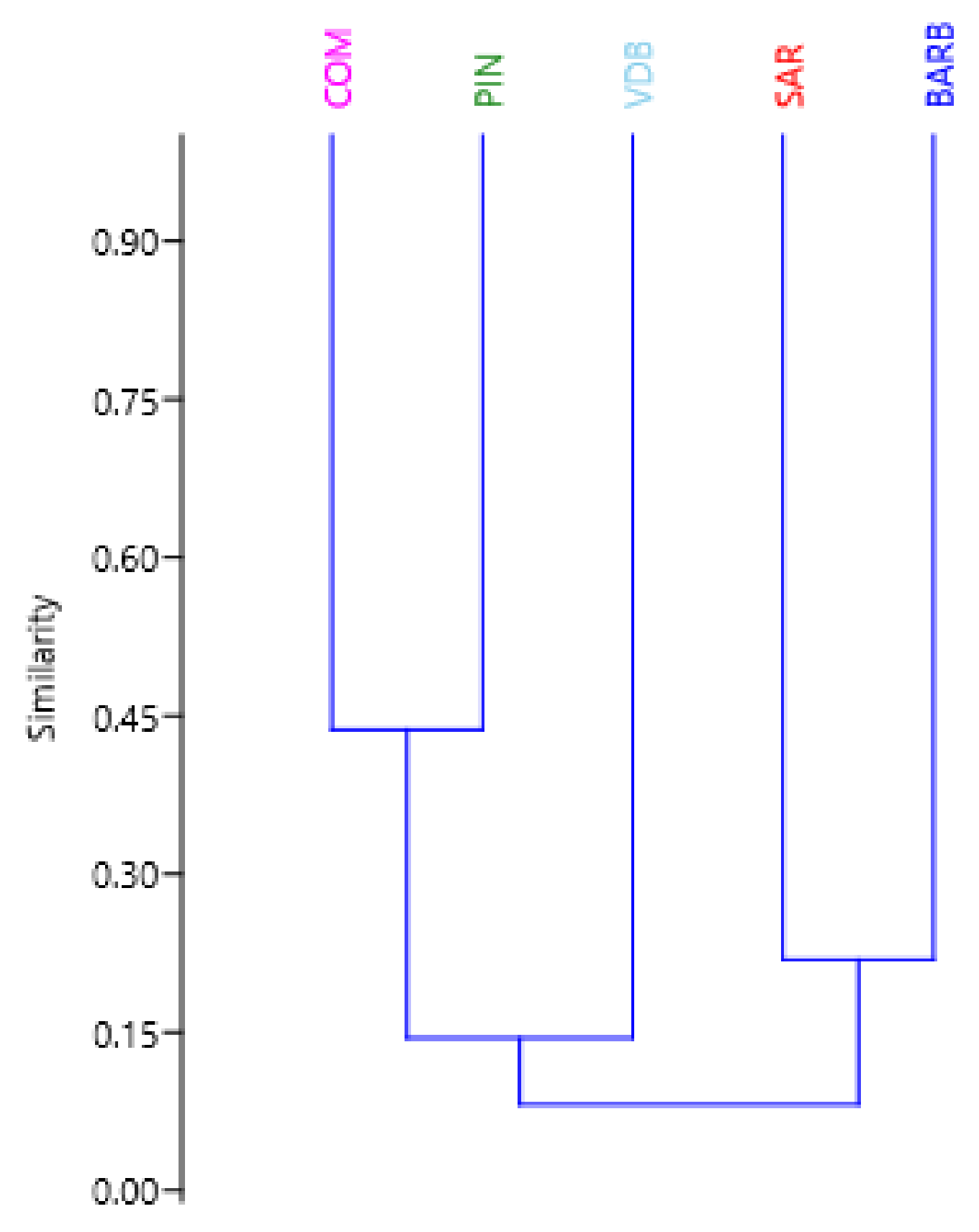
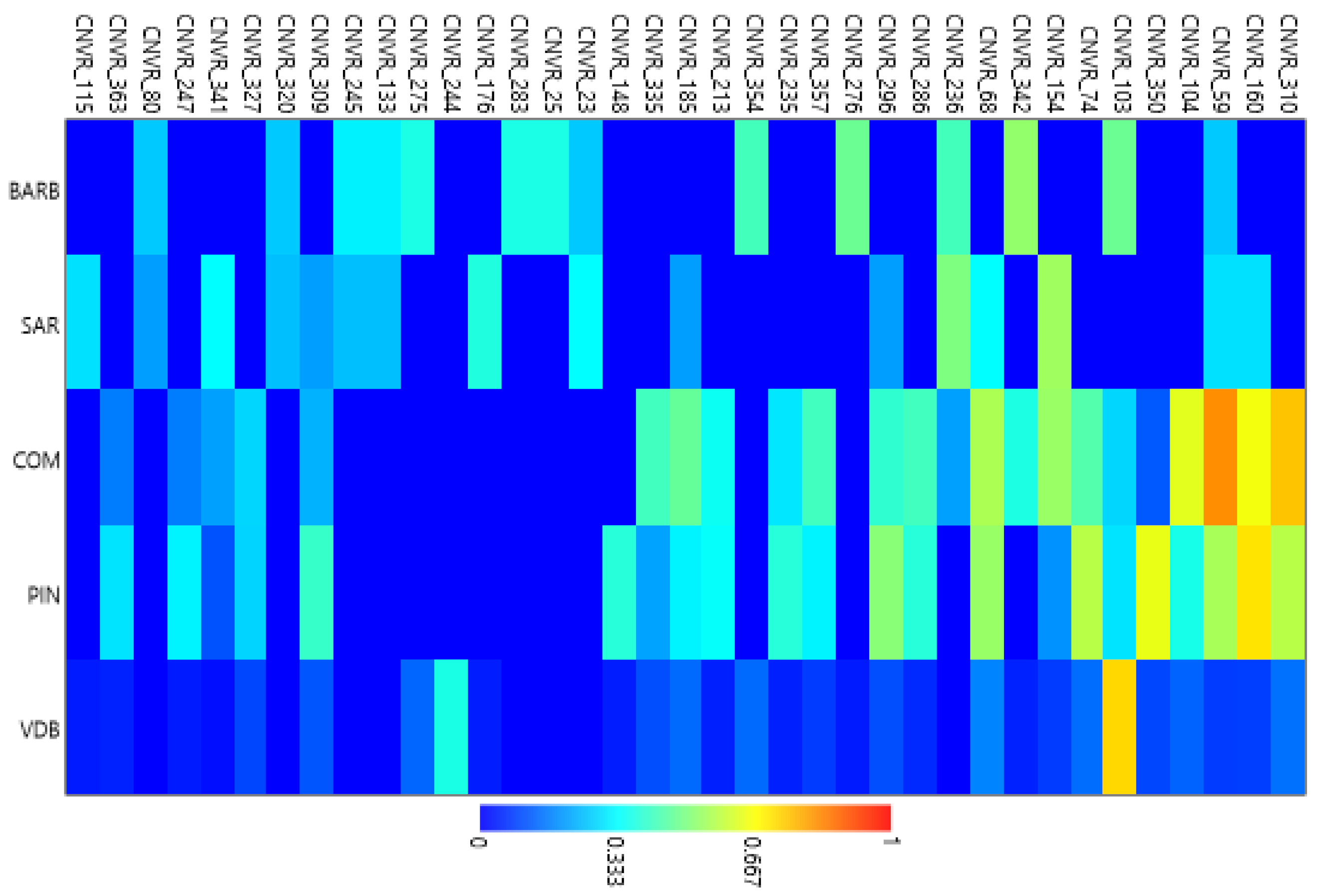
3.2. CNVR Genetic Diversity Analyses
3.3. Gene Enrichment and Functional Annotations of CNVRs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feuk, L.; Carson, A.R.; Scherer, S.W. Structural variation in the human genome. Nat. Rev. Genet. 2006, 7, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, L.; Xu, L.; Ren, H.; Lu, J.; Zhang, X.; Zhang, S.; Zhou, X.; Wei, C.; Zhao, F.; et al. Analysis of copy number variations in the sheep genome using 50K SNP BeadChip array. BMC Genom. 2013, 14, 229. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zhang, Q.; Lu, Z.; Zhao, X.; Zhang, Y. Analysis of copy number variations by SNP50 BeadChip array in Chinese sheep. Genomics 2015, 106, 295–300. [Google Scholar] [CrossRef]
- Yan, J.; Blair, H.T.; Liu, M.; Li, W.; He, S.; Chen, L.; Dittmer, K.E.; Garrick, D.J.; Biggs, P.J.; Dukkipati, V.S.R. Genome-wide detection of autosomal copy number variants in several sheep breeds using Illumina OvineSNP50 BeadChips. Small Rumin. Res. 2017, 155, 24–32. [Google Scholar] [CrossRef]
- Di Gerlando, R.; Sutera, A.M.; Mastrangelo, S.; Tolone, M.; Portolano, B.; Sottile, G.; Bagnato, A.; Strillacci, M.G.; Sardina, M.T. Genome-wide association study between CNVs and milk production traits in Valle del Belìce sheep. PLoS ONE 2019, 14, e0215204. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.D.; Dzama, K.; Hefer, C.A.; Muchadeyi, F.C. Genomic population structure and prevalence of copy number variations in South African Nguni cattle. BMC Genom. 2015, 16, 894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinsen, R.T.M.M.; Strillacci, M.G.; Schiavini, F.; Santus, E.; Rossoni, A.; Maurer, V.; Bieber, A.; Gredler, B.; Dolezal, M.; Bagnato, A. A genome-wide scan of copy number variants using high-density SNPs in Brown Swiss dairy cattle. Livest. Sci. 2016, 191, 153–160. [Google Scholar] [CrossRef]
- Strillacci, M.G.; Gorla, E.; Cozzi, M.C.; Vevey, M.; Genova, F.; Scienski, K.; Longeri, M.; Bagnato, A. A copy number variant scan in the autochthonous Valdostana Red Pied cattle breed and comparison with specialized dairy populations. PLoS ONE 2018, 13, e0204669. [Google Scholar] [CrossRef] [PubMed]
- Di Gerlando, R.; Sardina, M.T.; Tolone, M.; Sutera, A.M.; Mastrangelo, S.; Portolano, B. Genome-wide detection of copy-number variations in local cattle breeds. Anim. Prod. Sci. 2019, 59, 815. [Google Scholar] [CrossRef]
- Paudel, Y.; Madsen, O.; Megens, H.J.; Frantz, L.A.; Bosse, M.; Bastiaansen, J.W.; Crooijmans, R.P.; Groenen, M.A. Evolutionary dynamics of copy number variation in pig genomes in the context of adaptation and domestication. BMC Genom. 2013, 14, 449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholas, T.J.; Baker, C.; Eichler, E.E.; Akey, J.M. A high-resolution integrated map of copy number polymorphisms within and between breeds of the modern domesticated dog. BMC Genom. 2011, 12, 414. [Google Scholar] [CrossRef] [Green Version]
- Molin, A.M.; Berglund, J.; Webster, M.T.; Lindblad-Toh, K. Genome-wide copy number variant discovery in dogs using the CanineHD genotyping array. BMC Genom. 2014, 15, 210. [Google Scholar] [CrossRef] [PubMed]
