Simple Summary
Today’s swine production systems are mostly intensive, and pigs are continuously exposed to pathogens and immune stimulatory antigens that may negatively impact productivity. It is known that immune system stimulation may reduce appetite, growth, and nutrient use efficiency compared to healthy animals. On the other hand, there is progressive pressure towards the reduction in antibiotic usage in livestock production, which highlights the need for furthering our understanding of the relationship between nutrition and the immune response. Swine nutritionists will need to consider the role of nutrition on health in order to develop programs that support production and robustness of pigs under a variety of stressful conditions. Among the nutritional strategies whose utilization may directly impact on the immune status of pigs, feeding low protein, amino acid-supplemented diets, supplementation of functional amino acids, dietary fiber level and source, diet complexity, organic acids, and plant secondary metabolites are at the forefront. As such, this review will discuss the impact of immune status on swine production and the interaction between nutrients and animal health, focusing on the roles of each nutritional strategy during times of immune challenge.
Abstract
Our understanding of nutrition has been evolving to support both performance and immune status of pigs, particularly in disease-challenged animals which experience repartitioning of nutrients from growth towards the immune response. In this sense, it is critical to understand how stress may impact nutrient metabolism and the effects of nutritional interventions able to modulate organ (e.g., gastrointestinal tract) functionality and health. This will be pivotal in the development of effective diet formulation strategies in the context of improved animal performance and health. Therefore, this review will address qualitative and quantitative effects of immune system stimulation on voluntary feed intake and growth performance measurements in pigs. Due to the known repartitioning of nutrients, the effects of stimulating the immune system on nutrient requirements, stratified according to different challenge models, will be explored. Finally, different nutritional strategies (i.e., low protein, amino acid-supplemented diets; functional amino acid supplementation; dietary fiber level and source; diet complexity; organic acids; plant secondary metabolites) will be presented and discussed in the context of their possible role in enhancing the immune response and animal performance.
1. Nutrition and Health: The Merge
Current nutrient requirement estimates are largely based on growth performance response, however, nutrient effects on non-proteinogenic functions (e.g., intestinal development, immune status, and response) has been increasingly demonstrated. With legislation introduced in some countries to eliminate use of antimicrobial growth promoters and increasing pressure to reduce overall antibiotic use in livestock production, it will become increasingly important to understand how immune status affects nutrient requirements and how feed formulations can be adjusted to support animal robustness in addition to growth performance. The impact of disease-challenge can be substantial. For example, pigs kept in low sanitary conditions (LSC; e.g., lack of vaccination, environmental hygiene and biosecurity protocol, and preventive antibiotic injection) having a 55 g/d reduction in daily gain during the complete fattening period compared to counterparts housed in high sanitary conditions (HSC; [1]). This indicates a high cost associated with the activation of the immune system, including a higher production of immune cells and signaling molecules as well as losses in efficiency of affected tissues, which will decrease the efficiency of nutrient utilization for body protein deposition [2]. It is reasonable to infer that nutritionists must consider the components associated with building a strong immune system when formulating swine diets in commercial operations. Of note, while nutritional strategies may support animal robustness during a disease and recovery from illness, attention should also be given to dietary strategies that trigger excessive immune activation and may impair the efficiency of animal production.
Additional evidence for the importance of nutrition for immune status lies in the endocrine regulation of appetite and growth. Specifically, the reduction in performance due to immune system activation is not entirely, and not always, associated with a reduction in feed intake. Upon exposure to stressful situations, mononuclear myeloid cells of the innate immune system synthesize and release proinflammatory cytokines such as interleukin (IL)-1b, IL-6 and tumor necrosis factor-α (TNF-α) [3]. These cytokines will orchestrate (1) a redirection of nutrients from growth towards the immune system [3,4], (2) a specific response of the central nervous system with fever, decreased feed intake, and activation of hypothalamic–pituitary–adrenal axis [4,5], (3) a suppression of gastric emptying and motility [4], and (4) an increased concentration of glucagon, insulin, and leptin in blood [6]. These events combine to affect feed intake and weight gain, and guide how inflammation impacts growth.
In order to meet growing demand, pork production will need to increase its sustainability and efficiency, which will encompass the development of nutritional programs which support animal growth performance and robustness under commercial conditions. Several nutritional strategies have been identified as potential governors of the interaction between growth and health in pigs, including feeding low protein (LP) diets, functional amino acid (FAA) supplementation, and provision of fiber sources and natural (e.g., phytochemicals) compounds. As such, the present review will discuss the impact of different immune stimulating conditions on nutrient digestion and metabolism and how different nutritional interventions may help nutritionists formulate diets for improved health status of pigs. One important aspect approached by the present review is the differentiation between (1) situations where stressors decrease feed intake and consequently impair growth performance vs. (2) the effects of immune system stimulation on nutrient metabolism.
2. Immune System Stimulation, Nutrient Efficiency and Requirements
Immune stimulation and inflammation result in a number of effects in the pig, most notably a reduction in voluntary feed intake and efficiency of nutrient utilization, even in the absence of clinical manifestation of disease [7], although this may not always be the case. While both feed intake and nutrient utilization efficiency influence pig performance, the extent to which reductions in these factors are related to the reduction in performance is dependent on the specific immune challenge [8]. For example, Rodrigues et al. [9] showed that the reduction in growth in challenged compared with control pigs associated with a reduction in feed intake was 14, 4, 45, 1, and 1% in pigs challenged with enteric pathogens, environmental stressors, bacterial lipopolysaccharide (LPS), respiratory pathogens, and sanitary conditions, respectively. This is important to note as, in experimental environments, several challenge models are used to investigate stimulation of the immune system and the associated effects on outcomes such as growth performance impairment, nutrient requirements, and immune status. These include challenge with enteric [10,11] and respiratory pathogens [12,13], housing pigs under heat stress, high stocking densities, or suboptimal sanitary conditions [1,14,15,16,17], and triggering systemic immune stimulation using LPS [18]. Overall, it is likely that the effect of any nutrition intervention in challenged pigs will be, at least in part, due to the specific challenge model used and results may not be transferable to other conditions.
Another important factor differentially impacting performance response in challenged pigs is the stage of production. For example, the meta-analytical approach of Rodrigues et al. [9] demonstrated that recovery of performance in post-weaned pigs was dramatically influenced by the recovery of feed intake, which was not observed in nursery and grower phases. The authors also reported a more abrupt and greater decrease in both growth and feed intake in younger, lighter pigs compared to older, heavier pigs. This may be explained by (1) the impact of gut fill, where newly weaned pigs take longer to reach maximum feed intake and (2) decreased growth potential due to limited feed intake in the immediate post-weaning phase [19]. The nutritionist needs to understand the many stressors concomitantly reducing post-weaning feed intake, including introduction to solid feed [20], abrupt depletion of passive immunity transfer from the dam [21,22], and the building of a new hierarchy with unfamiliar littermates [23]. Under this scenario, reestablishment of nutrient intake through higher nutrient supply or recovery of feed intake will be one of the most important governors for restoring weight gain [24].
Since different stressors may impact productivity at different stages of production, it is pivotal to understand how the exposure to pathogens and other stressors changes nutrient utilization and the associated decrease in performance. According to Pluske et al. [25], it is possible to manipulate the immune system through nutrition and other supportive strategies, including suppressing the presence and action of pathogens, breeding for improved resilience, and controlling the immune system to prevent overt response. With the increased trend for reduction in in-feed antimicrobial growth promoters’ usage, and the need for a more sustainable production system, a multidisciplinary approach will be needed to attenuate the negative impact of stressful agents, which are expected to be more harmful as pork production becomes more efficient [26]. The next sections will address the specific effects of individual challenge models on immune system activation and the associated effects on nutrient digestion and metabolism and build a relationship with growth performance measures. Subsequently, key nutritional strategies with proven benefits on the ability of pigs to cope with challenge models, including LP, AA-supplemented diets, FAA supplementation, dietary fiber level and source, diet complexity with inclusion of highly digestible ingredients, organic acids (OA), essential oils (EO), and natural compounds will be discussed.
Stimulation of the immune system results in higher concentration of circulating acute-phase proteins (e.g., C-reactive protein, serum amyloid A, haptoglobin, pig-MAP) [27], immune cell proliferation (e.g., clonal lymphocyte and monocyte differentiation), secretion of various molecules secreted by immune cells (e.g., cytokines, immunoglobulins), and lymphoid tissue hyperplasia [7] (Figure 1), with AA necessary to support the activated immune system. Although the amount of AA required for the activated immune system is not a substantial component of whole-body protein turnover, the AA profile required to support the immune response is drastically different than for muscle protein synthesis [28,29], resulting in a disparity between AA requirement and supply and reduction in utilization efficiency. Another significant expenditure of AA during inflammation is associated with the syntheses of endogenous antioxidants to cope with oxidative stress. Although the mitochondrial electron transport chain is a major site of free radical production, immune cells, such as phagocytes and monocytes, produce free radicals in the form of bactericidal agents (e.g., O2 and HOCl) or during the process of producing immune-signaling compounds (e.g., leukotriene and nitric oxide) [30]. For example, phagocytic cells, in a process called respiratory burst [30], consume large amounts of oxygen during phagocytosis and rapidly release superoxide (via the activation of NADPH oxidase) into the phagosome to kill bacteria. Likewise, a tripeptide γ-glutamyl-cysteinyl-glycine or glutathione (GSH) is a key, non-enzyme antioxidant in the body, and its turnover increases in response to ISS. Indeed, studies using isotope tracer have confirmed that the LPS-induced ISS accelerated transsulfuration pathway, converting Met to Cys and increasing Cys metabolism, mainly due to synthesizes of GSH in pigs [31].
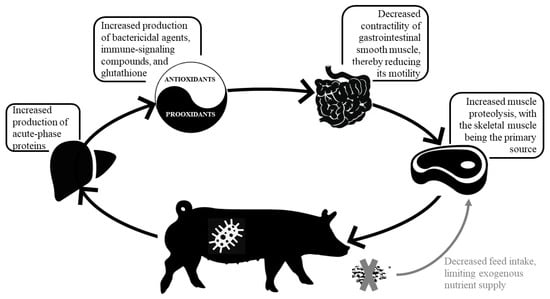
Figure 1.
Pigs subjected to stimulation of immune system experience increased production of acute-phase proteins (e.g., C-reactive protein, serum amyloid A, haptoglobin, and pig-MAP). Additionally, there is a marked syntheses of endogenous antioxidants to mitigate oxidative stress, including the production of immune-signaling compounds, the release of superoxide dismutase by phagocytic cells and the increased turnover of glutathione, which is a key, non-enzyme antioxidant in the body. Furthermore, a higher concentration of circulating pro-inflammatory cytokines decreases the contractility of gastrointestinal smooth muscle, thereby reducing its motility and sparing energy and nutrient expenditure for digestion. In attempt to minimize metabolic costs, pigs mobilize nutrients, particularly amino acids, from body reserves. This is mediated by proinflammatory cytokines and reactive oxygen species, which stimulate the IκB/NF-κB signaling pathway, eventually activating the ubiquitin proteasome system, and further triggering muscle proteolysis, with the skeletal muscle being the primary source. The reduced feed intake, which is mainly a result of upregulation of feed intake-lowering (anorexigenic) peptides and downregulation of orexigenic peptides, limits the exogenous nutrient supply and aggravates muscle proteolysis.
Therefore, it is reasonable to infer that traditional AA requirements are not applicable to immune-activated pigs because they were determined for optimal growth (protein accretion) under normal physiologic conditions [32]. Accumulating evidence has shown that dietary supplementation of specific AA can strengthen the pig’s immune system by minimizing body protein loss and accelerating its recovery [32]. This strategy seems to be more relevant for nursery pigs, who commonly undergo ISS post-weaning period.
3. Immune Challenges in Pork Production
3.1. Enteric Pathogen Challenge
The most common enteric diseases at pig farms are salmonellosis and swine enteric colibacillosis characterized by neonatal diarrhea and post-weaning diarrhea. The respective causal pathogens of these diseases are Escherichia coli and Salmonella spp. Of note, pathotypes F4+ and F18+ of enterotoxigenic Escherichia coli (ETEC) are the most predominant pathogens involved in post-weaning diarrhea [33]. Although colibacillosis is a major concern in nursery production, salmonellosis caused by infection of Salmonella enterica, particularly the serovar Typhimurium (ST), is widely observed throughout pork production [34,35]. In-feed antimicrobial growth promoters have been widely used to prevent or suppress the effects of enteric diseases, however, due to the ban on their use in many jurisdictions, animal nutritionists have made great efforts to seek alternative means to enhance gastrointestinal health while reducing antibiotic use. Although enteric pathogens have different pathotypes and pathogenicity, they all infect intestinal epithelial cells, triggering the action of the host’s immune system, generally leading to diarrhea [33]. For example, the ETEC flagella attach to the carbohydrate moieties of non-acid glycosphingolipids and glycoproteins present on the small intestinal epithelial cell layer, where they produce toxins (heat-labile and heat-stable toxins) (Table 1). These toxins activate the cyclic guanosine monophosphate and cyclic adenosine monophosphate systems, resulting in disruption of chloride and ion channels and, eventually, osmotic diarrhea [36]. Similarly, ST attaches to the intestinal epithelial cells and delivers a specialized set of effectors. Subsequently, ST injects Salmonella invasion protein into the intestinal epithelial cells, leading to cytoskeletal rearrangement in host cells, disruption of the normal epithelial brush border, and activation of the immune system [37]. The infected cells lose electrolyte absorption capabilities, thereby leading to diarrhea [38].

Table 1.
Mechanisms through which challenge models impact nutrient digestion and metabolism and growth performance.
Intestinal inflammation and disrupted integrity are common observations in pigs challenged with ETEC or ST. Inflammation and redox signaling systemically govern the fate and permeability of intestinal epithelial cells, which are relevant to intestinal integrity [39,40]. Pro-inflammatory cytokines trigger mitogen-activated protein kinase signaling and lead to apoptosis of enterocytes on the villus, mainly at the tip [41]. Furthermore, the expression of myosin light-chain kinase, which causes cytoskeletal contraction, is activated under inflammatory conditions, thereby decreasing intestinal permeability [42]. Pigs challenged with ETEC or ST are characterized by poor intestinal morphology (e.g., villus height, villus width, and crypt depth) and permeability, as well as reduced brush border enzyme activity and nutrient transporter expression compared to non-challenged counterparts [43,44,45]. Indeed, ETEC-challenged pigs have lower ileal digestibility of nutrients (e.g., AA) than sham-challenged control pigs [46].
