4.1. Dietary PPP Affected Growth Performance and Diarrhea Rate
Growth retardation and diarrhea of weaned piglets are important problems in pig production. CTC is often used in pig production to alleviate piglet diarrhea [
18]. Studies have shown that plant extracts such as flavonoids [
4], pectin [
5], and vitamins can relieve diarrhea in animals; pomelo peel contains bioactive substances such as flavonoids, pectin, and vitamins. The results in this study showed that the diarrhea rate in weaned piglets fed a CON was not relieved at the 3rd and 4th weeks. However, weaned piglets administered PPP and CTC significantly decreased the diarrhea rate, whereas there no significant difference in diarrhea between PPP and CTC, which indicated that PPP had the same effect as CTC in fighting piglet diarrhea. The essential oils in pomelo peel contain terpenes, aldehydes, and alcohol constituents (α-terpineol, α-pinene, α-phellandrene, β-myrcene, β-pinene, β-phellandrene, β-caryophyllene, ρ-cymene, citral, citronellal, linalool, and ocimene) which have antimicrobial activity [
19]. The mechanism of these constituents in pomelo peel to inhibit bacterial growth owes to attacking the phospholipid bilayer of the cellular membrane, disrupting enzyme systems, damaging genetic substances of pathogenic bacteria [
20].
The ADG and ADFI of weaned piglets fed PPP were significantly higher than those in CON, indicating that dietary PPP can effectively improve the loss of appetite in weaned piglets under weaning stress and relieve the symptoms of slow development caused by lack of food. Supplementation of PPP containing 1.56 mg/g flavonoids increased average daily gain of piglets, and decreased diarrhea rate. This result was consistent with previous studies reporting the addition of citrus flavonoids reduced the occurrence of zoonotic diseases and has a significant effect on improving animal performance [
21,
22].
4.2. Dietary PPP Affected Serum Antioxidant Enzyme Activities
Both GSH-Px and SOD can catalyze the reduction in cytotoxic peroxides or superoxide free radicals and protect cells, which is an important index for the evaluation of oxidative stress in the body. CAT catalyzes the decomposition of H
2O
2 and is a marker enzyme of peroxisomes, accounting for approximately 40% of the total peroxisome [
23]. Various antioxidants and antioxidant enzymes in the body constitute T-AOC, and the T-AOC level reflects antioxidant capacity in the measured substances.
In this study, serum CAT and GSH-Px activities of piglets in PPP were significantly higher than those in CON and CTC, whereas there were no significant differences between CON and CTC. Serum SOD activities and T-AOC of piglets in PPP were significantly higher than those in CON, whereas there were no significant differences between CON and CTC, or between PPP and CTC. This result suggests that oxidative damage caused by weaning of piglets can lead to a decrease in T-AOC and the activities of GSH-Px, CAT, and SOD, and the antioxidant capacity of PPP is higher than that of CTC. Here, PPP shows stronger antioxidant capacity because more antioxidants are released in PPP under digestion of piglets. The bioactive substances in PPP, such as naringin, naringin, and volatile oils, can enhance the antioxidant capacity of the body by increasing the activity of antioxidant enzymes [
7,
16]. CTC alleviates stress by inhibiting bacterial invasion, and SOD catalyzes the disproportionation of superoxide anion radicals to produce O
2 and H
2O
2 [
24].