- Di Gerlando, R.; Mastrangelo, S.; Sardina, M.T.; Ragatzu, M.; Spaterna, A.; Portolano, B.; Ciampolini, R. A genome-wide detection of copy number variations using SNP genotyping arrays in braque français type pyrénées dogs. Animals 2019, 9, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, M.; da Silva, V.H.; Megens, H.J.; Visker, M.H.; Ajmone-Marsan, P.; Bâlteanu, V.A.; Groenen, M.A. Distribution and functionality of copy number variation across European cattle populations. Front. Genet. 2017, 8, 108. [Google Scholar] [CrossRef]
- Pierce, M.D.; Dzama, K.; Muchadeyi, F.C. Genetic diversity of seven cattle breeds inferred using copy number variations. Front. Genet. 2018, 9, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzi, M.C.; Martinez-Ruiz, C.P.; Roman-Ponce, S.I.; Murillo, V.E.V.; Utrera, Á.R.; Montaño-Bermúdez, M.M.; Martinez-Velazquez, G.; Strillacci, M.G. Copy number variants reveal genomic diversity in a Mexican creole cattle population. Livest. Sci. 2019, 229, 194–202. [Google Scholar] [CrossRef]
- Yang, L.; Xu, L.; Zhou, Y.; Liu, M.; Wang, L.; Kijas, J.W.; Liu, G.E. Diversity of copy number variation in a worldwide population of sheep. Genomics 2018, 110, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhou, Y.; Rosen, B.D.; Van Tassell, C.P.; Stella, A.; Tosser-Klopp, G.; Crepaldi, P. Diversity of copy number variation in the worldwide goat population. Heredity 2019, 122, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Strillacci, M.G.; Gorla, E.; Ríos-Utrera, A.; Vega-Murillo, V.E.; Montaño-Bermudez, M.; Garcia-Ruiz, A.; Bagnato, A. Copy number variation mapping and genomic variation of autochthonous and commercial turkey populations. Front. Genet. 2019, 10, 982. [Google Scholar] [CrossRef]
- Matukumalli, L.K.; Lawley, C.T.; Schnabel, R.D.; Taylor, J.F.; Allan, M.F.; Heaton, M.P.; Van Tassell, C.P. Development and characterization of a high density SNP genotyping assay for cattle. PLoS ONE 2009, 4, e5350. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Liu, G.E.; Bickhart, D.M.; Cardone, M.F.; Wang, K.; Kim, E.S.; Van Tassell, C.P. Genomic characteristics of cattle copy number variations. BMC Genom. 2011, 12, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.E.; Hou, Y.; Zhu, B.; Cardone, M.F.; Lu, J.; Cellamare, A.; Mitra, A.; Alexander, L.J.; Coutinho, L.L.; Dell’Aquila, M.E.; et al. Analysis of Copy Number Variations among diverse cattle breeds. Genome Res. 2010, 20, 693–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorla, E.; Cozzi, M.C.; Román-Ponce, S.I.; López, F.R.; Vega-Murillo, V.E.; Cerolini, S.; Strillacci, M.G. Genomic variability in Mexican chicken population using copy number variants. BMC Genet. 2017, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tolone, M.; Mastrangelo, S.; Rosa, A.J.M.; Portolano, B. Genetic diversity and population structure of Sicilian sheep breeds using microsatellite markers. Small Rumin. Res. 2012, 102, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Mastrangelo, S.; Di Gerlando, R.; Tolone, M.; Tortorici, L.; Sardina, M.T.; Portolano, B. Genome wide linkage disequilibrium and genetic structure in Sicilian dairy sheep breeds. BMC Genet. 