As enteric pathogens eventually stimulate the systemic immune system, the metabolic pathways change. These changes are initiated by the immediate increase in concentration of acute-phase proteins (e.g., haptoglobin) and immunoglobulins following pathogen exposure [10,44]. As a result, the requirements for several AA such as Thr, Trp, and sulfur AA (SAA), may be elevated for optimal growth performance and immune response [47,48,49].
3.2. Respiratory Pathogen Challenge
As a result of intensive, confined systems of modern pig production, respiratory diseases continue to be a cause of concern in pork production, commonly resulting in impaired weight gain and feed efficiency, poor animal welfare, and increase in medication costs [74]. There are several reports from different parts of the world indicating significant prevalence of pneumonia (20 to 80%) and pleuritis (4 to 60%) in slaughtered pigs, which indicates an important impairment of the respiratory tract throughout the productive life [51,52,53] and potentially significant productive costs of immune stimulation. The term porcine respiratory disease complex (PRDC) is often used to address the several bacteria, viruses and mycoplasmas involved in the etiology of respiratory diseases [75,76]. The most important agents are Actinobacillus pleuropneumoniae, Mycoplasma hyopneumoniae, Pasteurella multocida, porcine circovirus type 2, porcine reproductive and respiratory syndrome virus (PRRSV), porcine respiratory coronavirus, and Salmonella choleraesuis. The growth performance response of pigs to respiratory pathogens will be dependent on the agent involved. Two recent meta-analyses showed similar results related to severity of respiratory challenge [8,9]. In general, respiratory diseases were less detrimental to performance than the other immune challenges, which may possibly be due to a greater degree of compensatory growth following respiratory infections [77]. Moreover, many respiratory pathogens produce an immune response that is more likely to be contained within the affected tissues (e.g., neutrophilia and damage to cilia [54]), resulting in more minor clinical response and a reduction in a significant response in growth performance. However, subclinical infections can still result in significant economic consequences. For example, an economic analysis indicated that an increase in lung lesions above 15.1% resulted in a loss of up to $6.55 per pig at slaughter.
Among respiratory pathogens, there is more evident information in the literature to show that PRRSV is the most detrimental to performance. For example, body weight of weaned barrows was decreased by 5% in PRRSV-infected compared to non-infected pigs [78]. This is in agreement with Schweer et al. [12] who reported a 10% reduction in weight gain, 6% reduction in feed intake, and 7% reduction in feed efficiency in PRRSV-infected pigs compared to non-infected pigs through a market weight of approximately 128 kg. In order to assess whether the reduction in performance is driven by decreased feed intake and/or nutrient metabolism, Helm et al. [57] compared PRRSV naïve, ad libitum fed, PRRSV-inoculated, ad libitum fed, and PRRSV naïve, pair-fed to the PRRSV-inoculated pigs’ daily feed intake. Growth performance and feed efficiency were negatively impacted by PRRSV infection compared to the pair-fed group which indicates modification in nutrient metabolism. Specifically, markers of skeletal muscle synthesis (e.g., phosphorylation of protein synthesis markers) were reduced while liver glycogen was more depleted in PRRSV-infected compared to the pair-fed group. This also agrees with a 13% reduction in both whole-body protein accretion when pigs are infected with PRRSV, which reduced carcass lean yield [12]. A recent study by Schweer et al. [79] further demonstrated that PRRSV infection dramatically reduces weight gain (30%) and feed intake (26%) which was not accompanied by a decrease in the apparent total tract digestibility (ATTD) of AA and energy. This provides evidence that PRRSV infection reduces growth performance through a combination of reduced feed intake and decreased nutrient utilization.
3.3. Degradation of Sanitary Condition
While it is important to understand the effects of individual pathogens on pig immune status and performance, in commercial settings, pigs are more likely to be exposed to a variety of pathogens. This exposure is the result of an increase in environmental pathogen load. Moreover, the response of pigs to pathogens is complicated in commercial settings through the use of antimicrobial growth promoters and vaccination protocols which reduce pathogen load and enhance herd immunity to pathogens and individual pig ability to avoid infection. Experimentally, sanitary challenge represents an increase in pathogen exposure via absence of cleaning and disinfection protocols, reduction in vaccination, reduction in antibiotic use, or a combination [2,15,80]. Pigs are exposed to various stressors, broadly classified as non-biological stressors (e.g., noxious gases, dust) and biological stressors (e.g., bacteria, virus). The sanitation challenge aims to mimic the practical commercial swine barn environment and mildly activate the pig’s immune system and is thus often regarded as subclinical compared to studies of other pathogen, toxin, and stress challenges. Nevertheless, this model has been advantageously adopted by studies that tested diet interventions for pigs raised in commercial environments [63] and in which nutrient requirements of pigs housed in experimental or commercial environments were compared [15]. Because the sanitation challenge model can be applied without special facility or inoculum preparation, the experimental design (e.g., treatments, replications) is more flexible than that of other challenge models. Due to this design flexibility, the model can be applied to a large number of pigs, which increases the statistical reliability of growth performance (weight gain and feed intake).
Because LSC are usually created by the accumulation of manure in the pens, the pigs are more likely to ingest pathogens from the manure orally. Thus, gut health indicators, such as intestinal integrity and microbial composition, are typically investigated in sanitary challenge studies [61,62]. Furthermore, pathogens or dust can be transferred into the body through the nasal tract, so respiratory health indicators are also of interest in these studies [1]. Indeed, LSC increased the incidence of respiratory tract inflammation, possibly because of the increased loads of respiratory pathogens [1]. Additionally, LSC are often characterized by the buildup of noxious gases, such as H2S and NH3, as well as dust [80,81], which possibly irritate the respiratory tract. Pigs raised in LSC generally have lower body weight gain compared to those raised in high sanitary conditions [1,60,64,81]. However, whether this low body weight gain is accompanied with low feed intake has been argued. Moreover, it has been consistently reported that intestinal inflammation, oxidative stress, and disturbed intestinal integrity are consequences of housing pigs in LSC. For example, LSC also upregulates the expression of inflammatory cytokines, such as interferon (IFN)-γ, IL-1β, IL-6, IL-10, and TNF-α, in mesenteric lymph nodes and the ileum [63]. Additionally, shorter villus height in the small intestine have been observed in pigs reared in LSC compared to those reared in HSC [64,81]. The change in intestinal integrity may explain the lower ATTD of dry matter, nitrogen, and gross energy in pigs housed in LSC [1,2]. Moreover, the physiological changes caused by LSC alter nutrient and energy metabolism, leading to shifts in AA and maintenance energy requirements. Kahindi et al. [80] reported that the standardized ileal digestible (SID) requirement of SAA increased by approximately 6–10% based on villus height and plasma urea nitrogen responses in weaned pigs. Similarly, Jayaraman et al. [81] estimated that SID Trp requirement is 4% higher when pigs are housed in LSC compared to HSC. Furthermore, van der Meer et al. [2] reported that pigs housed in LSC have 8% higher fasting heat production—the greatest portion of maintenance energy—than those in HSC.
As anticipated, LSC is able to modify the intestinal microbiome composition and functionality. A change in the abundance of lactic acid-producing bacteria has been noticeable in sanitary challenge studies. For example, Cho et al. [82] reported that LSC decreased the fecal abundance of the Lactobacillaceae family. Their findings were supported by Montagne et al. [61], who found lower Lactobacillus counts (colony forming unit) in feces of pigs reared in LSC. In another sanitary challenge study, Waititu et al. [63] reported lower Bifidobacterium spp. abundance in the cecum and colon, but no difference in Lactobacillus. Another study used metagenomic assay [62] and showed that the Megasphaera genus, known to be involved in lactate fermentation, was lower in the colon digesta of pigs reared in LSC. A reduced number of Megasphaera may be associated with lower concentration of lactic acid in the lumen. A general agreement exists that LSC results in greater protein fermentation. Cho et al. [82] reported that LSC increases the production of branched chain fatty acids in feces. Because branched chain fatty acids are produced from branched chain AA, an increase in the production of these fatty acids possibly reflects an increase in protein fermentation. Recent studies have consistently reported that LSC reduce butyrate concentrations in the colon [62], and feces [82]. Their findings were consistent with microbial assays in which the abundance of butyrate-producing bacteria, such as Clostridiales family XIII Incertae Sedis [82] and Clostridium IV and XIVa [63], decreased in LSC-challenged pigs. In contrast to butyrate, sanitary degradation generally increases total short chain fatty acids (SCFA) and volatile fatty acids (VFA) concentration. te Pas et al. [62] reported that LSC increases the abundance of Lachnospiraceae family in the colon digesta, which are known to be one of the major microbes involved in SCFA synthesis from complex polysaccharides. Montagne et al. [61] also reported an increase in VFA concentrations in the feces of nursery pigs housed in LSC. The authors postulated this phenomenon as the “hygiene hypothesis”, meaning that a clean environment can impede the optimal development of the immune system and microbiome establishment in young animals. However, LSC might decrease foregut digestion and increase undigested nutrients, thereby modifying hindgut fermentation patterns. Therefore, further studies are necessary to elucidate changes in fermentation patterns and microbial metabolites in relation to sanitary conditions.
Although the sanitary challenge model can be applied to nutritional studies, the model has been criticized due to the lack of reproducibility. While other immune challenge models can quantify the stimulation intensity (e.g., dose), quantifying sanitation level is difficult because the sanitary challenge model generates various stressors in different facilities and environments. Swine barns have different predominant pathogens and pathogenesis, causing the intensity of the stressors to vary depending on location. Furthermore, farms have different floor systems (e.g., slat, pit) which may affect the stressors’ intensity as this directly influences the quantity of manure accumulation and noxious gas exposure. These factors lead many studies to report no differences in growth performance or ISS between sanitary conditions. Therefore, to improve the reproducibility of the sanitation challenge model, details of experimental procedures and environments, including air quality, ventilation operation, ambient temperature, humidity, manure source and management, pen cleaning frequency, pen size, and floor design, should be provided.
3.4. Bacterial Lipopolysaccharide
Lipopolysaccharide is a component of the outer membrane of Gram-negative bacteria implicated in the pathogenesis of many disease states, causing endothelial cell injury and dysfunction. The LPS possesses a highly proinflammatory characteristic, which triggers the upregulation of cytokines, adhesion molecules, and tissue factor [83]. The model has been largely used in pigs to mimic systemic inflammation [68] and causes significant reduction in body weight gain [67,84]. It should be noted that the period after which LPS is administered may influence the response in growth performance of pigs, since tolerance may occur following multiple injections. A recent meta-analysis conducted by Rodrigues et al. [9] revealed a greater reduction in average daily gain in studies using LPS compared to studies using other challenge models (e.g., respiratory pathogens, low sanitary condition, environmental stress). This is consistent with sepsis and septic shock entailed by LPS administration mainly due to the overt cytokine production [85,86]. Among them, IL-6 circulating concentration, which is associated with fever, has been reported to increase after LPS injection [70,87]. Pastorelli et al. [8] reported in a meta-analysis that the reduction in performance during a LPS challenge was due to a depression in feed intake and not to increased maintenance requirements. More recently, these findings were contradicted by another meta-analytical approach which statistically contrasted different challenge models and showed that maintenance requirements had a substantial contribution to the decreased performance in LPS-challenged pigs [9]. Indeed, altered AA requirements have been demonstrated in multiple studies in LPS-challenged pigs, including Met [69], Thr [18], and Trp [65], which indicates that nutrient maintenance requirements/utilization efficiency play an important role in the observed reduction in performance.
Impairment in growth performance has been reported in weaned pigs, along with fever and increased production of major acute-phase proteins and cytokines (e.g., haptoglobin, tumor necrosis factor-α and interleukine-1 beta [71]. This is consistent with the findings of Xu et al. [72], who reported an increased plasma concentration of cortisol, prostaglandin E2 (PEG2), IL-6, TNF-α and IL-1β dramatically after LPS administration, which corroborates the systemic commitment after challenge. A recent study revealed the clear reduction in nutrient utilization efficiency in LPS-administered pigs, where not only growth performance but also the concentration of most microelements in feces and the expression of most microelement transport genes in the mucosa of the gastrointestinal tract were decreased after injection [73]. It is important to consider that LPS is a challenge model mainly used to mimic systemic activation of the immune system. However, since many pathogens and antigens are able to entail systemic response, it may be inferred that nutrient requirements are increased when there is a load to the production of immune cells and co-factors.
4. Nutritional Strategies for Improved Health Status
4.1. Low Protein, Amino Acid-Supplemented Diets
It is well known that high dietary protein (HP) content may have detrimental effects on gut health in pigs [88,89], which is presumably attributed to the indigestible fraction, available for microbial fermentation (i.e., protein not absorbed in the small intestine) and the potential for a high dietary protein level to support the proliferation of pathogenic bacteria, such as ETEC, by increasing the pH of the gut through the high buffering capacity of protein [90]. Studies have reported pro-inflammatory effects entailed by protein fermentation metabolites (e.g., branched-chain fatty acids, ammonia, biogenic amines, hydrogen sulfide, and phenolic and indolic compounds) including compromised colonic epithelial cell structure and metabolic functions, thinning of the mucus barrier, and increased colonic permeability [91,92,93] In this sense, studies have established a relationship between feeding HP diets and the incidence of post-weaning diarrhea in pigs [46]. Furthermore, it has been amply demonstrated that LP diets may improve gut health outcomes by suppressing the proliferation of pathogenic bacteria while promoting those with beneficial effects [94,95,96]. Therefore, the recommendation is that nutritionists should feed LP diets that are supplemented with crystalline AA to meet requirements for essential AA. It is thought that these diets reduce the amount of substrate (i.e., undigested protein) for pathogenic bacteria and production of harmful metabolites, leading to improved gut health and function in the postweaning phase [91].
