1. Introduction
Fish reared under intensive aquaculture conditions are subjected to a wide variety of stressors. Between them, the nutritional imbalances may induce a chronic stress status [
1,
2,
3] compromising fish growth performance and impairing fish immune response and tissue integrity [
4,
5,
6,
7,
8].
Fish gills have essential functions for fish physiological balance, gas exchange, hydro–mineral balance [
9], and immune response [
10]. As gills interact directly with the external environment, they are the first barrier of protection against external agents such as pathogens and chemicals, acquiring a significant importance in fish development and disease resistance [
10,
11]. One of the main cell types composing gill epithelia are mitochondria-rich cells (MRCs), which are involved in gas exchange, ion transport, and blood acid–base balance regulation [
9].
As a direct consequence of a stressful process, cortisol will target gills, increasing oxidative phosphorylation to ensure the energy availability to conduct all the physiological changes required to cope with the stress processes and an up-regulating Na
+/K
+ ATPase pump activity to maintain tissue hydro–mineral balance and functioning [
12]. Cortisol effects are mediated through glucocorticoid receptors (GR), which reside in the cytoplasm complexed with the co-chaperone heat-shock proteins heat-shock protein 70 (HSP70) and heat-shock protein 90 (HSP90) [
13]. Cortisol binds to the GR, inducing the dissociation of the chaperon proteins; then, cortisol–GR complex translocates into the nucleus to regulate gene expression of different stress-responsive factors, such as the pro-inflammatory nuclear factor
κβ (
nfκβ2). The NF
κβ protein is one of the most important mediators of inflammatory response, which can be activated by different extracellular stimuli, such as pro-inflammatory cytokines [
14,
15], reactive oxygen species (ROS) [
15,
16], pathogen-associated molecular patterns (PAMPs) [
17,
18], and acute stress events [
19]. In parallel, the GR can bind to the BCl-2 receptor in the mitochondrial membrane, inducing an increase in the oxidative phosphorylation rate by the mitochondrial electron-transport chain (ETC), generating energy to supply the adenosine triphosphate (ATP) synthase to produce ATP [
20]. However, not all the electrons in the ETC are transferred to the final acceptor, generating an electron leak, which leads to the formation of reactive oxygen species (ROS)—namely, superoxide-radical (O
2−) formation. Superoxide radicals are transformed by superoxide dismutase (SOD) to hydrogen peroxide (H
2O
2), which diffuses to the cytoplasm to be detoxified by glutathione peroxidase (GPX) and catalase (CAT) [
21,
22].
Oxidative stress results from an imbalance between ROS production and its neutralization by the antioxidant-defense system. It leads to the oxidation of essential biomolecules such as proteins and lipids, DNA damage, and the impairment of mitochondrial activity, causing cell death [
21]. Insufficient ATP production will also impair Na
+/K
+ ATPase activity, causing hydro–mineral imbalances [
23,
24].
Functional ingredients have been profiled as suitable tools to face these harmful side effects, boosting fish health and promoting fish welfare, reinforcing fish immune response [
25,
26,
27,
28], attenuating fish stress response [
28,
29,
30,
31], and reducing oxidative stress processes [
32,
33,
34]. This is of remarkable interest in European sea bass, which is an especially susceptible fish species to stress processes [
35,
36,
37]. Among the functional ingredients, phytogenic feed additives (PFAs) are plant-derived bioactive compounds such as flavonoids, mucilages, and tannins with antioxidant properties [
38,
39,
40]. In previous studies, dietary supplementation with a mixture of garlic and labiate plant essential oils attenuated European sea-bass juveniles’ stress response, with the fish fed the supplemented diets presenting lower circulating cortisol levels in comparison to fish fed a control diet [
30,
41,
42]. In addition, PFA supplementation enhanced fish gut-mucosal health, reducing pre-ileorectal valve-segment goblet-cell size as compared to fish fed the control diet [
41]. Another example of plant-derived functional ingredients are prebiotics, which are indigestible fibers with the ability to enhance host health by selectively stimulating the growth and activity of a limited number of intestinal bacterial species [
28,
43,
44,
45,
46]. Between them, galactomannan–oligosaccharides (GMOS) have demonstrated in previously reported studies to increase host antioxidant capacity, modulate gut microbiota, and promote gut health in this fish species [
41,
42,
47].