2014, 15, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrangelo, S.; Criscione, A.; Sottile, G.; Portolano, B.; Marletta, D.; Bordonaro, S. Genome-wide analysis identifies potentially causative genes explaining the phenotypic variability in Pinzirita sheep. Anim. Genet. 2019, 50, 189–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrangelo, S.; Ben Jemaa, S.; Sottile, G.; Casu, S.; Portolano, B.; Ciani, E.; Pilla, F. Combined approaches to identify genomic regions involved in phenotypic differentiation between low divergent breeds: Application in Sardinian sheep populations. J. Anim. Breed. Genet. 2019, 136, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Cesarani, A.; Sechi, T.; Gaspa, G.; Usai, M.G.; Sorbolini, S.; Macciotta, N.P.P.; Carta, A. Investigation of genetic diversity and selection signatures between Sarda and Sardinian Ancestral black, two related sheep breeds with evident morphological differences. Small Rumin. Res. 2019, 177, 68–75. [Google Scholar] [CrossRef]
- Mastrangelo, S.; Sardina, M.T.; Riggio, V.; Portolano, B. Study of polymorphisms in the promoter region of ovine β-lactoglobulin gene and phylogenetic analysis among the Valle del Belice breed and other sheep breeds considered as ancestors. Mol. Biol. Rep. 2012, 39, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Mastrangelo, S.; Portolano, B.; Di Gerlando, R.; Ciampolini, R.; Tolone, M.; Sardina, M.T. International Sheep Genomics Consortium. Genome-wide analysis in endangered populations: A case study in Barbaresca sheep. Animal 2017, 11, 1107–1116. [Google Scholar] [CrossRef]
- Mastrangelo, S.; Moioli, B.; Ahbara, A.; Latairish, S.; Portolano, B.; Pilla, F.; Ciani, E. Genome-wide scan of fat-tail sheep identifies signals of selection for fat deposition and adaptation. Anim. Prod. Sci. 2018, 59, 835–848. [Google Scholar] [CrossRef]
- Durán Aguilar, M.; Román Ponce, S.I.; Ruiz Lopez, F.J.; González Padilla, E.; Vásquez Peláez, C.G.; Bagnato, A.; Strillacci, M.G. Genome-wide association study for milk somatic cell score in Holstein cattle using copy number variation as markers. J. Anim. Breed. Genet. 2017, 134, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; Darvishi, K.; Shi, X.; Rajan, D.; Rigler, D.; Fitzgerald, T.; Lionel, A.C.; Thiruvahindrapuram, B.; MacDonald, J.R.; Mills, R.; et al. Comprehensive assessment of arraybased platforms and calling algorithms for detection of copy number variants. Nat. Biotechnol. 2011, 8, 512–520. [Google Scholar] [CrossRef] [Green Version]
- Diskin, S.J.; Li, M.; Hou, C.; Yang, S.; Glessner, J.; Hakonarson, H.; Bucan, M.; Maris, J.M.; Wang, K. Adjustment of genomic waves in signal intensities from whole-genome SNP genotyping platforms. Nucleic Acids Res. 2008, 36, e126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redon, R.; Ishikawa, S.; Fitch, K.R.; Feuk, L.; Perry, G.H.; Andrews, T.D.; Fiegler, H.; Shapero, M.H.; Carson, A.R.; Chen, W.; et al. Global variation in copy number in the human genome. Nature 2006, 444, 444–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef] [PubMed]