In a recent review, Zhang et al. [89] reported that feeding HP diets evoked a shift in gut microbial composition favoring nitrogen-utilizing communities, including pathogenic groups, which is highly associated with incidence of diarrhea. Moreover, protein fermentability increased the concentration of (primarily) biogenic amines in the gut leading to impaired intestinal morphology, increased gut permeability, and increased pro-inflammatory cytokine concentration. While some studies have reported attenuation of diarrhea in post-weaned piglets fed LP diets [96,97], there is no consensus across studies. For example, Rodrigues et al. [10] reported minimal effects of dietary protein content when ST inoculated pigs were fed 16% or 20% protein diets. Conversely, Pollock et al. [98] reported that a HP diet aggravated the disturbances in the gastrointestinal environment entailed by ETEC challenge. This indicates that there are factors other than simply total dietary protein content (e.g., indigestible content, protein type) involved in the aggravation of post-weaning diarrhea. The contribution of protein fermentation metabolites to the negative effects on gut health are not fully elucidated [99] and there is still a lack of consensus on methodologies to evaluate and classify intestinal health [100].
It is important to highlight that the reduction in dietary protein must be accompanied by the supplementation of crystalline essential AA (EAA). For example, Yu et al. [101] fed weaned pigs diets with 20, 17, and 14% protein supplemented with Lys, Met, Thr, and Trp only. After a 45 d feeding period, the authors reported small intestine atrophy (e.g., decreased villus heights and lower ratios of villus height to crypt depth) and impaired pepsin activity in the stomach, suggesting an AA deficiency for intestinal development and enzyme activity. Likewise, Chen et al. [102] fed pigs a normal protein (18%), LP (15%), or extremely LP (12%) diet for 30 days and reported that the moderate protein restriction only positively modulated bacterial communities, increased the expression of tight junction proteins and enhanced epithelial cell proliferation in ileum. Spring et al. [103] further demonstrated the importance of supplementing very LP diets with adequate amount of crystalline AA. Supplementation of branched-chain AA attenuated the impairment in growth performance, energy balance, and metabolic and gut microbiome profile due to protein restriction (e.g., [104]).
4.2. Dietary Fiber Level and Source
Dietary fiber is a general classification which includes a broad spectrum of oligosaccharides and starch resistant to proximal intestine hydrolysis, as well as non-starch polysaccharides such as pectin, cellulose, hemicellulose, β-glucans and fructans. Subsequently, fiber sources can be classified according to their solubility, viscosity, physical structure, and water-holding capacity, which will combinedly determine their physiological role. In pigs, soluble dietary fiber is fermented primarily in the colon producing gases and several physiologically active by-products [105]. Insoluble fiber, in turn, generally increases diet bulkiness due to its metabolic inert characteristic [105]. Moreover, the well-known limited feed intake capacity of piglets may limit the ability of young animals to digest high fiber diets properly, which can be detrimental to growth performance and feed efficiency. For example, fiber may act as an antinutritional factor by decreasing nutrient digestibility and increasing endogenous mucin secretion, further aggravating the amount of undigested protein reaching distal parts of the gut [93] and decreasing AA availability for growth [106]. This is the reason why fiber levels are kept generally low in starter and nursery diets, as opposed to grower and finisher diets, where feed intake and digestion are not limiting factors.
In a recent review, Williams et al. [107] explored the categorization of dietary fibers into soluble or insoluble and discussed the importance of moving beyond this simple stratification. Basically, it is pivotal to understand how the microbial populations inhabiting the gut respond to a variety of fiber sources and levels, and how recommendations must be addressed in terms of fiber fermentability, rather than only solubility. In this sense, the inclusion of fermentable fiber sources and/or lowering dietary protein content can be used as strategies to potentiate beneficial metabolites while suppressing negative metabolites [88,91,108]. This is partly explained by a prioritization of energetic over protein fermentation by gastrointestinal microbes and an increased amino acid and ammonia incorporation into the microbial biomass [92,93,109]. However, it should be reiterated here that feeding soluble and rapid fermentable fiber sources during the immediate post-weaning period, especially weaning age is decreased and the sanitary condition around weaning is suboptimal, may lead to negative results, again, due to very limited feed intake capacity [110]. After two weeks post-weaning, when it is expected an improved adaptation of piglets to solid feed, a gradual inclusion of soluble and fermentable fiber may be advantageous for enhancing fermentation of nutrients and improving absorption of short-chain fatty acids in the large intestine mucosa.
Specifically, and unlike with protein, the inclusion of fermentable fiber in swine diets can have positive effects on the gastrointestinal environment as reviewed by Jha and Berrocoso [111]. The mechanisms underlying the positive effects of fermentable fiber in pigs is related to the formation of end products of fermentation (acetate, propionate, butyrate) and include improvement in colonic barrier function and immune/metabolism-related gene expression [112], maintenance of microbial community homeostasis [113], and attenuated release of inflammatory intermediates [114]. Regarding gut microbial populations, it has been suggested that dietary fermentable fiber improves microbiota stability and improves its diversity with promotion of proliferation of potentially beneficial microorganisms, including Lactobacilli and Enterococci [88,115,116], and suppression of potentially harmful populations, including Clostridia spp. and Escherichia coli [109,117]. Furthermore, evidence exists to show that the physical form of a fiber type may also influence its ability to mitigate the pathogenesis of Escherichia coli in swine. For instance, Molist et al. [118] reported that supplementing a nursery pig diet with coarse (1088µ) wheat bran prevented ETEC colonization of the small intestine and reduced severity of diarrhea compared with supplementing a finely (455µ) ground wheat bran in ETEC-challenged pigs. It is important to note that the impact of dietary fiber on host health and pathogen susceptibility have not been consistent across studies, possibly due to differences in fiber properties as well as fermentability potential of different sources [119,120]. Of note, feeding a high fiber diet may potentiate mucus secretion and epithelial cell sloughing which damages the gut architecture [121,122,123] and may increase animal susceptibility to pathogens. Indeed, Wellington et al. [49] reported decreased growth performance in Salmonella-challenged pigs fed high fiber diets, where fiber consisted of a mixture of soluble and insoluble sources. Moreover, in a subsequent study, Wellington et al. [124] showed improvements in barrier function of LPS-injected pigs when fed high fiber diets. Likewise, feeding non-starch polysaccharides to growing pigs increased gastrointestinal water secretion after infection with swine dysentery [125]. Finally, while the inclusion of beta-glucan in the diets improved growth performance of nursery pigs, there was an increased susceptibility to Streptococcus suis infection [126].
4.3. Diet Complexity
Diet complexity has long been discussed in swine nutrition because of its economic significance in nursery pig production. Diet complexity generally refers to dietary composition, where higher diet complexity indicates a greater number of ingredients in the diet and, generally, the inclusion of animal-based ingredients (e.g., fish meal, blood meal, plasma meal, and dairy products) as well as feed additives and antibiotics for the purposes of creating a diet with greater nutrient availability, reduction in anti-nutritional factors, and reduction in pathogen load. Animal-based ingredients, such as fish meal, whey, and blood meals are good protein sources with high-quality AA profiles that lack antinutritional factors (e.g., antigenic compounds and non-starch polysaccharides) [127,128]. Furthermore, the functionality of fish and blood meals (e.g., particularly spray-dried porcine plasma) has been reported in nursery pigs as enhancers of health status and growth performance [129,130]. The inclusion of dairy products, such as lactose, whey powder, whey permeates, and skim milk powder, provide a source of lactose, enhance palatability, and are thought to ease the transition to non-milk-based diets post-weaning [131].
Animal-based ingredients increase both the complexity and feed cost of the nursery pig diet. In addition, there has been recent social pressure to reduce the use of animal-based ingredients in livestock feed, with plant-based diets being considered by the public to be healthier and more welfare-friendly. Thus, efforts have been made to simplify diets during the nursery stage without compromising growth performance and productivity. In general, pigs fed a conventional, complex diet have greater body weight gain and feed intake in the post-weaning period than those fed a simple diet [131,132,133]. Furthermore, higher nutrient digestibility and daily energy intake have been observed in pigs fed complex diets compared to pigs fed a simple diet [134]. Interestingly, Wang et al. [135] reported a more balanced protein metabolism, as determined by urea nitrogen concentration, when pigs were fed a complex compared to a simple diet. However, these benefits do not last throughout the subsequent production period. For example, nursery diet complexity (simple vs. complex) does not alter carcass characteristics, including longissimus muscle quality, loin meat quality, and ham composition [132,136,137]. Therefore, lagged growth in nursery pigs resulting from a simplified, low-quality nursery diet can be compensated for during the re-alimentation period with improvements in feed efficiency, a phenomenon commonly referred to as compensatory growth.
Attempts have been made to mitigate the negative effects of a simple diet on the immune system and intestinal microbiota. Koo et al. [134] showed that dietary feed enzyme supplementation to a simple diet can be beneficial in terms of nutrient and energy digestibility and intestinal morphology. In another study by Koo et al. [50], supplementation of 0.1% of L-Thr to the simple diet increased the number of jejunal goblet cells wherein mucins are produced. The authors postulated that this benefit led to a reduction in ammonia nitrogen concentrations in the jejunum and downregulated intestinal inflammation, thereby increasing the villus height to crypt depth ratio. However, the supplementation of L-Thr to the simple diet did not restore the increased IL-6 in the serum—a biomarker of systemic inflammation—to the levels typical of a complex diet. Therefore, to successfully replace a complex diet with a simple diet in commercial nursery production, nutritional strategies that can prevent systemic metabolic changes should be studied.
4.4. Functional Amino Acids
As previously stated, when the immune system is stimulated after an injury or infection there is a prioritization of AA utilization for the immune response at the expense of growth [138]. Moreover, the reduced feed intake, which commonly occurs as a result of ISS, exacerbates the reduction in supply of AA, further impairing lean tissue deposition [28,139,140]. In this sense, it is reasonable to infer that a significant amount of muscle protein, as the largest body AA pool, will be mobilized in order to meet changing requirements associated with the immune response [138]. However, as shown by Reeds et al. [138], there is a notable difference between the AA profile of muscle protein and the profile of many important acute-phase proteins involved in the immune response. This indicates an imbalance which, unless supplemented through the diet, will trigger significant whole-body AA catabolism and reduction in body protein deposition [138,141]. For example, pigs fed a SAA-free diet had lower intestinal Cys concentration, associated with reduced jejunal goblet cell number, which may indicate insufficient secretion of Cys-rich mucins involved in the intestinal innate immune response [142,143]. Albumin is a Cys-rich acute-phase protein, with production of 1 g of albumin requiring approximately 6 g of muscle protein breakdown in the absence of another source of Cys. Likewise, neonates, highly dependent on the innate defenses of mucus [144], show impaired mucin production and gut function when fed a Thr-deficient diet [145]. Traditionally, nutritionists have defined AA essentiality based on growth performance outcomes and there is a lack of information in the literature regarding the role they perform on the modulation of immune response, anti-oxidative defense, and recovery from injuries [146]. However, there is growing interest in the ‘functional’ roles AA have beyond their role as constituents of lean gain [32].
Liu et al. [147] conducted a dose–response study with 21 d old piglets investigating the ratio between SAA and Lys, testing the content of 70, 85, 100, 115, or 130% of the SAA: Lys ratio recommended for growth. The authors reported a downregulation of genes related to inflammation (e.g., TNF-a, transforming growth factor (TGF)-β, and IL-1β) as SAA intake increased from 0.63%, suggesting a functional role of SAA even considering weanling, presumably healthy piglets. Similarly, Yan et al. [148] fed weanling pigs a low (0.53%) or high (0.85%) SAA diet for one week and clearly showed that the increased intake enhanced jejunal cell proliferation and function, mainly through improved antioxidant capacity, and Wnt/β-catenin and mTOR signaling pathway. Weanling pigs fed increasing levels of SID Trp: Lys ratios (16.1%, 18.6%, 20.3%, 22.9%, and 24.6%) and challenged with Escherichia coli K88 had optimal performance when fed 21% and showed increased expression of the anti-inflammatory IL-10 with increasing ratios [81].
Recent studies have shown that endogenous (kynurenine, serotonin, and melatonin) and bacterial (indole, indolic acid, skatole, and tryptamine) Trp metabolites play an important role in gut microbial composition and metabolism, immune response of the host, and host-microbiome interaction [149]. Moreover, it has been highlighted that stress and disease, including irritable bowel syndrome and inflammatory bowel disease, directly alter Trp metabolism and disturb the Trp–microbiome–immune system interaction in the gut. It has been shown that higher Trp intake resulted in a number of beneficial effects in the piglet gastrointestinal tract, namely: (1) enhanced microbiome diversity, (2) decreased abundance of opportunistic bacteria, and (3) increased mucosal IL-8 mRNA level, and zonula occluden (ZO)-1 [150]. Koo et al. [50] reported an increased villus height and goblet cell density, and a higher expression of jejunal occluden and downregulation of IL-6 in pigs fed diets with 115% compared to 100% of the requirement for SID Thr. This is in line with recent findings showing that supplemental Thr (120% of requirements) improved growth performance in ST pigs fed low fiber (LF) diets [49], and increased fecal mucin output in ST pigs fed high fiber (HF) diets [124]. There is also recent evidence showing that the combination of AA can perform functional properties in pigs subjected to immune challenge. Rodrigues et al. [10] fed a basal or a FAA (Thr, Met, and Trp at 120% of requirements) profile to 14 kg pigs for 14 d, divided in pre- and post-inoculation with either saline or ST. Pigs inoculated with ST and fed the FAA profile had greater weight gain, improved antioxidant defenses, attenuated acute-phase response, and reduced pathogen shedding and colonization. In a subsequent study, the authors reported that the previously positive results were enhanced when pigs were fed the FAA profile for a longer period (e.g., 2 weeks) before ST inoculation, with no effects of diets during the pre-inoculation phase [11]. These results suggest that immune status, gut health, and overall pig robustness may be improved with the supplementation of FAA [151,152,153], which becomes particularly important in diseased pigs. Interestingly, a recent study revealed that FAA supplementation triggered a positive response in mitigating the effect of enteric disease challenge in normal birth weight (NBW), but not low birth weight (LBW) pigs [154]. Additionally, FAA supplementation partially attenuated the detrimental effects of plant-based (PB) diets on the response of pigs to ST challenge, while FAA supplementation had minimal effects in ST-challenged pigs fed animal-based (AB) diets [155].