4.3. Dietary PPP Affected Serum Cytokines and Immunoglobulin
IL-1β can promote T-cell activation, costimulatory antigen-presenting cells, B-cell proliferation, and antibody secretion [
25]. IL-8 attracts and activates neutrophils. After binding to IL-8, neutrophils activate displacement and release inflammatory mediators, leading to a local inflammatory response [
26]. IL-17 can induce endothelial cells, epithelial cells, and fibroblasts to secrete PGE2 IL-6 and IL-8 [
27]. IFN-γ has antitumor, immunomodulatory, and antiviral properties and activate APC and promote Th1 cell differentiation by upregulating transcription factors [
28]. In this experiment, we observed that dietary PPP significantly reduced pro-inflammatory cytokines (IL-1β, IL-8, IL-17, and IFN-γ) caused by weaning of piglets. Hu et al. [
16] observed that dietary citrus Peel Powder significantly reduced the increase in pro-inflammatory cytokines (MCP-1, IL-6, and TNF-α) caused by HFD. Liu et al. [
29] also suggested that citrus peels can reduce pro-inflammatory cytokines (IL-6, TNF-α, and MCP-1) to exhibit anti-inflammatory activity, which is consistent with our results. The anti-inflammatory cytokine IL-4 in PPP was significantly higher than in CON, indicating that dietary PPP alleviated the occurrence of inflammatory reactions in the body. IL-4 can promote Th2 cells and exert an anti-inflammatory effect [
30]. The bioactive substances in PPP can participate in anti-inflammatory reactions in the body [
31].
Immunoglobulins are proteins secreted by immune cells. They have immunomodulatory functions and can bind antigens to resist pathogenic infections. In this experiment, PPP can significantly increase serum IgA and IgG content of piglets, indicating that PPP can produce more immunoglobulin in body and enhance immune response, indicating that PPP can improve the immunity of the body. The active components such as flavonoids and volatile oil in pomelo peel have a good effect on improving the immunity of piglets [
32].
IGF-1 lowers blood sugar and blood lipids, relaxes blood vessels, and promotes cell growth and differentiation [
33]. It was found that naringin had a hypolipidemic effect in hyperlipidemic mice; however, in this experiment, the IGF-1 level in PPP was significantly higher than that of CON, indicating that PPP had an effective active substance that promotes the synthesis of IGF-1 in weaned piglets, which may indicate that pomelo peel can also have the same hypolipidemic effect in pigs.
A lot of bioactive compounds in CPP have an important role in the health of the organism. Citrus fruit peel contains essential oils, pectin, and polyphenol [
29]. Limonene and the main component in citrus essential oil have antibacterial and antioxidant activity [
34]. Pectin in citrus fruits is effective in improving intestinal inflammation [
29]. The active components, such as flavonoids and volatile oil, in pomelo peel have a good effect on improving the immunity of piglets and reducing the inflammatory reaction, thus reducing the diarrhea rate of piglets.
4.4. Dietary PPP Affected Intestinal Morphology and Intestinal pH of Weaned Piglets
This study showed that ileum villus height and V/C ratio in CTC and PPP were significantly higher than those in CON, which may indicate that PPP and CTC can increase the development of the ileum tissue of weaned piglets. This may be due to the fact that PPP and CTC can maintain an intestinal epithelial cell barrier and alleviate intestinal inflammation caused by weaning of piglets and PPP contains pectin which protects the integrity of intestinal mucosa. The height of the ileum villi reflects the absorption ability of the small intestine, and the decrease in villus height indicates that the absorption ability is low, and the depth of the crypt reflects the quality of the secretory function of the small intestine. A decrease in the V/C ratio indicates the low digestion and absorption ability of the ileum with diarrhea [
35].
The results showed that PPP and CTC can significantly reduce the pH value of the duodenum and cecum content in piglets, indicating that PPP and antibiotics can reduce the normal function of intestinal pH, maintain the stability of intestinal microorganisms in piglets, promote the absorption and utilization of nutrients in piglets, and promote the growth and development of piglets. pH is one of the most important factors for maintaining homeostasis in the internal environment of animals. Before weaning, breast milk was obtained from all piglets. After weaning, piglets were transferred directly from liquid breast milk to solid feed, feed intake decreased, lactic acid produced by lactose fermentation was limited, and insufficient gastric acid secretion led to an increase in gastrointestinal pH. Lactic acid produced by lactose fermentation is an important substance for maintaining low intestinal acidity [
36]. The secretion and activity of pepsin, trypsin, chymotrypsin, and amylase are affected by intestinal pH [
37]. The decrease in pH in the duodenum and cecum of weaned piglets in the CTC and PPP was attributable to the inhibition of pathogenic bacteria, and the promotion of beneficial bacteria, such as lactic acid bacteria of pomelo peel essential oils and CTC [
34,
38].