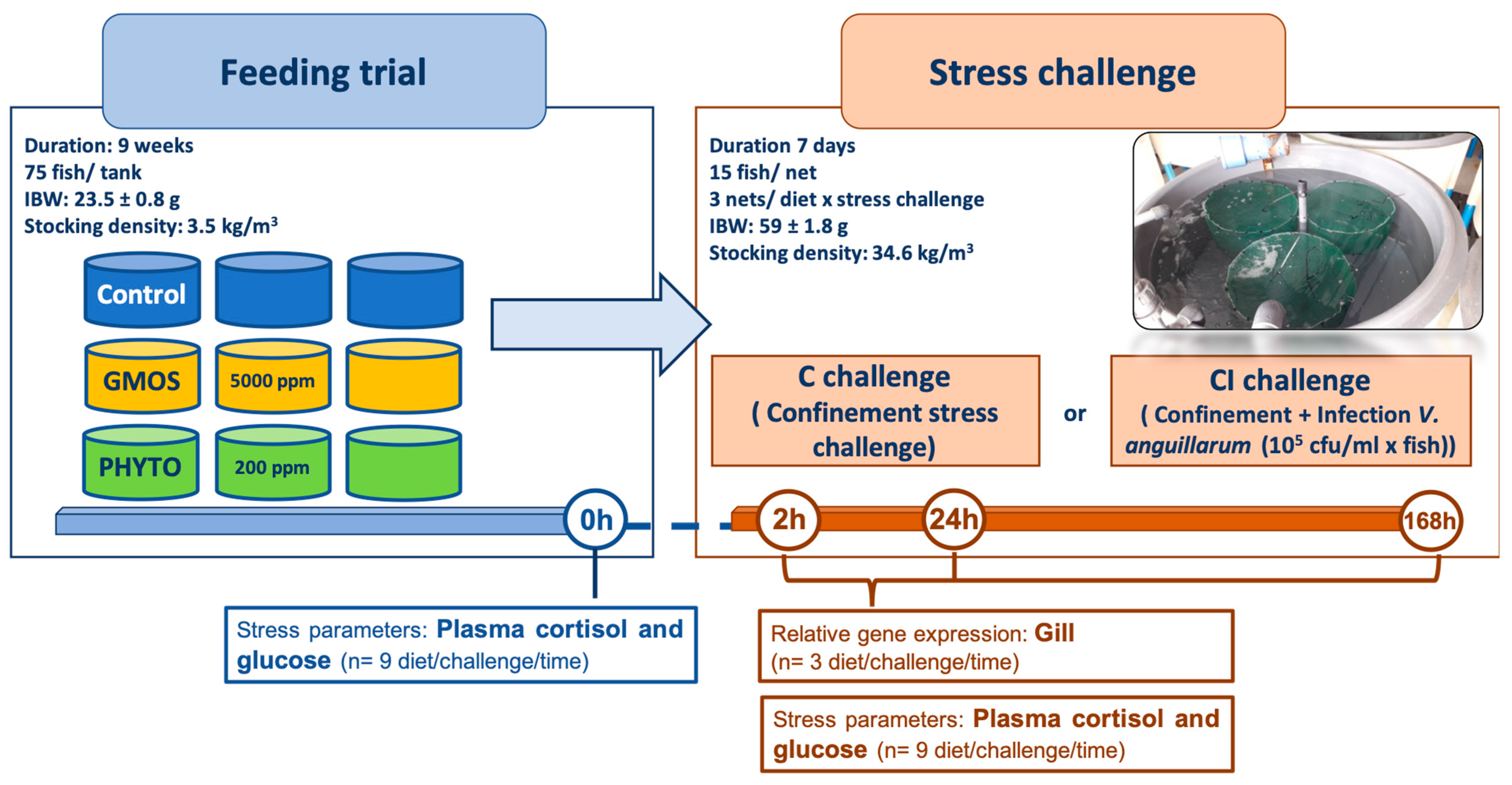
A scarce number of studies have investigated the effects of functional ingredients to offset the negative effects derived from low-FM/FO diet formulation and especially in fish subjected to stress processes. Thus, the aim of this study is to evaluate the effects of functional additives (PFAs and GMOS) as potential boosters of the gill endogenous antioxidant capacity of European sea-bass juveniles fed low-fish meal (FM)/fish oil (FO)-based diets when challenged against physical and biological stressors.