4.5. Organic Acids
Among various feed additives, OA have been vigorously researched in swine nutrition in relation to gastrointestinal health. Organic acids can be broadly classified into three categories based on the carbon chain: SCFA (e.g., formic acid, acetic acid, propionic acid, butyric acid), medium chain fatty acids (MCFA; e.g., caproic acid, caprylic acid, capric acid, lauric acid), and tricarboxylic acids (e.g., citric acid, fumaric acid, and malic acid). Most importantly, OA possess bacteriostatic and bactericidal actions. Undissociated forms of OA can diffuse across the bacterial cell membranes, where they are dissociated inside the cells and release H+ ions, thereby disrupting the acid-base balance and vital metabolic pathways of microbes [156]. However, the antimicrobial efficacy of OA against pathogens varies depending on their physicochemical properties (e.g., pKa, lipophilicity, and solubility) and the target microbes (e.g., the structure of cell walls or membranes) [157]. For example, SCFA have shown strong efficacy against Gram (−) bacteria, including E. coli and Salmonella spp., while MCFA have shown strong efficacy against Gram (+) bacteria, such as C. perfringens and Steptococcus spp. [157,158]. Therefore, combining different OA may be critical for maximizing efficacy against different pathogens. The present review focuses on the effects of dietary OA supplementation in immune-challenged pigs. This discussion will help introduce OA practicality on farms, in which the environment is more stressful to pigs than the experimental environment.
Unlike the straightforward efficacy found in vitro, in vivo studies have shown controversial results in ST or ETEC-challenged pigs. Gebru et al. [159] reported that dietary microencapsulated OA (citric acid, fumaric acid, malic acid, and phosphoric acid in a 2:2:1:1 combination) supplementation at 0.2% decreased the fecal shedding of ST and improved growth performance for the 4-week experimental period in growing pigs (initial BW: 38.7 kg). Similarly, Calveyra et al. [160] found that OA supplementation decreased the number of Salmonella spp. in the feces of growing pigs. In contrast, other studies in nursery pigs failed to find the benefits of OA supplementation alone (0.8%, 1%, or 2.58 mL/L of water) on the microbial profile or ST shedding in ST-challenged nursery pigs [161,162,163]. Interestingly, Fabà et al. [163] reported that a combination of OA with mannan-rich hydrolyzed copra meal or fermented rye reduced the shedding of ST in ST-challenged pigs. This may suggest that OA alone are not sufficient to control the pathogenicity of ST in nursery pigs. On the other hand, the efficacy of OA against ETEC in nursery pigs is much more apparent. Dietary OA supplementation reduced the incidence of diarrhea or fecal scores in ETEC-challenged pigs [164,165]. The immunomodulatory effects of OA have also been observed in ETEC-challenged pigs. Organic acids supplementation decreased concentration of pro-inflammatory cytokines, such as IL-1β, IL-6, TNF-α, and IFN-γ, in plasma to levels comparable to antibiotics supplementation [166]. Similarly, Jiménez et al. [167] reported that OA supplementation decreased the number of inflammatory cells in the jejunal and ileal lamina propria, which had been elevated by ETEC inoculation.
Further benefits of OA on intestinal and hepatic redox status were reported based on an increase in GSH and ferric-reducing ability potential and a decrease in thiobarbituric acid reactive substances (TBARS) concentrations in the intestine or liver of pigs fed an OA-supplemented diet [167]. However, the effect of OA on intestinal microbial composition seems to be inconsistent. For example, Han et al. [168] performed 16s rRNA sequencing with ileum digesta samples and showed that, based on the alpha diversity indices (Chao1, ACE, and Shannon indices) and shift in Firmicutes and Proteobacteria abundance, pigs fed an OA-supplemented diet had a more diverse and stabilized microbiome composition than control diet-fed pigs. In contrast, Ren et al. [166] failed to find the benefits of OA mixture supplementation on fecal counts of total coliforms and Lactobacillus. Stensland et al. [169] reported no effects of OA supplementation on the counts of total E. coli, F4 E. coli, Enterobacteriaceae, and Lactobacillus spp., but they did find an increase in total VFA concentration in the feces of pigs fed an OA-supplemented diet. Differences in ETEC inoculation dose, sampling time, and analysis methods may have contributed to the contradictory results among these studies.
4.6. Plant Secondary Metabolites
Due to the ban on antimicrobial growth promoters, growing interest in organic livestock farming has accelerated the use of plant secondary metabolites (PSM), often called phytogenic feed additives. Plants have evolved to adapt to the environment, which for some plants involves the production of special metabolites that serve as a defense mechanism against exogenous stressors such as germs and oxidative damage [170]. Various PSM can be beneficially ingested by animals, providing immunomodulatory, antioxidative, anti-inflammatory, antifungal, antiviral, antibacterial, and anti-diarrheal effects [171]. Plant secondary metabolites are broadly classified into terpenes (e.g., carvacrol, thymol), phenolics (e.g., eugenol, resveratrol, quercetin, tannins), N-containing compounds, and S-containing compounds (e.g., alliin and allicin). Phenolics are further divided into polyphenols (e.g., tannins and flavonoids), phenolic acids (e.g., benzoic acid, ferulic acid, gallic acid, vanillin), and miscellaneous (e.g., lignans, resveratrol) [170].
In pig nutrition, various sources of PSM have been studied. Generally, PSM have been delivered through plant extracts and agro-industrial by-products (e.g., grape pomace). These products usually contain a mixture of PSM and have reduced risk of causing the emergence of resistant bacteria due to their compositional and chemical complexity [172]. However, because their bioactive components and profiles change depending on the plants’ maturity, harvesting time, and weather, producing qualitatively consistent products is difficult [173], which creates challenges in diet formulation. Advanced technologies have allowed some PSM to be chemically synthesized and these products are commonly called nature-identical compounds [174]. These synthesized compounds (e.g., thymol and vanillin) can be precisely supplemented to swine diets and synergically combined with other feed additives.
Essential oils, which are either terpenes or phenolics, are concentrated, highly volatile hydrophobic liquids. They are usually extracted from plants, but several naturally identical EO are commercially available. Because of their phenolic ring, or capacity to disturb microbial membranes and intracellular homeostasis, EO usually show antimicrobial, anti-inflammatory, and antioxidative properties [175]. Dietary EO blends, such as thymol and cinnamaldehyde, improve antioxidant capacity by enhancing antioxidant enzymes, including superoxide dismutase and catalase in the serum or mucosa of weaned piglets [176,177]. Improved digestibility of dry matter, gross energy, crude protein, and AA has been consistently reported in pigs fed EO blend-supplemented diets [178,179,180]. In a study by Ruzauskas et al. [181], pigs received 3 tablets daily, each of which contained 986 mg of oregano extract, 3 mg of peppermint EO, and 7 mg thyme. These pigs had a higher abundance of bacteria considered probiotic, such as Lactobacillus, Bacillus, and Bifidobacterium, in the gut (i.e., ileum, cecum, and colon) compared to control pigs that did not receive EO. Similarly, a reduction in E. coli or Enterobacteria counts and increase in Lactobacillus counts in feces [178] and the jejunum and cecum digesta [182] was found in pigs fed EO-containing diets compared to pigs fed a control diet. The microbial modulation by dietary EO possibly leads to a reduction in the incidence of post-weaning diarrhea [178,180,182]. However, the anti-diarrheal effect seems to be dependent on the dose. Cairo et al. [182] reported that 0.15% of red pepper EO reduced diarrhea incidence by 43% compared to a control group, but 0.1% supplementation increased the incidence by 21%.
Dietary EO supplementation to a swine diet has been often tested in combination with other compounds, particularly OA, as EO and OA have better efficacy against Gram (+) and Gram (−) bacteria, respectively [44,157,183]. Apart from EO, plant extracts as a source of polyphenolic compounds and polyphenol-rich ingredients have been investigated frequently. Coddens et al. [184] reported that cranberry extracts rich in proanthocyanin could inhibit the adhesion of F4+ and F18+ E. coli on the ileum in vitro. They found that supplementing the diet (0.1%) and water (1 g/L) with cranberry extracts reduced the diarrhea score and F18+ E. coli shedding in nursery pigs. Similarly, Xu et al. [72] found that 250 mg/kg of holly polyphenols elevated jejunal and disaccharides (sucrase and lactase) activity, upregulated tight junction proteins in the ileum (claudin-1 and occludin), and decreased pro-inflammatory cytokine contents in the plasma. Furthermore, holly polyphenol supplementation increased Lactobacillus in the cecum and colon, regardless of LPS challenge, and it restored the cecal abundance of Prevotella, a major dietary fiber fermenter, which is suppressed by LPS administration.
Grape by-products can also be beneficially fed to pigs as a polyphenol source [185]. For example, Kafantaris et al. [186] reported that the dietary inclusion of fermented grape pomace at 48.5% enhanced the antioxidant defense system based on the levels of GSH, total antioxidant capacity, TBARS, and protein carbonyls in various organs, including the liver and pancreas of nursery pigs. Additionally, the inclusion of grape pomace beneficially modified the fecal bacteria composition by increasing Bifidobacterium and lactic acid bacteria counts and reducing Enterobacteriaceae counts. Gessner et al. [187] found that supplementation of grape seed and grape marc meal extracts in a nursery diet at 1% suppressed the activity of NF-κB, an inducible transcription factor for inflammatory responses and the expression of inflammatory cytokines in the duodenal mucosa of nursery pigs.
Nevertheless, the efficacy of PSM follows the hormesis concept, meaning that an excessive amount of PSM may serve as an antinutritional factor, stimulating the immune system and reducing nutrient digestibility. Therefore, dietary supplementation levels should be optimized for the promising PSM before it is commercialized.
5. Conclusions
Swine production systems expose pigs to a variety of immune-stimulating agents, which impact nutrient utilization in the pig to support of the immune response. With reductions in antibiotic use, there is an increased need and focus on research into potential nutritional strategies to maintain both animal health and performance when pigs are exposed to immune stimulating conditions. There is robust evidence showing that feeding LP, AA-supplemented diets may overcome the negative effects of undigested protein on gastrointestinal health. Moreover, research has increasingly highlighted AA not only as building blocks for muscle protein, but also as functional agents. Supplementation with key FAA has been shown to enhance pig performance and immune status. Feeding different fiber sources and levels may improve gastrointestinal health by ameliorating intestinal development and renewal, and positively modulating the microbiota. Finally, and particularly for the immediate post-weaning period, increasing diet complexity by incorporating highly digestible (i.e., animal-based) ingredients, and supplementing diets with organic acids and/or plant secondary metabolites may improve the pig’s ability to cope with the weaning transition. Overall, nutritionists will need to incorporate animal health requirements into future nutrition programs and consider multiple nutritional strategies depending on the specific conditions to which pigs are exposed in order to optimize both immune status and productivity of pigs.