- Ciani, E.; Crepaldi, P.; Nicoloso, L.; Lasagna, E.; Sarti, F.M.; Moioli, B.; Pilla, F. Genome-wide analysis of I talian sheep diversity reveals a strong geographic pattern and cryptic relationships between breeds. Anim. Genet. 2014, 45, 256–266. [Google Scholar] [CrossRef]
- Mastrangelo, S.; Ciani, E.; Sardina, M.T.; Sottile, G.; Pilla, F.; Portolano, B.; Bi.Ov. Ita Consortium. Runs of homozygosity reveal genome-wide autozygosity in Italian sheep breeds. Anim. Genet. 2018, 49, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyache, F.; Fernández, I.; Tapsoba, A.S.R.; Traoré, A.; Menéndez-Arias, N.A.; Álvarez, I. Functional characterization of Copy Number Variations regions in Djallonké sheep. J. Anim. Breed. Genet. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Fan, H.; Yuan, Z.; Hu, S.; Ma, X.; Xuan, J.; Wang, H.; Zhang, L.; Wei, C.; Zhang, Q.; et al. Genome-wide detection of CNVs in Chinese indigenous sheep with different types of tails using ovine high-density 600K SNP arrays. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Liu, X.; Pan, J.; Ma, L.; Ma, Y.; He, X.; Jiang, L. Genome-wide detection of copy number variation in Chinese indigenous sheep using an ovine high-density 600 K SNP array. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Guo, J.; Guo, Y.; Yang, Y.; Teng, T.; Yu, Q.; Wang, T.; Zhou, M.; Zhu, Q.; Wang, W.; et al. Genome-wide detection of CNVs and association with body weight in sheep based on 600K SNP arrays. Front. Genet. 2020, 11, 558. [Google Scholar] [CrossRef]
- Fontanesi, L.; Beretti, F.; Martelli, P.L.; Colombo, M.; Dall’Olio, S.; Occidente, M.; Portolano, B.; Casadio, R.; Matassino, D.; Russo, V. A first comparative map of copy number variations in the sheep genome. Genomics 2011, 97, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Tolone, M.; Giosa, D.; Di Gerlando, R.; Sutera, A.M.; Moscarelli, A.; Rizzuto, I.; Portolano, B.; Mastrangelo, S. Whole-genome resequencing reveals specific genomic variants in Italian insular sheep breeds. Ital. J. Anim. Sci. 2021, 20, 70–71. [Google Scholar]
- Xu, L.; Hou, Y.; Bickhart, D.M.; Zhou, Y.; Song, J.; Sonstegard, T.S.; Van Tassell, C.P.; Liu, G.E. Population-genetic properties of differentiated copy number variations in cattle. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solé, M.; Ablondi, M.; Binzer-Panchal, A.; Velie, B.D.; Hollfelder, N.; Buys, N.; Ducro, B.J.; François, L.; Janssens, S.; Schurink, A.; et al. Inter-and intra-breed genome-wide copy number diversity in a large cohort of European equine breeds. BMC Genom. 2019, 20, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bickhart, D.M.; Xu, L.; Hutchison, J.L.; Cole, J.B.; Null, D.J.; Schroeder, S.G.; Song, J.; Garcia, J.F.; Sonstegard, T.S.; Van Tassell, C.P.; et al. Diversity and population-genetic properties of copy number variations and multicopy genes in cattle. DNA Res. 2016, 23, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Perry, G.H.; Dominy, N.J.; Claw, K.G.; Lee, A.S.; Fiegler, H.; Redon, R.; Werner, J.; Villanea, F.A.; Mountain, J.L.; Misra, R.; et al. Diet and the evolution of human amylase gene copy number variation. Nat. Genet. 2007, 39, 1256–1260. [Google Scholar] [CrossRef] [Green Version]