4.5. Dietary PPP Affected Colonic Microflora
The microbial barrier in intestine is formed by the settlement of intestinal microorganisms in the mucosa to prevent excessive reproduction of pathogenic bacteria in the intestinal segment [
39]. Their main function is to maintain the balance of intestinal microecology, assist the immune cells and immune factors of intestinal mucosa to perform their functions, promote the development of intestinal immune system, and improve the immune ability of the body [
40].
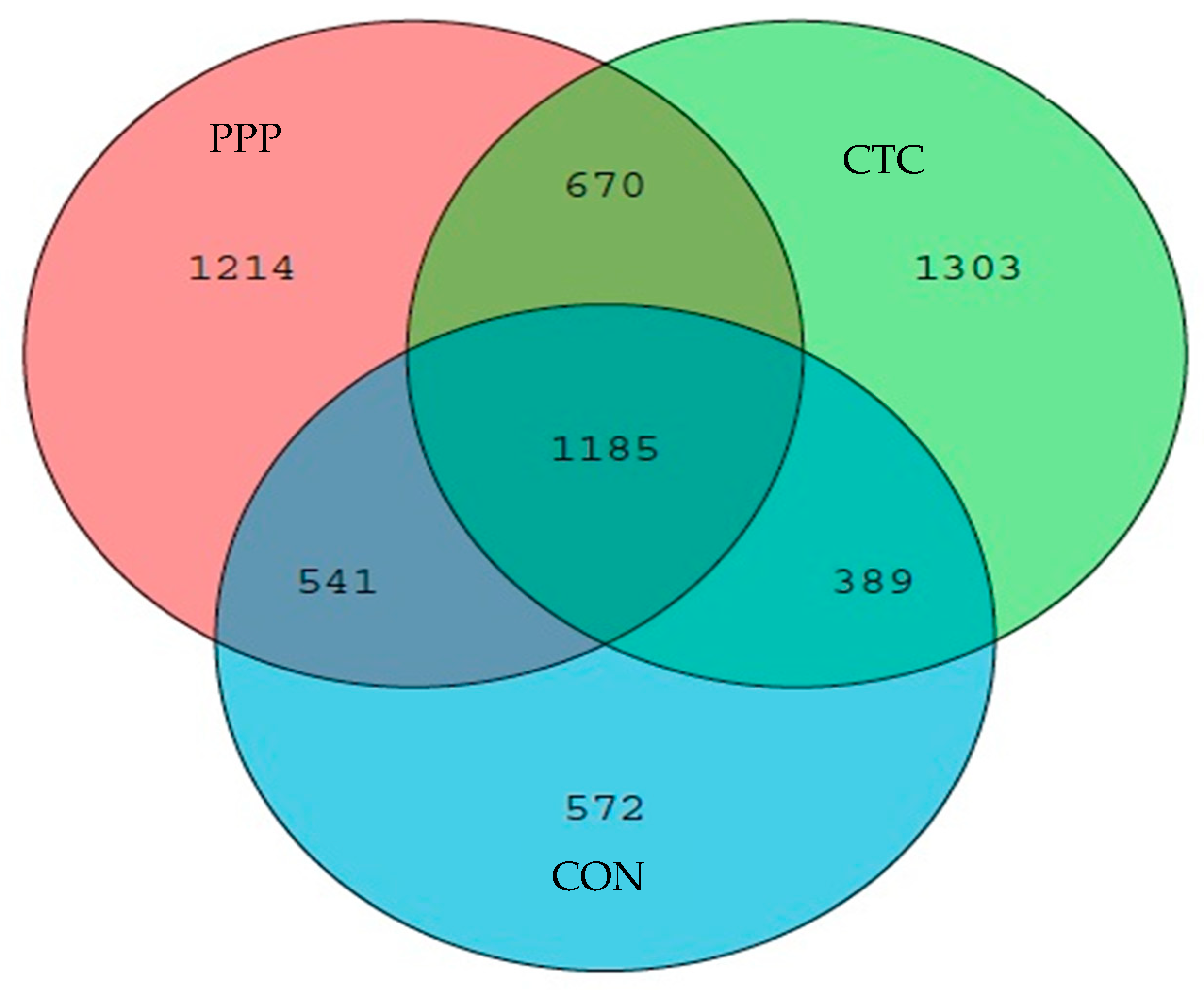
The nutrition of the intestinal flora comes from the dietary composition and exfoliated epithelial cells of the host, which is an organ with extensive metabolic capacity and substantial functional plasticity. In this experiment, the dominant phylum flora in the three groups contained
Firmicutes,
Bacteroidetes, and
Proteobacteria. In this experiment, the
Bacteroidetes relative abundance in PPP and CTC was significantly higher than those of CON, indicating that PPP and CTC can improve the colonic microbial of piglets. These characteristics of the intestinal flora make the focus of the study shift from the richness and diversity of microbial members to function. A clinical study by Olaisen et al. found that the
Bacteroidetes relative abundance in patients’ intestines with Crohn’s disease and ulcerative colitis decreased, indicating that the abundance of
Bacteroidetes was related to intestinal inflammation and immunity to a certain extent [
41].
In this experiment, the
Firmicutes relative abundance in PPP decreased significantly, indicating that PPP can control lipid synthesis and reduce fat synthesis and that PPP can be used to improve the lean meat percentage of pigs. The ratio of the relative abundance of
Firmicutes to that of
Bacteroidetes is related to obesity, and individuals with a high ratio are more likely to be obese [
42]. Weaning oxidative stress can increase the intestinal
Cyanobacteria relative abundance in weaned piglets [
43]. However, in this experiment, the
Cyanobacteria relative abundance decreased in CTC and PPP, indicating that the piglets in these two groups were relieved of their degree of weaning oxidative stress.