Author Contributions
All authors participated in conceptualization, writing, and review and editing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- van der Meer, Y.; Lammers, A.; Jansman, A.J.M.; Rijnen, M.M.J.A.; Hendriks, W.H.; Gerrits, W.J.J. Performance of Pigs Kept under Different Sanitary Conditions Affected by Protein Intake and Amino Acid Supplementation. J. Anim. Sci. 2016, 94, 4704–4719. [Google Scholar] [CrossRef]
- van der Meer, Y.; Jansman, A.J.M.; Gerrits, W.J.J. Low Sanitary Conditions Increase Energy Expenditure for Maintenance and Decrease Incremental Protein Efficiency in Growing Pigs. Animal 2020, 14, 1811–1820. [Google Scholar] [CrossRef]
- Buchanan, J.B.; Johnson, R.W. Regulation of Food Intake by Inflammatory Cytokines in the Brain. Neuroendocrinology 2007, 86, 183–190. [Google Scholar] [CrossRef]
- Johnson, R.W. Immune and Endocrine Regulation of Food Intake in Sick Animals. Domest. Anim. Endocrinol. 1998, 15, 309–319. [Google Scholar] [CrossRef]
- Karrow, N.A. Activation of the Hypothalamic-Pituitary-Adrenal Axis and Autonomic Nervous System during Inflammation and Altered Programming of the Neuroendocrine-Immune Axis during Fetal and Neonatal Development: Lessons Learned from the Model Inflammagen, Lipopolysaccharide. Brain Behav. Immun. 2006, 20, 144–158. [Google Scholar] [CrossRef]
- Meier, U.; Gressner, A.M. Endocrine Regulation of Energy Metabolism: Review of Pathobiochemical and Clinical Chemical Aspects of Leptin, Ghrelin, Adiponectin, and Resistin. Clin. Chem. 2004, 50, 1511–1525. [Google Scholar] [CrossRef]
- Le Floc’h, N.; Melchior, D.; Obled, C. Modifications of Protein and Amino Acid Metabolism during Inflammation and Immune System Activation. Livest. Prod. Sci. 2004, 87, 37–45. [Google Scholar] [CrossRef]
- Pastorelli, H.; van Milgen, J.; Lovatto, P.; Montagne, L. Meta-Analysis of Feed Intake and Growth Responses of Growing Pigs after a Sanitary Challenge. Animal 2012, 6, 952–961. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Ferreira, F.N.A.; Costa, M.O.; Wellington, M.O.; Columbus, D.A. Factors Affecting Performance Response of Pigs Exposed to Different Challenge Models: A Multivariate Approach. J. Anim. Sci. 2021, 99, skab035. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Wellington, M.O.; González-Vega, J.C.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Functional Amino Acid Supplementation, Regardless of Dietary Protein Content, Improves Growth Performance and Immune Status of Weaned Pigs Challenged with Salmonella Typhimurium. J. Anim. Sci. 2021, 99, skaa365. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Wellington, M.O.; González-Vega, J.C.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. A Longer Adaptation Period to a Functional Amino Acid-Supplemented Diet Improves Growth Performance and Immune Status of Salmonella Typhimurium-Challenged Pigs. J. Anim. Sci. 2021, 99, skab146. [Google Scholar] [CrossRef]
- Schweer, W.; Schwartz, K.; Patience, J.F.; Karriker, L.; Sparks, C.; Weaver, M.; Fitzsimmons, M.; Burkey, T.E.; Gabler, N.K. Porcine Reproductive and Respiratory Syndrome Virus Reduces Feed Efficiency, Digestibility, and Lean Tissue Accretion in Grow-Finish Pigs. Trans. Anim. Sci. 2017, 1, 480–488. [Google Scholar] [CrossRef]
- Dee, S.; Guzman, J.E.; Hanson, D.; Garbes, N.; Morrison, R.; Amodie, D.; Galina Pantoja, L. A Randomized Controlled Trial to Evaluate Performance of Pigs Raised in Antibiotic-Free or Conventional Production Systems Following Challenge with Porcine Reproductive and Respiratory Syndrome Virus. PLoS ONE 2018, 13, e0208430. [Google Scholar] [CrossRef]
- Khafipour, E.; Munyaka, P.M.; Nyachoti, C.M.; Krause, D.O.; Rodriguez-Lecompte, J.C. Effect of Crowding Stress and Escherichia coli K88+ Challenge in Nursery Pigs Supplemented with Anti-Escherichia coli K88+ Probiotics. J. Anim. Sci. 2014, 92, 2017–2029. [Google Scholar] [CrossRef]
- Jayaraman, B.; Htoo, J.; Nyachoti, C.M. Effects of Dietary Threonine:Lysine Ratioes and Sanitary Conditions on Performance, Plasma Urea Nitrogen, Plasma-Free Threonine and Lysine of Weaned Pigs. Anim. Nutr. 2015, 1, 283–288. [Google Scholar] [CrossRef]
- Laskoski, F. Effects of Pigs per Feeder Hole and Group Size on Feed Intake Onset, Growth Performance, and Ear and Tail Lesions in Nursery Pigs with Consistent Space Allowance. J. Swine Health Prod. 2019, 27, 12–18. [Google Scholar]
- Rauw, W.M.; de Mercado de la Peña, E.; Gomez-Raya, L.; García Cortés, L.A.; Ciruelos, J.J.; Gómez Izquierdo, E. Impact of Environmental Temperature on Production Traits in Pigs. Sci. Rep. 2020, 10, 2106. [Google Scholar] [CrossRef]
- Wellington, M.O.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Impact of Dietary Fiber and Immune System Stimulation on Threonine Requirement for Protein Deposition in Growing Pigs. J. Anim. Sci. 2018, 96, 5222–5232. [Google Scholar] [CrossRef]
- Dong, G.Z.; Pluske, J.R. The Low Feed Intake in Newly-Weaned Pigs: Problems and Possible Solutions. Asian-Austral. J. Anim. Sci. 2007, 20, 440–452. [Google Scholar] [CrossRef]
- Zheng, L.; Duarte, M.E.; Sevarolli Loftus, A.; Kim, S.W. Intestinal Health of Pigs Upon Weaning: Challenges and Nutritional Intervention. Front. Vet. Sci. 2021, 8, 628258. [Google Scholar] [CrossRef]
- Blecha, F.; Pollman, D.S.; Nichols, D.A. Weaning Pigs at an Early Age Decreases Cellular Immunity. J. Anim. Sci. 1983, 56, 396–400. [Google Scholar] [CrossRef]
- Rooke, J.A.; Bland, I.M. The Acquisition of Passive Immunity in the New-Born Piglet. Livest. Prod. Sci. 2002, 78, 13–23. [Google Scholar] [CrossRef]
- Yuan, Y.; Jansen, J.; Charles, D.; Zanella, A.J. The Influence of Weaning Age on Post-Mixing Agonistic Interactions in Growing Pigs. Appl. Anim. Behav. Sci. 2004, 88, 39–46. [Google Scholar] [CrossRef]
- Spreeuwenberg, M.A.; Verdonk, J.M.; Gaskins, H.R.; Verstegen, M.W. Small Intestine Epithelial Barrier Function Is Compromised in Pigs with Low Feed Intake at Weaning. J. Nutr. 2001, 131, 1520–1527. [Google Scholar] [CrossRef]
- Pluske, J.R.; Kim, J.C.; Black, J.L.; Pluske, J.R.; Kim, J.C.; Black, J.L. Manipulating the Immune System for Pigs to Optimise Performance. Anim. Prod. Sci. 2018, 58, 666–680. [Google Scholar] [CrossRef]
- Marquardt, R.R.; Li, S. Antimicrobial Resistance in Livestock: Advances and Alternatives to Antibiotics. Anim. Front. 2018, 8, 30–37. [Google Scholar] [CrossRef]
- Parra, M.D.; Fuentes, P.; Tecles, F.; Martínez-Subiela, S.; Martínez, J.S.; Muñoz, A.; Cerón, J.J. Porcine Acute Phase Protein Concentrations in Different Diseases in Field Conditions. J. Vet. Med. B 2006, 53, 488–493. [Google Scholar] [CrossRef]
- Reeds, P.J.; Jahoor, F. The Amino Acid Requirements of Disease. Clin. Nutr. 2001, 20, 15–22. [Google Scholar] [CrossRef]
- Obled, C. Amino Acid Requirements in Inflammatory States. Can. J. Anim. Sci. 2003, 83, 365–373. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef]
- Rakhshandeh, A.; de Lange, C.F.M.; Htoo, J.K.; Gheisari, A.; Rakhshandeh, A.R. Immune System Stimulation Increases the Plasma Cysteine Flux and Whole-Body Glutathione Synthesis Rate in Starter Pigs. J. Anim. Sci. 2019, 97, 3871–3881. [Google Scholar] [CrossRef]
- Le Floc’h, N.; Wessels, A.; Corrent, E.; Wu, G.; Bosi, P. The Relevance of Functional Amino Acids to Support the Health of Growing Pigs. Anim. Feed Sci. Technol. 2018, 245, 104–116. [Google Scholar] [CrossRef]
- Luppi, A. Swine Enteric Colibacillosis: Diagnosis, Therapy and Antimicrobial Resistance. Porc. Health Manag. 2017, 3, 16. [Google Scholar] [CrossRef]
- Luzzi, I.; Galetta, P.; Massari, M.; Rizzo, C.; Dionisi, A.M.; Filetici, E.; Cawthorne, A.; Tozzi, A.; Argentieri, M.; Bilei, S.; et al. An Easter Outbreak of Salmonella Typhimurium DT 104A Associated with Traditional Pork Salami in Italy. Eurosurveillance 2007, 12, 11–12. [Google Scholar] [CrossRef]
- Kuhn, K.G.; Sørensen, G.; Torpdahl, M.; Kjeldsen, M.K.; Jensen, T.; Gubbels, S.; Bjerager, G.O.; Wingstrand, A.; Porsbo, L.J.; Ethelberg, S. A Long-Lasting Outbreak of Salmonella Typhimurium U323 Associated with Several Pork Products, Denmark, 2010. Epidemiol. Infect. 2013, 141, 260–268. [Google Scholar] [CrossRef]
- Clements, A.; Young, J.C.; Constantinou, N.; Frankel, G. Infection Strategies of Enteric Pathogenic Escherichia coli. Gut Microbes 2012, 3, 71–87. [Google Scholar] [CrossRef]
- Fàbrega, A.; Vila, J. Salmonella enterica Serovar Typhimurium Skills To Succeed in the Host: Virulence and Regulation. Clin. Microb. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef]
- Das, S.; Jayaratne, R.; Barrett, K.E. The Role of Ion Transporters in the Pathophysiology of Infectious Diarrhea. Cell Mol. Gastroenterol. Hepatol. 2018, 6, 33–45. [Google Scholar] [CrossRef]
- Vereecke, L.; Beyaert, R.; van Loo, G. Enterocyte Death and Intestinal Barrier Maintenance in Homeostasis and Disease. Trends Mol. Med. 2011, 17, 584–593. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Intestinal Redox Biology and Oxidative Stress. Semin. Cell Dev. Biol. 2012, 23, 729–737. [Google Scholar] [CrossRef]
- Parker, A.; Vaux, L.; Patterson, A.M.; Modasia, A.; Muraro, D.; Fletcher, A.G.; Byrne, H.M.; Maini, P.K.; Watson, A.J.M.; Pin, C. Elevated Apoptosis Impairs Epithelial Cell Turnover and Shortens Villi in TNF-Driven Intestinal Inflammation. Cell Death Dis. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Boivin, M.; Ma, T. Mechanism of Cytokine Modulation of Epithelial Tight Junction Barrier. Front. Biosci 2009, 14, 2765–2778. [Google Scholar] [CrossRef]
- Pan, L.; Zhao, P.F.; Ma, X.K.; Shang, Q.H.; Xu, Y.T.; Long, S.F.; Wu, Y.; Yuan, F.M.; Piao, X.S. Probiotic Supplementation Protects Weaned Pigs against Enterotoxigenic Escherichia coli K88 Challenge and Improves Performance Similar to Antibiotics. J. Anim. Sci. 2017, 95, 2627–2639. [Google Scholar] [CrossRef]
- Choi, J.; Wang, L.; Liu, S.; Lu, P.; Zhao, X.; Liu, H.; Lahaye, L.; Santin, E.; Liu, S.; Nyachoti, M.; et al. Effects of a Microencapsulated Formula of Organic Acids and Essential Oils on Nutrient Absorption, Immunity, Gut Barrier Function, and Abundance of Enterotoxigenic Escherichia coli F4 in Weaned Piglets Challenged with E. coli F4. J. Anim. Sci. 2020, 98, skaa259. [Google Scholar] [CrossRef]
- Koo, B.; Bustamante-García, D.; Kim, J.W.; Nyachoti, C.M. Health-Promoting Effects of Lactobacillus-Fermented Barley in Weaned Pigs Challenged with Escherichia coli K88+. Animal 2020, 14, 39–49. [Google Scholar] [CrossRef]
- Heo, J.-M. Reducing the Protein Content in Diets for Weaner Pigs to Control Post-Weaning Diarrhoea: Physiological and Metabolic Responses of the Gastrointestinal Tract. Ph.D. Thesis, Murdoch University, Perth, Australia, 2010. [Google Scholar]
- Ren, M.; Liu, X.T.; Wang, X.; Zhang, G.J.; Qiao, S.Y.; Zeng, X.F. Increased Levels of Standardized Ileal Digestible Threonine Attenuate Intestinal Damage and Immune Responses in Escherichia coli K88+ Challenged Weaned Piglets. Anim. Feed Sci. Technol. 2014, 195, 67–75. [Google Scholar] [CrossRef]
- Capozzalo, M.M.; Resink, J.W.; Htoo, J.K.; Kim, J.C.; de Lange, F.M.; Mullan, B.P.; Hansen, C.F.; Pluske, J.R. Determination of the Optimum Standardised Ileal Digestible Sulphur Amino Acids to Lysine Ratio in Weaned Pigs Challenged with Enterotoxigenic Escherichia coli. Anim. Feed Sci. Technol. 2017, 227, 118–130. [Google Scholar] [CrossRef]
- Wellington, M.O.; Agyekum, A.K.; Hamonic, K.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Effect of Supplemental Threonine above Requirement on Growth Performance of Salmonella Typhimurium Challenged Pigs Fed High-Fiber Diets. J. Anim. Sci. 2019, 97, 3636–3647. [Google Scholar] [CrossRef]
- Koo, B.; Choi, J.; Yang, C.; Nyachoti, C.M. Diet Complexity and L-Threonine Supplementation: Effects on Growth Performance, Immune Response, Intestinal Barrier Function, and Microbial Metabolites in Nursery Pigs. J. Anim. Sci. 2020, 98, skaa125. [Google Scholar] [CrossRef] [PubMed]
- Enøe, C.; Mousing, J.; Schirmer, A.L.; Willeberg, P. Infectious and Rearing-System Related Risk Factors for Chronic Pleuritis in Slaughter Pigs. Prev. Vet. Med. 2002, 54, 337–349. [Google Scholar] [CrossRef]