- May, P.; Woldt, E.; Matz, R.L.; Boucher, P. The LDL receptor-related protein (LRP) family: An old family of proteins with new physiological functions. Ann. Med. 2007, 39, 219–228. [Google Scholar] [CrossRef] [PubMed]
- RincónFlórez, J.C.; López Herrera, A.; EcheverriZuluaga, J.J. Genome-wide association study using the Bayes C method for important traits in dairy yield in Colombian Holstein Cattle. Acta Sci. Anim. Sci. 2018, 40, 39015. [Google Scholar] [CrossRef]
- Peng, W.F.; Xu, S.S.; Ren, X.; Lv, F.H.; Xie, X.L.; Zhao, Y.X.; Zhang, M.; Shen, Z.Q.; Ren, Y.L.; Gao, L.; et al. A genome-wide association study reveals candidate genes for the supernumerary nipple phenotype in sheep (Ovis aries). Anim. Genet. 2017, 48, 570–579. [Google Scholar] [CrossRef] [PubMed]
- De León, C.; Manrique, C.; Martínez, R.; Rocha, J.F. Research Article Genomic association study for adaptability traits in four Colombian cattle breeds. Genet. Mol. Res. 2019, 18, gmr18373. [Google Scholar] [CrossRef]
- Islam, R.; Liu, X.; Gebreselassie, G.; Abied, A.; Ma, Q.; Ma, Y. Genome-wide association analysis reveals the genetic locus for high reproduction trait in Chinese Arbas Cashmere goat. Genes Genom. 2020, 42, 893–899. [Google Scholar] [CrossRef]
- Salehian-Dehkordi, H.; Xu, Y.X.; Xu, S.S.; Li, X.; Luo, L.Y.; Liu, Y.J.; Wang, D.F.; Cao, Y.H.; Shen, M.; Gao, L.; et al. Genome-Wide Detection of Copy Number Variations and Their Association with Distinct Phenotypes in the World’s Sheep. Front. Genet. 2021, 12, 670582. [Google Scholar] [CrossRef]
- He, M.; Rutledge, S.L.; Kelly, D.R.; Palmer, C.A.; Murdoch, G.; Majumder, N.; Nicholls, R.D.; Pei, Z.; Watkins, P.A.; Vockley, J. A new genetic disorder in mitochondrial fatty acid β-oxidation: ACAD9 deficiency. Am. J. Hum. Genet. 2007, 81, 87–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Luo, J.; Wang, H.; Shi, H.; Zhu, J.; Sun, Y.; Yu, K.; Yao, D. Adipose triglyceride lipase regulates lipid metabolism in dairy goat mammary epithelial cells. Gene 2015, 554, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Puig-Oliveras, A.; Revilla, M.; Castelló, A.; Fernández, A.I.; Folch, J.M.; Ballester, M. Expression-based GWAS identifies variants, gene interactions and key regulators affecting intramuscular fatty acid content and composition in porcine meat. Sci. Rep. 2016, 6, 31803. [Google Scholar] [CrossRef]
- Gui, L.; Hong, J.; Raza, S.H.A.; Zan, L. Genetic variants in SIRT3 transcriptional regulatory region affect promoter activity and fat deposition in three cattle breeds. Mol. Cell. Probes 2017, 32, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Izadnia, H.R.; Tahmoorespur, M.; Bakhtiarizadeh, M.R.; Nassiri, M.; Esmaeilkhanien, S. Gene expression profile analysis of residual feed intake for Isfahan native chickens using RNA-SEQ data. Ital. J. Anim. Sci. 2019, 18, 246–260. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Huang, J.; Wang, X.; Ma, Y. Transcription factors regulate adipocyte differentiation in beef cattle. Anim. Genet. 2020, 51, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Takegahara, N.; Takamatsu, H.; Toyofuku, T.; Tsujimura, T.; Okuno, T.; Yukawa, K.; Mizui, M.; Yamamoto, M.; Prasad, D.V.R.; Suzuki, K.; et al. Plexin-A1 and its interaction with DAP12 in immune responses and bone homeostasis. Nat. Cell Biol. 2006, 8, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Feske, S. ORAI1 and STIM1 deficiency in human and mice: Roles of store-operated Ca2+ entry in the immune system and beyond. Immunol. Rev. 2009, 231, 189–209. [Google Scholar] [CrossRef]