In this experiment, the dominant family bacteria in CON were Veillonellaceae, Lachnospiraceae, Ruminococcaceae, and BacteroidalesS24-7, while those in the antibiotic group were Prevotellaceae, Veillonellaceae, and Lachnospiraceae. The dominant bacteria in PPP were Prevotellaceae, Lachnospiraceae, and Ruminococcaceae species. The Prevotellaceae relative abundance in the antibiotic group was significantly higher than those in CON and PPP.
Prevotellaceae plays a major role in the digestion of starch, cellulose, and proteins [
44], indicating that CTC can improve intestinal microorganisms and improve the digestion and absorption of nutrients in the intestinal tract. The
Veillonellaceae relative abundance in PPP decreased significantly, and the
Veillonellaceae relative abundance increased when the body was infected, while the
Veillonellaceae relative abundance in PPP was significantly lower than in CON, the results showed that the intestinal infection of piglets in PPP was alleviated.
Ruminococcaceae ferments cellulose and pentosans into organic acids, which can inhibit pathogenic bacteria growth in intestines and protect the intestinal tract [
45]. The improvement in the
Ruminococcaceae relative abundance in PPP indicated that dietary PPP to the diet can help maintain the stability of intestinal microorganisms and exert normal digestion and absorption function in the intestinal tract.
Through functional prediction of colonic microflora, it was found that the microbial community of piglets supplemented with PPP had relative advantages in cell metabolic function (such as cell activity, cell growth, and apoptosis) and body metabolic function (such as lipids, amino acids, and terpenoid metabolism). CTC has a relative advantage in terms of its cell metabolic activity. It has been suggested that PPP may improve the microbial community in cell life processes and body metabolism. The above results show that CTC and PPP can improve colonic microflora and have a positive impact on health of piglets through colonic microorganisms and their products.



 ,
, 


{kind=link}
{kind=link}