- Fraile, L.; Alegre, A.; López-Jiménez, R.; Nofrarías, M.; Segalés, J. Risk Factors Associated with Pleuritis and Cranio-Ventral Pulmonary Consolidation in Slaughter-Aged Pigs. Vet. J. 2010, 184, 326–333. [Google Scholar] [CrossRef]
- Meyns, T.; Van Steelant, J.; Rolly, E.; Dewulf, J.; Haesebrouck, F.; Maes, D. A Cross-Sectional Study of Risk Factors Associated with Pulmonary Lesions in Pigs at Slaughter. Vet. J. 2011, 187, 388–392. [Google Scholar] [CrossRef]
- Blanchard, B.; Vena, M.M.; Cavalier, A.; Lannic, J.L.; Gouranton, J.; Kobisch, M. Electron Microscopic Observation of the Respiratory Tract of SPF Piglets Inoculated with Mycoplasma hyopneumoniae. Vet. Microbiol. 1992, 30, 329–341. [Google Scholar] [CrossRef]
- Ferraz, M.E.S.; Almeida, H.M.S.; Storino, G.Y.; Sonálio, K.; Souza, M.R.; Moura, C.A.A.; Costa, W.M.T.; Lunardi, L.; Linhares, D.C.L.; de Oliveira, L.G. Lung Consolidation Caused by Mycoplasma hyopneumoniae Has a Negative Effect on Productive Performance and Economic Revenue in Finishing Pigs. Prev. Vet. Med. 2020, 182, 105091. [Google Scholar] [CrossRef]
- Gauger, P.C.; Lager, K.M.; Vincent, A.L.; Opriessnig, T.; Kehrli, M.E.; Cheung, A.K. Postweaning Multisystemic Wasting Syndrome Produced in Gnotobiotic Pigs Following Exposure to Various Amounts of Porcine Circovirus Type 2a or Type 2b. Vet. Microbiol. 2011, 153, 229–239. [Google Scholar] [CrossRef]
- Helm, E.T.; Curry, S.M.; De Mille, C.M.; Schweer, W.P.; Burrough, E.R.; Zuber, E.A.; Lonergan, S.M.; Gabler, N.K. Impact of Porcine Reproductive and Respiratory Syndrome Virus on Muscle Metabolism of Growing Pigs. J. Anim. Sci. 2019, 97, 3213–3227. [Google Scholar] [CrossRef]
- Cho, H.M.; Kim, E.; Wickramasuriya, S.S.; Shin, T.K.; Heo, J.M. Growth and Gut Performance of Young Pigs in Response to Different Dietary Cellulose Concentration and Rearing Condition. Anim. Biosci. 2021, 34, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.M.; Macelline, S.P.; Wickramasuriya, S.S.; Shin, T.K.; Kim, E.; Son, H.C.; Heo, J.M. Moderate Dietary Boron Supplementation Improved Growth Performance, Crude Protein Digestibility and Diarrhea Index in Weaner Pigs Regardless of the Sanitary Condition. Anim. Biosci. 2022, 35, 434–443. [Google Scholar] [CrossRef]
- Le Floc’h, N.; LeBellego, L.; Matte, J.J.; Melchior, D.; Sève, B. The Effect of Sanitary Status Degradation and Dietary Tryptophan Content on Growth Rate and Tryptophan Metabolism in Weaning Pigs. J. Anim. Sci. 2009, 87, 1686–1694. [Google Scholar] [CrossRef]
- Montagne, L.; Le Floc’h, N.; Arturo-Schaan, M.; Foret, R.; Urdaci, M.C.; Le Gall, M. Comparative Effects of Level of Dietary Fiber and Sanitary Conditions on the Growth and Health of Weanling Pigs. J. Anim. Sci. 2012, 90, 2556–2569. [Google Scholar] [CrossRef] [PubMed]
- te Pas, M.F.W.; Jansman, A.J.M.; Kruijt, L.; van der Meer, Y.; Vervoort, J.J.M.; Schokker, D. Sanitary Conditions Affect the Colonic Microbiome and the Colonic and Systemic Metabolome of Female Pigs. Front. Vet. Sci. 2020, 7, 585730. [Google Scholar] [CrossRef] [PubMed]
- Waititu, S.M.; Yin, F.; Patterson, R.; Yitbarek, A.; Rodriguez-Lecompte, J.C.; Nyachoti, C.M. Dietary Supplementation with a Nucleotide-Rich Yeast Extract Modulates Gut Immune Response and Microflora in Weaned Pigs in Response to a Sanitary Challenge. Animal 2017, 11, 2156–2164. [Google Scholar] [CrossRef]
- Zhao, J.; Harper, A.F.; Estienne, M.J.; Webb, K.E., Jr.; McElroy, A.P.; Denbow, D.M. Growth Performance and Intestinal Morphology Responses in Early Weaned Pigs to Supplementation of Antibiotic-Free Diets with an Organic Copper Complex and Spray-Dried Plasma Protein in Sanitary and Nonsanitary Environments. J. Anim. Sci. 2007, 85, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- de Ridder, K.; Levesque, C.L.; Htoo, J.K.; de Lange, C.F.M. Immune System Stimulation Reduces the Efficiency of Tryptophan Utilization for Body Protein Deposition in Growing Pigs. J. Anim. Sci. 2012, 90, 3485–3491. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.W.; Mellencamp, M.A.; Carroll, J.A.; Boyd, R.D.; Allee, G.L. Acute Feed Intake and Acute-Phase Protein Responses Following a Lipopolysaccharide Challenge in Pigs from Two Dam Lines. Vet. Immunol. Immunopath. 2005, 107, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.Y.; Sun, L.H.; Lin, Y.C.; Ma, X.Y.; Zheng, C.T.; Zhou, G.L.; Chen, F.; Zou, S.T. Effects of Dietary Glycyl-Glutamine on Growth Performance, Small Intestinal Integrity, and Immune Responses of Weaning Piglets Challenged with Lipopolysaccharide. J. Anim. Sci. 2009, 87, 4050–4056. [Google Scholar] [CrossRef]
- Johnson, R.W.; von Borell, E. Lipopolysaccharide-Induced Sickness Behavior in Pigs Is Inhibited by Pretreatment with Indomethacin. J. Anim. Sci. 1994, 72, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Litvak, N.; Rakhshandeh, A.; Htoo, J.K.; de Lange, C.F.M. Immune System Stimulation Increases the Optimal Dietary Methionine to Methionine plus Cysteine Ratio in Growing Pigs. J. Anim. Sci. 2013, 91, 4188–4196. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.; Conn, C.A.; Kluger, M.J.; Zeisberger, E. Kinetics of Systemic and Intrahypothalamic IL-6 and Tumor Necrosis Factor during Endotoxin Fever in Guinea Pigs. Am. J. Physiol.-Reg. I 1993, 265, R653–R658. [Google Scholar] [CrossRef]
- Wu, C.; Xu, Q.; Wang, R.; Qin, L.; Peng, X.; Hu, L.; Liu, Y.; Fang, Z.; Lin, Y.; Xu, S.; et al. Effects of Dietary β-Glucan Supplementation on Growth Performance and Immunological and Metabolic Parameters of Weaned Pigs Administered with Escherichia coli Lipopolysaccharide. Food Funct. 2018, 9, 3338–3343. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Lahaye, L.; He, Z.; Zhang, J.; Yang, C.; Piao, X. Micro-Encapsulated Essential Oils and Organic Acids Combination Improves Intestinal Barrier Function, Inflammatory Responses and Microbiota of Weaned Piglets Challenged with Enterotoxigenic Escherichia coli F4 (K88+). Anim. Nutr. 2020, 6, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Xie, L.; Liu, B.; Zhou, L.; Hu, Y.; Ajuwon, K.M.; Fang, R. Dietary Supplementation of EGF Ameliorates the Negatively Effects of LPS on Early-Weaning Piglets: From Views of Growth Performance, Nutrient Digestibility, Microelement Absorption and Possible Mechanisms. Animals 2021, 11, 1598. [Google Scholar] [CrossRef] [PubMed]
- Brockmeier, S.L.; Halbur, P.G.; Thacker, E.L. Porcine Respiratory Disease Complex. In Polymicrobial Diseases; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2002; pp. 231–258. ISBN 978-1-68367-232-6. [Google Scholar]
- Thacker, E.L. Immunology of the Porcine Respiratory Disease Complex. Vet. Clin. N. Am.-Food A 2001, 17, 551–565. [Google Scholar] [CrossRef]
- Hansen, M.S.; Pors, S.E.; Jensen, H.E.; Bille-Hansen, V.; Bisgaard, M.; Flachs, E.M.; Nielsen, O.L. An Investigation of the Pathology and Pathogens Associated with Porcine Respiratory Disease Complex in Denmark. J. Comp. Pathol. 2010, 143, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Kyriazakis, I.; Emmans, G.C. The Growth of Mammals Following a Period of Nutritional Limitation. J. Theor. Biol. 1992, 156, 485–498. [Google Scholar] [CrossRef]
- Smith, B.N.; Oelschlager, M.L.; Abdul Rasheed, M.S.; Dilger, R.N. Dietary Soy Isoflavones Reduce Pathogen-Related Mortality in Growing Pigs under Porcine Reproductive and Respiratory Syndrome Viral Challenge. J. Anim. Sci. 2020, 98, skaa024. [Google Scholar] [CrossRef]
- Schweer, W.P.; Schwartz, K.; Burrough, E.R.; Yoon, K.J.; Sparks, J.C.; Gabler, N.K. The Effect of Porcine Reproductive and Respiratory Syndrome Virus and Porcine Epidemic Diarrhea Virus Challenge on Growing Pigs: Growth Performance and Digestibility1. J. Anim. Sci. 2016, 94, 514–522. [Google Scholar] [CrossRef]
- Kahindi, R.; Regassa, A.; Htoo, J.; Nyachoti, M. Optimal Sulfur Amino Acid to Lysine Ratio for Post Weaning Piglets Reared under Clean or Unclean Sanitary Conditions. Anim. Nutr. 2017, 3, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, B.; Htoo, J.K.; Nyachoti, C.M. Effects of Different Dietary Tryptophan: Lysine Ratios and Sanitary Conditions on Growth Performance, Plasma Urea Nitrogen, Serum Haptoglobin and Ileal Histomorphology of Weaned Pigs. Anim. Sci. J. 2017, 88, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.M.; González-Ortiz, G.; Melo-Durán, D.; Heo, J.M.; Cordero, G.; Bedford, M.R.; Kim, J.C. Stimbiotic Supplementation Improved Performance and Reduced Inflammatory Response via Stimulating Fiber Fermenting Microbiome in Weaner Pigs Housed in a Poor Sanitary Environment and Fed an Antibiotic-Free Low Zinc Oxide Diet. PLoS ONE 2020, 15, e0240264. [Google Scholar] [CrossRef] [PubMed]
- Bannerman, D.D.; Goldblum, S.E. Mechanisms of Bacterial Lipopolysaccharide-Induced Endothelial Apoptosis. Am. J. PHysiol.-Lung C 2003, 284, L899–L914. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.; Zhang, L.; Hou, Y.; Ding, B.; Yi, D.; Wang, L.; Zhu, H.; Liu, Y.; Yin, Y.; Wu, G. Effects of L-Proline on the Growth Performance, and Blood Parameters in Weaned Lipopolysaccharide (LPS)-Challenged Pigs. Asian-Australas. J. Anim. Sci. 2014, 27, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Opal, S.M.; Scannon, P.J.; Vincent, J.-L.; White, M.; Carroll, S.F.; Palardy, J.E.; Parejo, N.A.; Pribble, J.P.; Lemke, J.H. Relationship between Plasma Levels of Lipopolysaccharide (LPS) and LPS-Binding Protein in Patients with Severe Sepsis and Septic Shock. J. Infect. Dis. 1999, 180, 1584–1589. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, A.; Watson, J.J.; Kammerer, R.A.; Tasab, M.; Progias, P.; Reeves, K.; Brown, N.J.; Brookes, Z.L. Angiopoietin-1 Variant Reduces LPS-Induced Microvascular Dysfunction in a Murine Model of Sepsis. Crit. Care 2012, 16, R182. [Google Scholar] [CrossRef] [PubMed]
- LeMay, L.G.; Vander, A.J.; Kluger, M.J. Role of Interleukin 6 in Fever in Rats. Am. J. Physiol.-Reg. I 1990, 258, R798–R803. [Google Scholar] [CrossRef] [PubMed]
- Bikker, P.; Dirkzwager, A.; Fledderus, J.; Trevisi, P.; le Huërou-Luron, I.; Lallès, J.P.; Awati, A. The Effect of Dietary Protein and Fermentable Carbohydrates Levels on Growth Performance and Intestinal Characteristics in Newly Weaned Piglets. J. Anim. Sci. 2006, 84, 3337–3345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wielen, N.v.d.; Hee, B.v.d.; Wang, J.; Hendriks, W.; Gilbert, M. Impact of Fermentable Protein, by Feeding High Protein Diets, on Microbial Composition, Microbial Catabolic Activity, Gut Health and beyond in Pigs. Microorganisms 2020, 8, 1735. [Google Scholar] [CrossRef] [PubMed]
- Prohászka, L.; Baron, F. The Predisposing Role of High Dietary Protein Supplies in Enteropathogenic E. coli Infections of Weaned Pigs. Zent. Vet. Riehe B 1980, 27, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Nyachoti, C.M.; Omogbenigun, F.O.; Rademacher, M.; Blank, G. Performance Responses and Indicators of Gastrointestinal Health in Early-Weaned Pigs Fed Low-Protein Amino Acid-Supplemented Diets. J. Anim. Sci. 2006, 84, 125–134. [Google Scholar] [CrossRef]
- Pieper, R.; Kröger, S.; Richter, J.F.; Wang, J.; Martin, L.; Bindelle, J.; Htoo, J.K.; von Smolinski, D.; Vahjen, W.; Zentek, J.; et al. Fermentable Fiber Ameliorates Fermentable Protein-Induced Changes in Microbial Ecology, but Not the Mucosal Response, in the Colon of Piglets. J. Nutr. 2012, 142, 661–667. [Google Scholar] [CrossRef]
- Yao, C.K.; Muir, J.G.; Gibson, P.R. Review Article: Insights into Colonic Protein Fermentation, Its Modulation and Potential Health Implications. Aliment. Pharm. Ther. 2016, 43, 181–196. [Google Scholar] [CrossRef]