3. Results
As reported in our previous studies [
44], fish grew properly during the feeding trial with no significant effects on fish growth performance associated with the use of the functional diets. After the nine-week feeding trial, the fish presented a mean increase of 2.6× body weight, representing a relative growth (%) of 158.8 ± 16.3. During the feeding trial, no mortality was registered in any of the specific dietary treatments.
Functional-ingredient dietary inclusion did not induce any differences on fish basal (t = 0 h) stress parameters, with values ranging from 4.67 to 5.82 ng/mL for circulating plasma cortisol and from 67.43 to 67.71 mg/dL for circulating plasma glucose (
Table 3). At 2 h after crowding stress a generalized increase (
p < 0.05) in circulating plasma-cortisol concentration was observed, with significantly higher (
p < 0.05) values in those fish fed the control diet. In the early hours after C challenge (2 h), all the dietary treatments presented an increase in circulating cortisol levels, especially those fish fed the control diet, with significantly higher (
p < 0.05) levels than fish fed GMOS and PHYTO diets (
p < 0.05). At 2 h after CI challenge, a similar trend was observed, with fish fed GMOS presenting lower (
p < 0.05) cortisol levels than fish fed the control diet. On the contrary, at 24 h after CI challenge fish fed GMOS and PHYTO diets presented significantly higher (
p < 0.05) circulating plasma-cortisol levels than those fed the control diet (
Table 3). At the end of the CI challenge (t = 168 h), fish fed the PHYTO diet presented higher (