- Ripoll, V.M.; Meadows, N.A.; Bangert, M.; Lee, A.W.; Kadioglu, A.; Cox, R.D. Nicotinamide nucleotide transhydrogenase (NNT) acts as a novel modulator of macrophage inflammatory responses. FASEB J. 2012, 26, 3550–3562. [Google Scholar] [CrossRef]
- Wilk, E.; Pandey, A.K.; Leist, S.R.; Hatesuer, B.; Preusse, M.; Pommerenke, C.; Wang, J.; Schughart, K. RNAseq expression analysis of resistant and susceptible mice after influenza A virus infection identifies novel genes associated with virus replication and important for host resistance to infection. BMC Genom. 2015, 16, 655. [Google Scholar] [CrossRef]
- Bassano, I.; Ong, S.H.; Sanz-Hernandez, M.; Vinkler, M.; Kebede, A.; Hanotte, O.; Onuigbo, E.; Fife, M.; Kellam, P. Comparative analysis of the chicken IFITM locus by targeted genome sequencing reveals evolution of the locus and positive selection in IFITM1 and IFITM3. BMC Genom. 2019, 20, 272. [Google Scholar] [CrossRef] [PubMed]
- González-Ruiz, S.; Strillacci, M.G.; Durán-Aguilar, M.; Cantó-Alarcón, G.J.; Herrera-Rodríguez, S.E.; Bagnato, A.; Guzmán, L.F.; Milián-Suazo, F.; Román-Ponce, S.I. Genome-Wide Association Study in Mexican Holstein Cattle Reveals Novel Quantitative Trait Loci Regions and Confirms Mapped Loci for Resistance to Bovine Tuberculosis. Animals 2019, 9, 636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Nguyen, D.T.; Choi, M.; Cha, S.E.; Kim, J.H.; Dadi, H.; Seo, H.S.; Seo, H.; Chun, T.; Park, C. Analysis of cattle olfactory subgenome: The first detail study on the characteristics of the complete olfactory receptor repertoire of a ruminant. BMC Genom. 2013, 14, 596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palouzier-Paulignan, B.; Lacroix, M.C.; Aimé, P.; Baly, C.; Caillol, M.; Congar, P.; Karyn Julliard, P.; Tucker, K.; Fadool, D.A. Olfaction under metabolic influences. Chem. Senses 2012, 37, 769–797. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.; Machado, P.C.; Pinto, L.F.B.; Silva, M.R.; Schenkel, F.S.; Brito, L.F.; Pedrosa, V.B. Genome-wide association study and pathway analysis for fat deposition traits in nellore cattle raised in pasture–based systems. J. Anim. Breed. Genet. 2021, 138, 360–378. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Seibert, J.T.; Edea, Z.; Graves, K.L.; Kim, E.S.; Keating, A.F.; Baumgard, L.H.; Ross, J.W.; Rothschild, M.F. Characterization of the acute heat stress response in gilts: III. Genome-wide association studies of thermotolerance traits in pigs. J. Anim. Sci. 2018, 96, 2074–2085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moioli, B.; Pilla, F.; Ciani, E. Signatures of selection identify loci associated with fat tail in sheep. J. Anim. Sci. 2015, 93, 4660–4669. [Google Scholar] [CrossRef] [PubMed]