- Wellock, I.J.; Fortomaris, P.D.; Houdijk, J.G.M.; Kyriazakis, I. Effects of Dietary Protein Supply, Weaning Age and Experimental Enterotoxigenic Escherichia coli Infection on Newly Weaned Pigs: Health. Animal 2008, 2, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Rist, V.T.S.; Weiss, E.; Eklund, M.; Mosenthin, R. Impact of Dietary Protein on Microbiota Composition and Activity in the Gastrointestinal Tract of Piglets in Relation to Gut Health: A Review. Animal 2013, 7, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.M.; Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Pluske, J.R. Feeding a Diet with Decreased Protein Content Reduces Indices of Protein Fermentation and the Incidence of Postweaning Diarrhea in Weaned Pigs Challenged with an Enterotoxigenic Strain of Escherichia coli. J. Anim. Sci. 2009, 87, 2833–2843. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.M.; Kim, J.C.; Yoo, J.; Pluske, J.R. A Between-Experiment Analysis of Relationships Linking Dietary Protein Intake and Post-Weaning Diarrhea in Weanling Pigs under Conditions of Experimental Infection with an Enterotoxigenic Strain of Escherichia coli. Anim. Sci. J. 2015, 86, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.; Hutchings, M.R.; Hutchings, K.E.K.; Gally, D.L.; Houdijk, J.G.M. Changes in the Ileal, but Not Fecal, Microbiome in Response to Increased Dietary Protein Level and Enterotoxigenic Escherichia coli Exposure in Pigs. Appl. Environ. Microb. 2019, 85, e01252-19. [Google Scholar] [CrossRef]
- Gilbert, M.S.; Ijssennagger, N.; Kies, A.K.; van Mil, S.W.C. Protein Fermentation in the Gut; Implications for Intestinal Dysfunction in Humans, Pigs, and Poultry. Am. J. Physiol.-Gastr. L 2018, 315, G159–G170. [Google Scholar] [CrossRef] [PubMed]
- Lallès, J.-P.; Montoya, C.A. Dietary Alternatives to In-Feed Antibiotics, Gut Barrier Function and Inflammation in Piglets Post-Weaning: Where Are We Now? Anim. Feed Sci. Technol. 2021, 274, 114836. [Google Scholar] [CrossRef]
- Yu, D.; Zhu, W.; Hang, S. Effects of Low-Protein Diet on the Intestinal Morphology, Digestive Enzyme Activity, Blood Urea Nitrogen, and Gut Microbiota and Metabolites in Weaned Pigs. Arch. Anim. Nutr. 2019, 73, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, P.; Fan, P.; He, T.; Jacobs, D.; Levesque, C.L.; Johnston, L.J.; Ji, L.; Ma, N.; Chen, Y.; et al. Moderate Dietary Protein Restriction Optimized Gut Microbiota and Mucosal Barrier in Growing Pig Model. Front. Cell. Infect. Microbiol. 2018, 8, 246. [Google Scholar] [CrossRef]
- Spring, S.; Premathilake, H.; Bradway, C.; Shili, C.; DeSilva, U.; Carter, S.; Pezeshki, A. Effect of Very Low-Protein Diets Supplemented with Branched-Chain Amino Acids on Energy Balance, Plasma Metabolomics and Fecal Microbiome of Pigs. Sci. Rep. 2020, 10, 15859. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qiao, S.; Ren, M.; Zeng, X.; Ma, X.; Wu, Z.; Thacker, P.; Wu, G. Supplementation with Branched-Chain Amino Acids to a Low-Protein Diet Regulates Intestinal Expression of Amino Acid and Peptide Transporters in Weanling Pigs. Amino Acids 2013, 45, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, S.; Ashworth, C.J. The Role of Dietary Fibre in Pig Production, with a Particular Emphasis on Reproduction. J. Anim. Sci. Biotechnol. 2018, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Diether, N.E.; Willing, B.P. Microbial Fermentation of Dietary Protein: An Important Factor in Diet–Microbe–Host Interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef]
- Williams, B.A.; Mikkelsen, D.; Flanagan, B.M.; Gidley, M.J. “Dietary Fibre”: Moving beyond the “Soluble/Insoluble” Classification for Monogastric Nutrition, with an Emphasis on Humans and Pigs. J. Anim. Sci. Biotechnol. 2019, 10, 45. [Google Scholar] [CrossRef]
- Htoo, J.K.; Araiza, B.A.; Sauer, W.C.; Rademacher, M.; Zhang, Y.; Cervantes, M.; Zijlstra, R.T. Effect of Dietary Protein Content on Ileal Amino Acid Digestibility, Growth Performance, and Formation of Microbial Metabolites in Ileal and Cecal Digesta of Early-Weaned Pigs1,2. J. Anim. Sci. 2007, 85, 3303–3312. [Google Scholar] [CrossRef]
- Jeaurond, E.A.; Rademacher, M.; Pluske, J.R.; Zhu, C.H.; de Lange, C.F.M. Impact of Feeding Fermentable Proteins and Carbohydrates on Growth Performance, Gut Health and Gastrointestinal Function of Newly Weaned Pigs. Can. J. Anim. Sci. 2008, 88, 271–281. [Google Scholar] [CrossRef]
- Molist, F.; van Oostrum, M.; Pérez, J.F.; Mateos, G.G.; Nyachoti, C.M.; van der Aar, P.J. Relevance of Functional Properties of Dietary Fibre in Diets for Weanling Pigs. Anim. Feed Sci. Technol. 2014, 189, 1–10. [Google Scholar] [CrossRef]
- Jha, R.; Berrocoso, J.D. Review: Dietary Fiber Utilization and Its Effects on Physiological Functions and Gut Health of Swine. Animal 2015, 9, 1441–1452. [Google Scholar] [CrossRef]
- Che, L.; Chen, H.; Yu, B.; He, J.; Zheng, P.; Mao, X.; Yu, J.; Huang, Z.; Chen, D. Long-Term Intake of Pea Fiber Affects Colonic Barrier Function, Bacterial and Transcriptional Profile in Pig Model. Nutr. Cancer 2014, 66, 388–399. [Google Scholar] [CrossRef]
- Li, Q.; Peng, X.; Burrough, E.R.; Sahin, O.; Gould, S.A.; Gabler, N.K.; Loving, C.L.; Dorman, K.S.; Patience, J.F. Dietary Soluble and Insoluble Fiber With or Without Enzymes Altered the Intestinal Microbiota in Weaned Pigs Challenged With Enterotoxigenic E. coli F18. Front. Microbiol. 2020, 11, 1110. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Burrough, E.R.; Gabler, N.K.; Loving, C.L.; Sahin, O.; Gould, S.A.; Patience, J.F. A Soluble and Highly Fermentable Dietary Fiber with Carbohydrases Improved Gut Barrier Integrity Markers and Growth Performance in F18 ETEC Challenged Pigs. J. Anim. Sci. 2019, 97, 2139–2153. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, L.L.; Jakobsen, M.; Jensen, B.B. Effects of Dietary Oligosaccharides on Microbial Diversity and Fructo-Oligosaccharide Degrading Bacteria in Faeces of Piglets Post-Weaning. Anim. Feed Sci. Technol. 2003, 109, 133–150. [Google Scholar] [CrossRef]
- Konstantinov, S.R.; Awati, A.; Smidt, H.; Williams, B.A.; Akkermans, A.D.L.; de Vos, W.M. Specific Response of a Novel and Abundant Lactobacillus amylovorus-Like Phylotype to Dietary Prebiotics in the Guts of Weaning Piglets. Appl. Environ. Microb. 2004, 70, 3821–3830. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Piao, X.S.; Kim, S.W.; Wang, L.; Shen, Y.B.; Lee, H.S.; Li, S.Y. Effects of Chito-Oligosaccharide Supplementation on the Growth Performance, Nutrient Digestibility, Intestinal Morphology, and Fecal Shedding of Escherichia coli and Lactobacillus in Weaning Pigs. J. Anim. Sci. 2008, 86, 2609–2618. [Google Scholar] [CrossRef]
- Molist, F.; Gómez de Segura, A.; Pérez, J.F.; Bhandari, S.K.; Krause, D.O.; Nyachoti, C.M. Effect of Wheat Bran on the Health and Performance of Weaned Pigs Challenged with Escherichia coli K88+. Livest. Sci. 2010, 133, 214–217. [Google Scholar] [CrossRef]
- Bach Knudsen, K.E.; Hedemann, M.S.; Lærke, H.N. The Role of Carbohydrates in Intestinal Health of Pigs. Anim. Feed Sci. Technol. 2012, 173, 41–53. [Google Scholar] [CrossRef]
- Taciak, M.; Barszcz, M.; Święch, E.; Tuśnio, A.; Bachanek, I. Interactive Effects of Protein and Carbohydrates on Production of Microbial Metabolites in the Large Intestine of Growing Pigs. Arch. Anim. Nutr. 2017, 71, 192–209. [Google Scholar] [CrossRef]
- Mariscal-Landín, G.; Sève, B.; Colléaux, Y.; Lebreton, Y. Endogenous Amino Nitrogen Collected from Pigs with End-to-End Ileorectal Anastomosis Is Affected by the Method of Estimation and Altered by Dietary Fiber. J. Nutr. 1995, 125, 136–146. [Google Scholar] [CrossRef]
- Schmidt-Wittig, U.; Enss, M.-L.; Coenen, M.; Gärtner, K.; Hedrich, H.J. Response of Rat Colonic Mucosa to a High Fiber Diet. Ann. Nutr. Metab. 1996, 40, 343–350. [Google Scholar] [CrossRef]
- Hedemann, M.S.; Eskildsen, M.; Lærke, H.N.; Pedersen, C.; Lindberg, J.E.; Laurinen, P.; Knudsen, K.E.B. Intestinal Morphology and Enzymatic Activity in Newly Weaned Pigs Fed Contrasting Fiber Concentrations and Fiber Properties. J. Anim. Sci. 2006, 84, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Wellington, M.O.; Hamonic, K.; Krone, J.E.C.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Effect of Dietary Fiber and Threonine Content on Intestinal Barrier Function in Pigs Challenged with Either Systemic E. Coli Lipopolysaccharide or Enteric Salmonella Typhimurium. J. Anim. Sci. Biotechnol. 2020, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R.; Siba, P.M.; Pethick, D.W.; Durmic, Z.; Mullan, B.P.; Hampson, D.J. The Incidence of Swine Dysentery in Pigs Can Be Reduced by Feeding Diets That Limit the Amount of Fermentable Substrate Entering the Large Intestine. J. Nutr. 1996, 126, 2920–2933. [Google Scholar] [CrossRef]
- Dritz, S.S.; Shi, J.; Kielian, T.L.; Goodband, R.D.; Nelssen, J.L.; Tokach, M.D.; Chengappa, M.M.; Smith, J.E.; Blecha, F. Influence of Dietary β-Glucan on Growth Performance, Nonspecific Immunity, and Resistance to Streptococcus suis Infection in Weanling Pigs. J. Anim. Sci. 1995, 73, 3341–3350. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Li, D.F.; Nelssen, J.L.; Reddy, P.G.; Blecha, F.; Hancock, J.D.; Allee, G.L.; Goodband, R.D.; Klemm, R.D. Transient Hypersensitivity to Soybean Meal in the Early-Weaned Pig. J. Anim. Sci. 1990, 68, 1790–1799. [Google Scholar] [CrossRef] [PubMed]
- Stoner, G.R.; Allee, G.L.; Nelssen, J.L.; Johnston, M.E.; Goodband, R.D. Effect of Select Menhaden Fish Meal in Starter Diets for Pigs. J. Anim. Sci. 1990, 68, 2729–2735. [Google Scholar] [CrossRef] [PubMed]
- Peace, R.M.; Campbell, J.; Polo, J.; Crenshaw, J.; Russell, L.; Moeser, A. Spray-Dried Porcine Plasma Influences Intestinal Barrier Function, Inflammation, and Diarrhea in Weaned Pigs. J. Nutr. 2011, 141, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Mahan, D.C.; Fastinger, N.D.; Peters, J.C. Effects of Diet Complexity and Dietary Lactose Levels during Three Starter Phases on Postweaning Pig Performance. J. Anim. Sci. 2004, 82, 2790–2797. [Google Scholar] [CrossRef]
- Dritz, S.S.; Owen, K.Q.; Goodband, R.D.; Nelssen, J.L.; Tokach, M.D.; Chengappa, M.M.; Blecha, F. Influence of Lipopolysaccharide-Induced Immune Challenge and Diet Complexity on Growth Performance and Acute-Phase Protein Production in Segregated Early-Weaned Pigs. J. Anim. Sci. 1996, 74, 1620–1628. [Google Scholar] [CrossRef]
- Wolter, B.F.; Ellis, M.; Corrigan, B.P.; DeDecker, J.M.; Curtis, S.E.; Parr, E.N.; Webel, D.M. Impact of Early Postweaning Growth Rate as Affected by Diet Complexity and Space Allocation on Subsequent Growth Performance of Pigsin a Wean-to-Finish Production System. J. Anim. Sci. 2003, 81, 353–359. [Google Scholar] [CrossRef]
- Koo, B.; Kim, J.W.; de Lange, C.F.M.; Hossain, M.M.; Nyachoti, C.M. Effects of Diet Complexity and Multicarbohydrase Supplementation on Growth Performance, Nutrient Digestibility, Blood Profile, Intestinal Morphology, and Fecal Score in Newly Weaned Pigs. J. Anim. Sci. 2017, 95, 4060–4071. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, J.; Wang, G.; Cai, S.; Zeng, X.; Qiao, S. Advances in Low-Protein Diets for Swine. J. Anim. Sci. Biotechnol. 2018, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.G.; Biden, R.S. The Effect of Protein Nutrition between 5–5 and 20 Kg Live Weight on the Subsequent Performance and Carcass Quality of Pigs. Anim. Sci. 1978, 27, 223–228. [Google Scholar] [CrossRef]
- Skinner, L.D.; Levesque, C.L.; Wey, D.; Rudar, M.; Zhu, J.; Hooda, S.; de Lange, C.F.M. Impact of Nursery Feeding Program on Subsequent Growth Performance, Carcass Quality, Meat Quality, and Physical and Chemical Body Composition of Growing-Finishing Pigs. J. Anim. Sci. 2014, 92, 1044–1054. [Google Scholar] [CrossRef]
- Reeds, P.J.; Fjeld, C.R.; Jahoor, F. Do the Differences between the Amino Acid Compositions of Acute-Phase and Muscle Proteins Have a Bearing on Nitrogen Loss in Traumatic States? J. Nutr. 1994, 124, 906–910. [Google Scholar] [CrossRef]
- Quiniou, N.; Dourmad, J.-Y.; Noblet, J. Effect of Energy Intake on the Performance of Different Types of Pig from 45 to 100 Kg Body Weight. 1. Protein and Lipid Deposition. Anim. Sci. 1996, 63, 277–288. [Google Scholar] [CrossRef]
- Le Naou, T.; Le Floc’h, N.; Louveau, I.; Gilbert, H.; Gondret, F. Metabolic Changes and Tissue Responses to Selection on Residual Feed Intake in Growing Pigs. J. Anim. Sci. 2012, 90, 4771–4780. [Google Scholar] [CrossRef]
- Klasing, K.C. Nutrition and the Immune System. Br. Poult. Sci. 2007, 48, 525–537. [Google Scholar] [CrossRef]
- Jan-Willem Van Klinken, B.; Einerhand, A.W.C.; Büller, H.A.; Dekker, J. Strategic Biochemical Analysis of Mucins. Anal. Biochem. 1998, 265, 103–116. [Google Scholar] [CrossRef]
- Bauchart-Thevret, C.; Stoll, B.; Chacko, S.; Burrin, D.G. Sulfur Amino Acid Deficiency Upregulates Intestinal Methionine Cycle Activity and Suppresses Epithelial Growth in Neonatal Pigs. Am. J. Physiol.-Endoc. M 2009, 296, E1239–E1250. [Google Scholar] [CrossRef]