p < 0.05) levels of circulating plasma cortisol than fish fed the control diet. Regarding circulating plasma-glucose concentrations, the use of functional additives did not induce significant differences in fish pattern of response against crowding stress (C challenge). Meanwhile, at 2 h and 168 h after the CI challenge, fish fed GMS and PHYTO diets presented significantly higher (
p < 0.05) plasma-glucose levels than fish fed the control diet (
Table 3).
At the end of the stress challenge, fish subjected to the confinement (C challenge) did not present mortality regardless of dietary treatment. Nevertheless, the combination of both confinement and the pathogen gut inoculation (CI challenge) resulted in a relative percentage of survival (RPS = [1 – (%) surviving fish fed functional diet/(%) surviving fish fed control diet] × 100) of 47% and 33% in fish fed the PHYTO and GMOS diets, respectively (
Figure 2), compared to fish fed the control diet [
45].
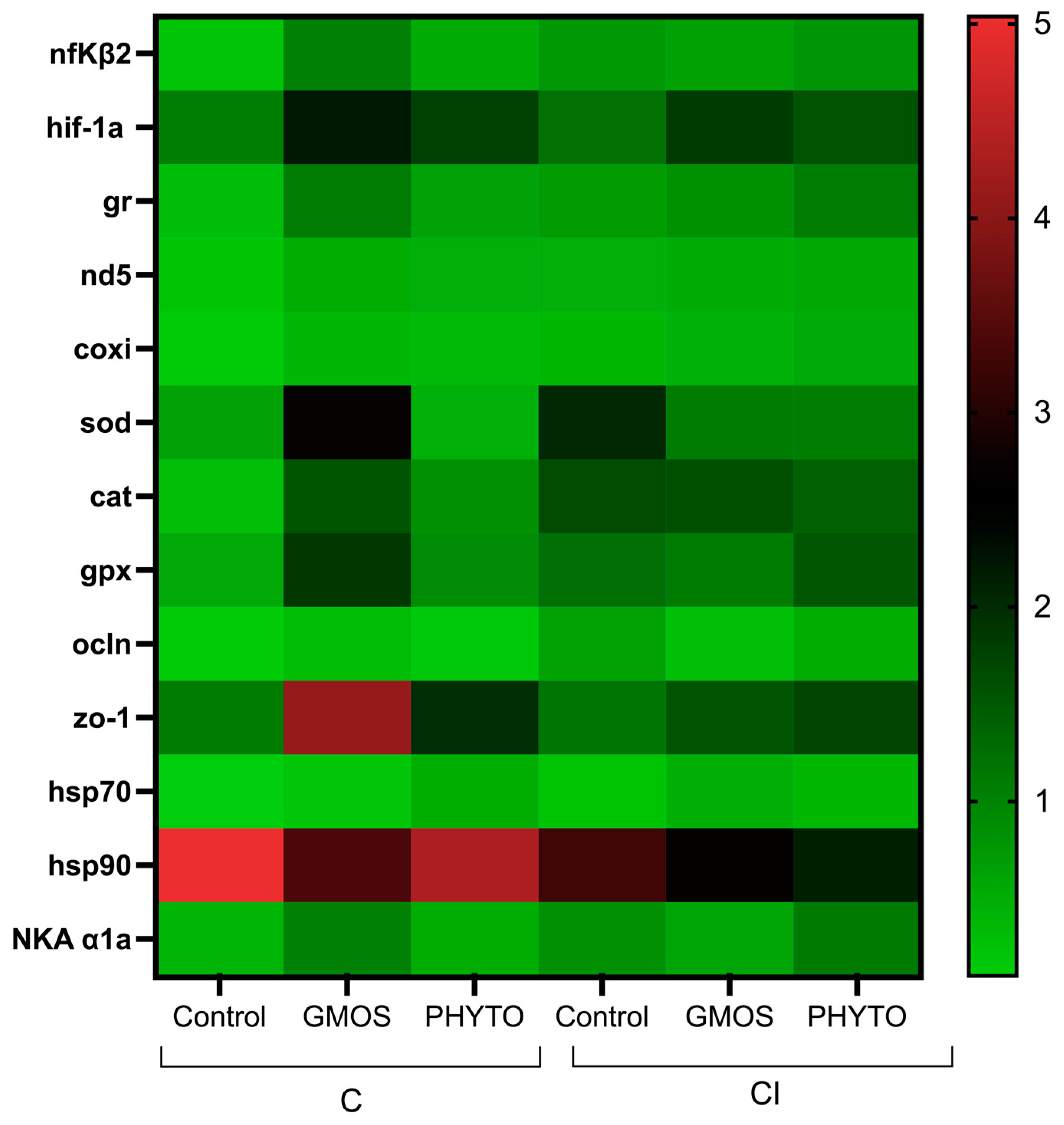
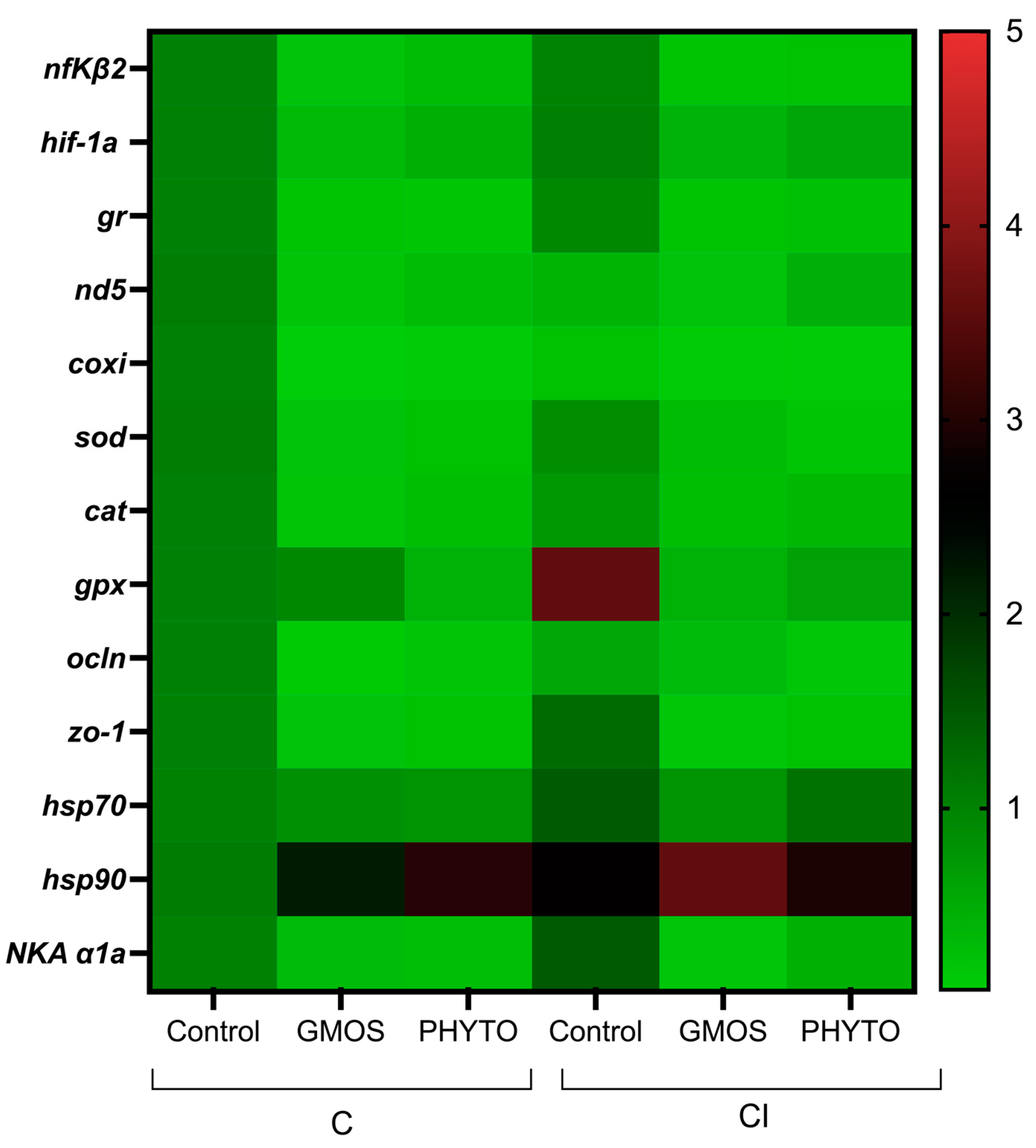
When fish were subjected to the stress challenge, two hours after confinement (C challenge) those fed the control diet presented significantly higher (
p < 0.05) gill-transcript levels of
nfκβ2,
hif-1α,
gr,
nd5,
coxi,
sod,
cat,
hsp70,
hsp90, and
NKA α1a genes than those fed GMOS and PHYTO diets (
Table 4,
Figure 3). No differences were found among fish fed the different dietary treatments and subjected to C challenge for
gpx,
ocln, and
zo-1 relative gene expression. Similarly, two hours after the CI challenge, fish fed the control diet presented a significant (