- Mwacharo, J.M.; Kim, E.S.; Elbeltagy, A.R.; Aboul-Naga, A.M.; Rischkowsky, B.A.; Rothschild, M.F. Genomic footprints of dryland stress adaptation in Egyptian fat-tail sheep and their divergence from East African and western Asia cohorts. Sci. Rep. 2017, 7, 17647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Liu, E.; Liu, Z.; Kijas, J.W.; Zhu, C.; Hu, S.; Ma, S.; Zhang, L.; Du, L.; Wang, H.; et al. Selection signature analysis reveals genes associated with tail type in Chinese indigenous sheep. Anim. Genet. 2017, 48, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.S.; Ren, X.; Yang, G.L.; Xie, X.L.; Zhao, Y.X.; Zhang, M.; Shen, Z.Q.; Ren, Y.L.; Gao, L.; Shen, M. Genome-wide association analysis identifies the genetic basis of fat deposition in the tails of sheep (Ovis aries). Anim. Genet. 2017, 48, 560–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | N. Sample | N. CNVs | CNVs per Sample Min–Max (Average) | Loss | Gain | Min Length (bp) | Max Length (bp) | Mean Length (bp) |
|---|---|---|---|---|---|---|---|---|
| BARB | 19 | 431 | 15–33 (23) | 328 | 103 | 19,028 | 2,499,938 | 222,990 |
| COM | 43 | 1172 | 18–39 (27) | 957 | 215 | 19,041 | 3,660,245 | 344,790 |
| PIN | 47 | 1216 | 11–43 (26) | 963 | 253 | 23,587 | 4,399,691 | 399,121 |
| SAR | 24 | 481 | 12–28 (20) | 335 | 146 | 23,587 | 3,692,295 | 272,509 |
| VDB | 294 | 6907 | 11–52 (24) | 5469 | 1438 | 13,128 | 14,995,713 | 569,560 |
| Total | 427 | 10,207 | 11–52 (24) | 8052 | 2155 | 13,128 | 14,995,713 | 1,808,970 |
| Breed | N. CNVRs | Loss | Gain | Min Length (bp) | Max Length (bp) | Mean Length (bp) |
|---|---|---|---|---|---|---|
| BARB | 83 | 61 | 22 | 42,405 | 2,013,519 | 178,021 |
| COM | 195 | 159 | 36 | 19,322 | 3,295,789 | 324,599 |
| PIN | 186 | 147 | 39 | 23,587 | 2,962,879 | 347,976 |
| SAR | 89 | 61 | 28 | 43,456 | 1,954,981 | 196,644 |
| VDB | 687 | 532 | 155 | 14,264 | 11,305,268 | 434,543 |
| Total | 1240 | 960 | 280 | 14,264 | 11,305,268 | 1,481,783 |
| BARB | COM | PIN | SAR | VDB | |
|---|---|---|---|---|---|
| BARB | 0.000 | ||||
| COM | 0.255 | 0.000 | |||
| PIN | 0.264 | 0.077 | 0.000 | ||
| SAR | 0.220 | 0.196 | 0.203 | 0.000 | |
| VDB | 0.097 | 0.126 | 0.114 | 0.084 | 0.000 |
| Accession Number | Biological Process | Gene Symbol |
| GO:0065007 | Biological regulation | DEAF1, HRAS, HNRNPF, PIDD1, TFAP2D, EPS8L2, EEFSEC, PAIP1, PNPLA2, KLF15, EPHB3, TXNRD3, ZXDC, FXYD4, CNBP, PLXNA1, GATA2, KHDRBS2, RUVBL1, CREB1, PDGFA, STIM1, IRF7, SYT4, SENP6, STIM1, TBL1XR1 |
| GO:0009987 | Cellular process | DEAF1, PSMD13, HRAS, EFCC1, HNRNPF, SUN1, PIDD1, TFAP2D, EPS8L2, MCM2, EEFSEC, MYO6, CHCHD6, METTL21A, NTNG2, PAIP1, PNPLA2, TMEM80, NNT, KLF15, EPHB3, FAM20C, SEC61A1, PKP3, GET4, SLC25A22, RPN1, RRM1, PKP3, TXNRD, ZXDC, RAB7A, ISY1, COPG1, FXYD4, RTKN2, CNBP, PLXNA1, GATA2, KHDRBS2, MYO6, RUVBL1, ANO9, CREB1, PDGFA, CHCHD6, STIM1, INTS1, IRF7, SYT4, SENP6, STIM1, SHROOM3, TBL1XR1, EFCC1, IQSEC1 |
| GO:0008152 | Metabolic process | DEAF1, PSMD13, EFCC1, HNRNPF, PIDD1, TFAP2D, MCM2, EEFSEC, METTL21A, PAIP1, PNPLA2, NNT, KLF15, EPHB3, FAM20C, SEC61A1, SLC25A22, RPN1, RRM1, ZXDC, METTL21A, ISY1, CNBP, GATA2, KHDRBS2, RUVBL1, CREB1, PDGFA, INTS1, IRF7 |