- Cebra, J.J. Influences of Microbiota on Intestinal Immune System Development. Am. J. Clin Nutr. 1999, 69, 1046s–1051s. [Google Scholar] [CrossRef] [PubMed]
- Law, G.K.; Bertolo, R.F.; Adjiri-Awere, A.; Pencharz, P.B.; Ball, R.O. Adequate Oral Threonine Is Critical for Mucin Production and Gut Function in Neonatal Piglets. Am. J. Physiol.-Gastr. L 2007, 292, G1293–G1301. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Functional Amino Acids in Nutrition and Health. Amino Acids 2013, 45, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zong, E.Y.; Huang, P.F.; Yang, H.S.; Yan, S.L.; Li, J.Z.; Li, Y.L.; Ding, X.Q.; He, S.P.; Xiong, X.; et al. The Effects of Dietary Sulfur Amino Acids on Serum Biochemical Variables, Mucosal Amino Acid Profiles, and Intestinal Inflammation in Weaning Piglets. Livest. Sci. 2019, 220, 32–36. [Google Scholar] [CrossRef]
- Yan, S.; Long, L.; Zong, E.; Huang, P.; Li, J.; Li, Y.; Ding, X.; Xiong, X.; Yin, Y.; Yang, H. Dietary Sulfur Amino Acids Affect Jejunal Cell Proliferation and Functions by Affecting Antioxidant Capacity, Wnt/β-Catenin, and the Mechanistic Target of Rapamycin Signaling Pathways in Weaning Piglets. J. Anim. Sci. 2018, 96, 5124–5133. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef]
- Liang, H.; Dai, Z.; Liu, N.; Ji, Y.; Chen, J.; Zhang, Y.; Yang, Y.; Li, J.; Wu, Z.; Wu, G. Dietary L-Tryptophan Modulates the Structural and Functional Composition of the Intestinal Microbiome in Weaned Piglets. Front. Microbiol. 2018, 9, 1736. [Google Scholar] [CrossRef]
- Klasing, K.C. Nutritional Modulation of Resistance to Infectious Diseases. Poult. Sci. 1998, 77, 1119–1125. [Google Scholar] [CrossRef]
- Defa, L.; Changting, X.; Shiyan, Q.; Jinhui, Z.; Johnson, E.W.; Thacker, P.A. Effects of Dietary Threonine on Performance, Plasma Parameters and Immune Function of Growing Pigs. Anim. Feed Sci. Technol. 1999, 78, 179–188. [Google Scholar] [CrossRef]
- Wang, X.; Qiao, S.Y.; Liu, M.; Ma, Y.X. Effects of Graded Levels of True Ileal Digestible Threonine on Performance, Serum Parameters and Immune Function of 10–25kg Pigs. Anim. Feed Sci. Technol. 2006, 129, 264–278. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Panisson, J.C.; Kpogo, L.A.; González-Vega, J.C.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Functional Amino Acid Supplementation Postweaning Mitigates the Response of Normal Birth Weight More than for Low Birth Weight Pigs to a Subsequent Salmonella Challenge. Animal 2022, 16, 100566. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.A.; Panisson, J.C.; Van Kessel, A.G.; Columbus, D.A. Functional Amino Acid Supplementation Attenuates the Negative Effects of Plant-Based Nursery Diets on the Response of Pigs to a Subsequent Salmonella Typhimurium Challenge. J. Anim. Sci. 2022, skac267. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.H.; Seok, W.J.; Kim, I.H. Organic Acids Mixture as a Dietary Additive for Pigs—A Review. Animals 2020, 10, 952. [Google Scholar] [CrossRef] [PubMed]
- Gómez-García, M.; Sol, C.; de Nova, P.J.G.; Puyalto, M.; Mesas, L.; Puente, H.; Mencía-Ares, Ó.; Miranda, R.; Argüello, H.; Rubio, P.; et al. Antimicrobial Activity of a Selection of Organic Acids, Their Salts and Essential Oils against Swine Enteropathogenic Bacteria. Porc. Health Manag. 2019, 5, 32. [Google Scholar] [CrossRef]
- Zentek, J.; Buchheit-Renko, S.; Ferrara, F.; Vahjen, W.; Kessel, A.G.V.; Pieper, R. Nutritional and Physiological Role of Medium-Chain Triglycerides and Medium-Chain Fatty Acids in Piglets. Anim. Health Res. Rev. 2011, 12, 83–93. [Google Scholar] [CrossRef]
- Gebru, E.; Lee, J.S.; Son, J.C.; Yang, S.Y.; Shin, S.A.; Kim, B.; Kim, M.K.; Park, S.C. Effect of Probiotic-, Bacteriophage-, or Organic Acid-Supplemented Feeds or Fermented Soybean Meal on the Growth Performance, Acute-Phase Response, and Bacterial Shedding of Grower Pigs Challenged with Salmonella enterica Serotype Typhimurium. J. Anim. Sci. 2010, 88, 3880–3886. [Google Scholar] [CrossRef] [PubMed]
- Calveyra, J.C.; Nogueira, M.G.; Kich, J.D.; Biesus, L.L.; Vizzotto, R.; Berno, L.; Coldebella, A.; Lopes, L.; Morés, N.; Lima, G.J.M.M.; et al. Effect of Organic Acids and Mannanoligosaccharide on Excretion of Salmonella Typhimurium in Experimentally Infected Growing Pigs. Res. Vet. Sci. 2012, 93, 46–47. [Google Scholar] [CrossRef]
- Martín-Peláez, S.; Costabile, A.; Hoyles, L.; Rastall, R.A.; Gibson, G.R.; La Ragione, R.M.; Woodward, M.J.; Mateu, E.; Martín-Orúe, S.M. Evaluation of the Inclusion of a Mixture of Organic Acids or Lactulose into the Feed of Pigs Experimentally Challenged with Salmonella Typhimurium. Vet. Microbiol. 2010, 142, 337–345. [Google Scholar] [CrossRef]
- Walsh, M.C.; Rostagno, M.H.; Gardiner, G.E.; Sutton, A.L.; Richert, B.T.; Radcliffe, J.S. Controlling Salmonella Infection in Weanling Pigs through Water Delivery of Direct-Fed Microbials or Organic Acids. Part I: Effects on Growth Performance, Microbial Populations, and Immune Status. J. Anim. Sci. 2012, 90, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Fabà, L.; Litjens, R.; Allaart, J.; van den Hil, P.R. Feed Additive Blends Fed to Nursery Pigs Challenged with Salmonella. J. Anim. Sci. 2020, 98, skz382. [Google Scholar] [CrossRef]
- Owusu-Asiedu, A.; Nyachoti, C.M.; Marquardt, R.R. Response of Early-Weaned Pigs to an Enterotoxigenic Escherichia coli (K88) Challenge When Fed Diets Containing Spray-Dried Porcine Plasma or Pea Protein Isolate plus Egg Yolk Antibody, Zinc Oxide, Fumaric Acid, or Antibiotic. J. Anim. Sci. 2003, 81, 1790–1798. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.J.; Park, J.W.; Baek, D.H.; Kim, J.K.; Kim, I.H. Feeding the Blend of Organic Acids and Medium Chain Fatty Acids Reduces the Diarrhea in Piglets Orally Challenged with Enterotoxigenic Escherichia coli K88. Anim. Feed Sci. Technol. 2017, 224, 46–51. [Google Scholar] [CrossRef]
- Ren, C.; Zhou, Q.; Guan, W.; Lin, X.; Wang, Y.; Song, H.; Zhang, Y. Immune Response of Piglets Receiving Mixture of Formic and Propionic Acid Alone or with Either Capric Acid or Bacillus licheniformis after Escherichia coli Challenge. BioMed Res. Int. 2019, 2019, 6416187. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, M.J.; Berrios, R.; Stelzhammer, S.; Hohmann, M.; Verri, W., Jr.; Bracarense, A.P.F.R.L. Ingestion of Organic Acids and Cinnamaldehyde Improves Tissue Homeostasis of Piglets Exposed to Enterotoxic Escherichia coli (ETEC). J. Anim. Sci. 2020, 98, skaa012. [Google Scholar] [CrossRef]
- Han, Y.; Zhan, T.; Zhao, Q.; Tang, C.; Zhang, K.; Han, Y.; Zhang, J. Effects of Mixed Organic Acids and Medium Chain Fatty Acids as Antibiotic Alternatives on the Performance, Serum Immunity, and Intestinal Health of Weaned Piglets Orally Challenged with Escherichia coli K88. Anim. Feed Sci. Technol. 2020, 269, 114617. [Google Scholar] [CrossRef]
- Stensland, I.; Kim, J.C.; Bowring, B.; Collins, A.M.; Mansfield, J.P.; Pluske, J.R. A Comparison of Diets Supplemented with a Feed Additive Containing Organic Acids, Cinnamaldehyde and a Permeabilizing Complex, or Zinc Oxide, on Post-Weaning Diarrhoea, Selected Bacterial Populations, Blood Measures and Performance in Weaned Pigs Experimentally Infected with Enterotoxigenic E. coli. Animals 2015, 5, 1147–1168. [Google Scholar] [CrossRef] [PubMed]
- Jamwal, K.; Bhattacharya, S.; Puri, S. Plant Growth Regulator Mediated Consequences of Secondary Metabolites in Medicinal Plants. J. Appl. Res. Med. Aromat. Plants 2018, 9, 26–38. [Google Scholar] [CrossRef]
- Nath, H.; Samtiya, M.; Dhewa, T. Beneficial Attributes and Adverse Effects of Major Plant-Based Foods Anti-Nutrients on Health: A Review. Hum. Nutr. Metab. 2022, 28, 200147. [Google Scholar] [CrossRef]
- Redondo, L.; Chacana, P.; Dominguez, J.; Fernandez Miyakawa, M. Perspectives in the Use of Tannins as Alternative to Antimicrobial Growth Promoter Factors in Poultry. Front. Microbiol. 2014, 5, 118. [Google Scholar] [CrossRef] [PubMed]
- Isaak, C.K.; Petkau, J.C.; Karmin, O.; Ominski, K.; Rodriguez-Lecompte, J.C.; Siow, Y.L. Seasonal Variations in Phenolic Compounds and Antioxidant Capacity of Cornus Stolonifera Plant Material: Applications in Agriculture. Can. J. Plant Sci. 2013, 93, 725–734. [Google Scholar] [CrossRef]
- Rossi, B.; Toschi, A.; Piva, A.; Grilli, E. Single Components of Botanicals and Nature-Identical Compounds as a Non-Antibiotic Strategy to Ameliorate Health Status and Improve Performance in Poultry and Pigs. Nutr. Res. Rev. 2020, 33, 218–234. [Google Scholar] [CrossRef]
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential Oils as Alternatives to Antibiotics in Swine Production. Anim. Nutr. 2018, 4, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhou, Y.F.; Zou, Y.; Hu, X.M.; Zheng, L.F.; Wei, H.K.; Giannenas, I.; Jin, L.Z.; Peng, J.; Jiang, S.W. Effects of Dietary Oregano Essential Oil Supplementation on the Stress Response, Antioxidative Capacity, and HSPs MRNA Expression of Transported Pigs. Livest. Sci. 2015, 180, 143–149. [Google Scholar] [CrossRef]
- Su, G.; Zhou, X.; Wang, Y.; Chen, D.; Chen, G.; Li, Y.; He, J. Effects of Plant Essential Oil Supplementation on Growth Performance, Immune Function and Antioxidant Activities in Weaned Pigs. Lipids Health Dis. 2018, 17, 139. [Google Scholar] [CrossRef]
- Li, S.Y.; Ru, Y.J.; Liu, M.; Xu, B.; Péron, A.; Shi, X.G. The Effect of Essential Oils on Performance, Immunity and Gut Microbial Population in Weaner Pigs. Livest. Sci. 2012, 145, 119–123. [Google Scholar] [CrossRef]
- Lan, R.X.; Li, T.S.; Kim, I.H. Effects of Essential Oils Supplementation in Different Nutrient Densities on Growth Performance, Nutrient Digestibility, Blood Characteristics and Fecal Microbial Shedding in Weaning Pigs. Anim. Feed Sci. Technol. 2016, 214, 77–85. [Google Scholar] [CrossRef]
- Tian, Q.; Piao, X. Essential Oil Blend Could Decrease Diarrhea Prevalence by Improving Antioxidative Capability for Weaned Pigs. Animals 2019, 9, 847. [Google Scholar] [CrossRef]
- Ruzauskas, M.; Bartkiene, E.; Stankevicius, A.; Bernatoniene, J.; Zadeike, D.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Grigas, J.; Zokaityte, E.; et al. The Influence of Essential Oils on Gut Microbial Profiles in Pigs. Animals 2020, 10, 1734. [Google Scholar] [CrossRef]
- Cairo, P.L.G.; Gois, F.D.; Sbardella, M.; Silveira, H.; de Oliveira, R.M.; Allaman, I.B.; Cantarelli, V.S.; Costa, L.B. Effects of Dietary Supplementation of Red Pepper (Schinus Terebinthifolius Raddi) Essential Oil on Performance, Small Intestinal Morphology and Microbial Counts of Weanling Pigs. J. Sci. Food Agric. 2018, 98, 541–548. [Google Scholar] [CrossRef]
- Oh, H.J.; Kim, I.H.; Song, M.H.; Kwak, W.G.; Yun, W.; Lee, J.H.; Lee, C.H.; Oh, S.Y.; Liu, S.; An, J.S.; et al. Effects of Microencapsulated Complex of Organic Acids and Essential Oils on Growth Performance, Nutrient Retention, Blood Profiles, Fecal Microflora, and Lean Meat Percentage in Weaning to Finishing Pigs. Can. J. Anim. Sci. 2019, 99, 41–49. [Google Scholar] [CrossRef]
- Coddens, A.; Loos, M.; Vanrompay, D.; Remon, J.P.; Cox, E. Cranberry Extract Inhibits in Vitro Adhesion of F4 and F18+ Escherichia coli to Pig Intestinal Epithelium and Reduces in Vivo Excretion of Pigs Orally Challenged with F18+ Verotoxigenic E. coli. Vet. Microbiol. 2017, 202, 64–71. [Google Scholar] [CrossRef]
- Brenes, A.; Viveros, A.; Chamorro, S.; Arija, I. Use of Polyphenol-Rich Grape by-Products in Monogastric Nutrition. A Review. Anim. Feed Sci. Technol. 2016, 211, 1–17. [Google Scholar] [CrossRef]
- Kafantaris, I.; Stagos, D.; Kotsampasi, B.; Hatzis, A.; Kypriotakis, A.; Gerasopoulos, K.; Makri, S.; Goutzourelas, N.; Mitsagga, C.; Giavasis, I. Grape Pomace Improves Performance, Antioxidant Status, Fecal Microbiota and Meat Quality of Piglets. Animal 2018, 12, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Gessner, D.K.; Fiesel, A.; Most, E.; Dinges, J.; Wen, G.; Ringseis, R.; Eder, K. Supplementation of a Grape Seed and Grape Marc Meal Extract Decreases Activities of the Oxidative Stress-Responsive Transcription Factors NF-ΚB and Nrf2 in the Duodenal Mucosa of Pigs. Acta Vet. Scand. 2013, 55, 18. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).