p < 0.05) up-regulation of
nfκβ2,
hif-1α,
gr,
sod,
gpx,
NKA α1a, and
hsp90 gill gene expression compared to gills of fish fed GMOS and PHYTO diets. At this sampling point, fish fed the control diet and subjected to the CI challenge presented significantly lower (
p < 0.05)
nd5 and
coxi gill transcript levels than those fed the same diet but subjected to the C challenge.
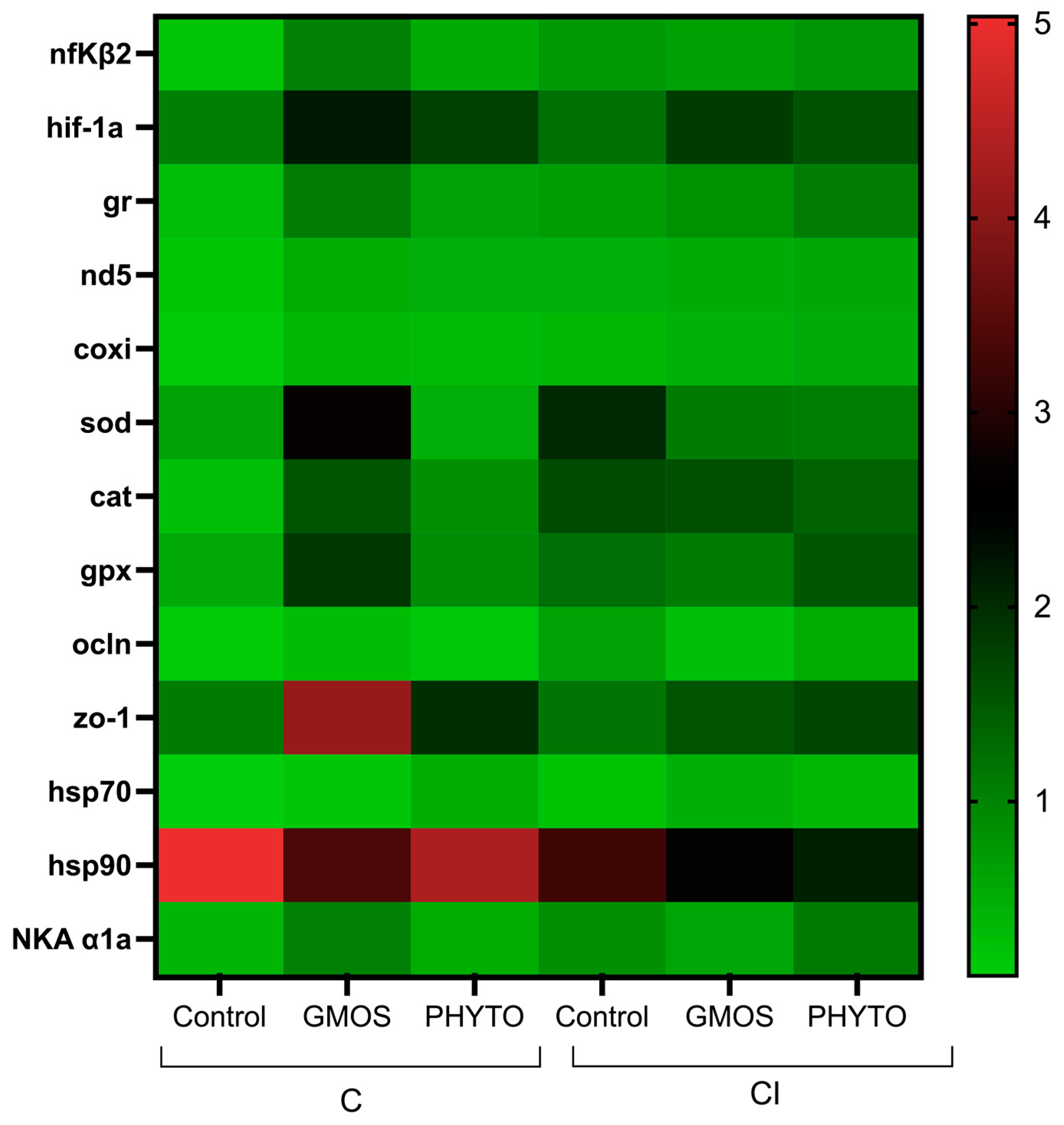
At 24 h after confinement and in relation to the gene-expression levels observed after 2 h of confinement, fish fed the control diet presented a significant down-regulation (
p < 0.05) of
nfκβ2,
gr,
nd5,
coxi, and
hsp70 and an up-regulation (
p < 0.05) of
zo-1 gill relative gene expression compared to the initial transcript levels at 2 h after confinement (
Figure 4,
Table 5). On the contrary, fish fed the GMOS diet presented an up-regulation (
p < 0.05) of
nfκβ2,
hif-1α,
gr,
sod,
cat,
hsp70, and
hsp90 gill relative gene expression and fish fed the PHYTO diet presented an up-regulation of
hif-1α compared to the previous sampling point at 2 h.
For fish confined and infected after 24 h post challenge the pattern of response for all the dietary treatments resulted in a down-regulation (p < 0.05) of ocln gill relative gene expression compared to the previous sampling point, whereas coxi and cat transcript levels presented a significant up-regulation (p < 0.05) compared to the initial transcript levels. Besides, fish fed the control and PHYTO diets presented an up-regulation (p < 0.05) of sod gill relative gene expression compared to 2 h post CI