| GO:0050896 | Response to stimulus | HRAS, PIDD1, EPS8L2, NLRP6, MCM2, CHCHD6, NTNG2, EPHB3, PLXNA1, PDGFA, CHCHD6, SYT4 |
| GO:0002376 | Immune system process | ANO9, SIGGIR, PKP3, STIM1, PLXNA1, IFITM3, IFITN5, NNT, CREB1, IRF7 |
| Accession number | Molecular function | Gene Symbol |
| GO:0005488 | Binding | DEAF1, HRAS, EFCC1, HNRNPF, SUN1, TFAP2D, EPS8L2, MCM2, EEFSEC, MYO6, PAIP1, NNT, ACAD9, KLF15, SEC61A1, PKP3, RRM1, PKP3, RTKN2, CNBP, GATA2, KHDRBS2, MYO6, CREB1, PDGFA, STIM1, IRF7, POLR2L, SYT4, STIM1, SHROOM3, EFCC1 |
| GO:0003824 | Catalytic activity | HRAS, PIDD1, MGLL, MCM2, MYO6, METTL21A, PNPLA2, B4GALNT4, NNT, ACAD9, EPHB3, FAM20C, ALDH1L1, RRM1, TXNRD3, MYO6, RUVBL1, POLR2L, SENP6 |
| GO:0098772 | Molecular function regulator | DEAF1, EFCC1, TFAP2D, EPS8L2, KLF15, ZXDC, FXYD4, GATA2, CREB1, STIM1, IRF7, TBL1XR1 |
| GO:0005215 | Trasporter activity | SEC61A1, SLC25A22, FXYD4, ANO9, STIM1 |
| Accession number | Cellular Component | Gene Symbol |
| GO:0110165 | Cellular anatomical entity | DEAF1, PSMD13, ABTB1, HRAS, EFCC1, HNRNPF, SUN1, PIDD1, MGLL, LRP1B, STBD1, TFAP2D, EPS8L2, MCM2, MYO6, CHCHD6, METTL21A, NTNG2, PNPLA2, TMEM80, NNT, ACAD9, KLF15, EPHB3, FAM20C, SEC61A1, PKP3, GET4, RPN1, RRM1, TXNRD3, NUP210, ZXDC, CD151, RAB7A, RAB43, ISY1, COPG1, RTKN2, CNBP, IFITM5, PLXNA1, GATA2, KHDRBS2, RUVBL1, ANO9, CREB1, PDGFA, SLC41A3, INTS1, IRF7, SYT4, SENP6, STIM1, SHROOM3, TBL1XR1, ADAP1 |
| GO:0005622 | Intracellular | DEAF1, PSMD13, ABTB1, EFCC1, HNRNPF, SUN1, PIDD1, TFAP2D, MCM2, MYO6, CHCHD6, METTL21A, PNPLA2, NNT, ACAD9, KLF15, FAM20C, SEC61A1, PKP3, GET4, RPN1, RRM1, TXNRD3, NUP210, ZXDC, RAB7A, RAB43, ISY1, COPG1, RTKN2, CNBP, GATA2, KHDRBS2, RUVBL1, CREB1, STIM1, INTS1, IRF7, SYT4, SENP6, SHROOM3, TBL1XR1, ADAP1 |
| GO:0032991 | Protein-containing complex | PSMD13, ABTB1, HNRNPF, SUN1, MCM2, TMEM80, EPHB3, SEC61A1, GET4, RPN1, RRM1, NUP210, ISY1, COPG1, PLXNA1, RUVBL1, CREB1, INTS1, TBL1XR1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Gerlando, R.; Mastrangelo, S.; Tolone, M.; Rizzuto, I.; Sutera, A.M.; Moscarelli, A.; Portolano, B.; Sardina, M.T. Identification of Copy Number Variations and Genetic Diversity in Italian Insular Sheep Breeds. Animals 2022, 12, 217. https://doi.org/10.3390/ani12020217
Di Gerlando R, Mastrangelo S, Tolone M, Rizzuto I, Sutera AM, Moscarelli A, Portolano B, Sardina MT. Identification of Copy Number Variations and Genetic Diversity in Italian Insular Sheep Breeds. Animals. 2022; 12(2):217. https://doi.org/10.3390/ani12020217
Chicago/Turabian StyleDi Gerlando, Rosalia, Salvatore Mastrangelo, Marco Tolone, Ilaria Rizzuto, Anna Maria Sutera, Angelo Moscarelli, Baldassare Portolano, and Maria Teresa Sardina. 2022. "Identification of Copy Number Variations and Genetic Diversity in Italian Insular Sheep Breeds" Animals 12, no. 2: 217. https://doi.org/10.3390/ani12020217
APA StyleDi Gerlando, R., Mastrangelo, S., Tolone, M., Rizzuto, I., Sutera, A. M., Moscarelli, A., Portolano, B., & Sardina, M. T. (2022). Identification of Copy Number Variations and Genetic Diversity in Italian Insular Sheep Breeds. Animals, 12(2), 217. https://doi.org/10.3390/ani12020217