challenge. Fish fed the GMOS diet, and in relation to 2 h post CI challenge, presented a significant up-regulation (p < 0.05) of hif-1α gill transcript levels, whereas fish fed the PHYTO diet presented an up-regulation (p < 0.05) of nfκβ2, gr, and NKA α1a gill gene expression compared to 2 h post CI challenge.
Within the 24 h sampling point for the C challenge, fish fed the GMOS diet presented higher (p < 0.05) gill gene-transcript levels of nfκβ2, sod, gpx, and hsp90 than those fed the control diet, as well as higher (p < 0.05) sod gill relative gene-expression values than those fed the PHTYO diet. Similarly, fish fed the PHYTO diet presented higher (p < 0.05) gill relative gene expression of ocln than those fed the control diet. On the other hand, in fish subjected to the CI challenge after 24 h of stress challenge, the diet did not induce significant differences in the gill relative gene expression of the target genes, despite both stress challenges differing between them in the gill relative gene-expression patterns presented. At 24 h post CI challenge, those fish fed the control diet presented higher (p < 0.05) sod and hsp70 gene-expression levels than those fed the same dietary treatment but subjected to the C challenge. On the contrary, the CI challenge induced a significant down-regulation (p < 0.05) of sod gill transcript levels in fish fed the GMOS diet and subjected to the C challenge. Regardless of the dietary treatment, the CI challenge induced a significant down-regulation (p < 0.05) of zo-1 relative gene expression compared to C challenge after 24 h post challenge.
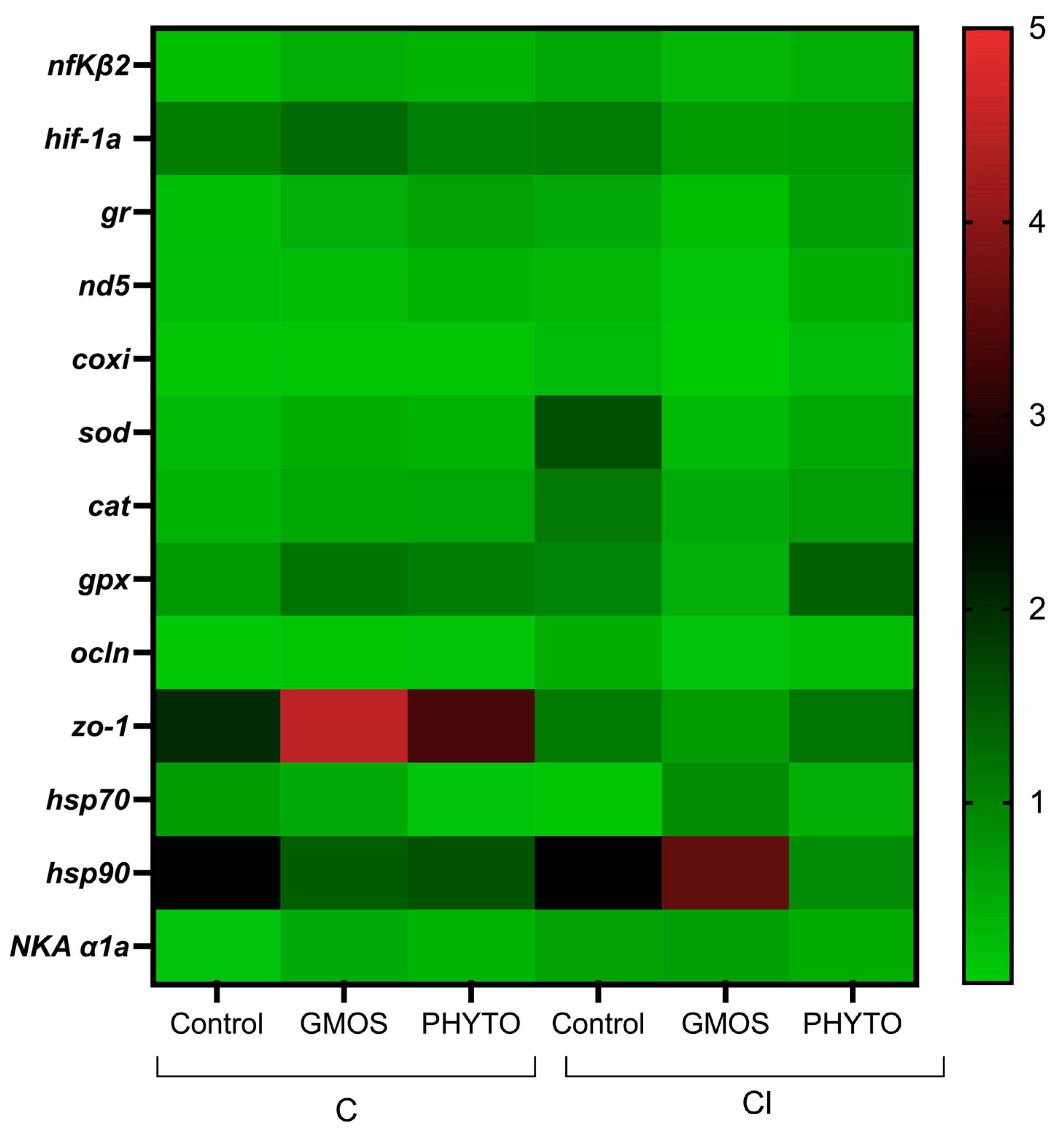
At the end of the confinement stress challenge (168 h after C challenge), fish fed the GMOS diet presented a significant down-regulation (
p < 0.05) of
sod and
cat gill transcript levels (
Table 6,
Figure 5) down to the levels observed at 2 h post challenge, as presented by fish fed the PHYTO diet for
zo-1 gill relative gene expression. Fish fed the control diet and confined presented a significant down-regulation (
p < 0.05) of
NKA α1a gill relative gene expression compared to the previous sampling points at 2 h and 24 h post C challenge. In the case of the CI challenge, fish fed GMOS presented a significant down-regulation (
p < 0.05) of sod gill gene-expression levels down to the initial levels.
Within this last sampling point (168h), fish from the C challenge and fed GMOS diet presented higher (p < 0.05) NKA α1a gill relative gene expression than those fed the control diet, whereas fish fed the PHYTO diet presented lower (p < 0.05) gill gene expression ocln than those fed the control diet. Regarding the CI challenge, fish fed the control diet presented higher (p < 0.05) gill sod transcript levels than fish fed the GMOS and PHYTO diets, whereas fish fed the control and PHYTO diet presented higher (p < 0.05) gr gill relative gene-expression values than those presented by fish fed the GMOS diet. On the contrary, fish fed the GMOS diet presented higher (p < 0.05) ocln and zo-1 gill gene-expression levels than fish fed the control and PHYTO diets, respectively.
At 168 h after CI challenge, the infection itself induced significantly higher (p < 0.05) sod, NKA α1a, and hsp70 gill gene-transcript levels, whereas it down-regulated (p < 0.05) ocln gill gene expression. In the case of the fish fed the GMOS diet, the CI challenge significantly increased (p < 0.05) ocln and zo-1 gill relative gene expression and reduced (p < 0.05) hsp90 in comparison to fish subjected to the C challenge. No effects were detected for fish fed the PHYTO diet when comparing the C and CI challenges at the end of the stress trial.
4. Discussion
Stress induces a physiological response to reestablish fish homeostasis, which is orchestrated by the release of cortisol into the bloodstream. As expected, in the present study an increased concentration of fish circulating plasma concentration was observed in the few hours after the initiation of both stress challenges and regardless of the dietary treatment. However, both GMOS and PHYTO diets attenuated fish stress response, with supplemented fish presenting lower circulating-cortisol levels than fish fed the control diet.
In response to the stress process, the organism undergoes an alarm status in which the energetic resources are rearranged in order to cope with the surplus of activity. In this sense, gills play a fundamental role in energy supply with the activation of ATP synthesis [
20,
51]. In the present study, and in agreement with the attenuated stress response observed in cortisol patterns for supplemented fish, at 2 h after both the C and CI challenge, fish fed the GMOS and PHYTO diets presented lower
gr,
nd5, and
coxi gill relative gene expression than fish fed the control diet. This may suggest a lower responsiveness of fish fed functional diets against the stressor, with a lower activity of the ETC and thus lower energetic requirements to cope with the stress process [
51]. Nevertheless, fish fed the GMOS and PHYTO diets presented an up-regulation of
gr relative gene expression at 24 h after the stress challenges. This delayed increase in
gr gene expression in fish fed the supplemented diets could be understood as a mechanism to restore the GR protein content after exposure to glucocorticoids. Vijayan t al. (2013) reported on an in vitro experiment using hepatocytes, in which down-regulated
gr gene expression was found in treatments presenting the higher GR protein contents [
52].
Considering that the ETC is one of the major sources of endogenous ROS [
20,
34,
53], an increased aerobic-metabolism rate in response to a stress process may induce elevated ROS production, leading to oxidative stress [
21]. In the present study, at 2 h after the C challenge, fish fed the functional diets presented lower
sod and
cat gill gene-expression levels. A lower activation of the endogenous antioxidant defenses could suggest an attenuated stress response, leading to a lower production of mitochondrial ROS and thus an attenuated response against the stress processes. Although both functional diets reduced fish-stress and oxidative-stress response, each functional diet induced a different antioxidant response against the confinement stress challenge. In fact, along with the stress challenge, fish fed PHYTO did not presented an activation of endogenous antioxidant machinery gene expression. This lack of response in the endogenous antioxidant defenses could be present and associated with the antioxidant properties of the phytogenic compounds, making them capable of inhibiting ROS formation and quenching them once they are formed [
54,
55,
56]. Indeed, our previous studies in this mucosal tissue indicated that both functional additives can reduce gill oxidative-stress status in basal conditions after dietary supplementation [
42]. Between the different plant-origin biomolecules, the flavonoids are polyphenolic compounds with high antioxidant properties and can be found in a wide spectrum of plant extracts as garlic oil and labiate plant extracts, such as origanum [
57] and thyme [
58]. On the other hand, fish fed the GMOS diet presented an increase in
sod and
cat gill gene expression at 24 h after the C challenge. In addition, fish fed the GMOS diet presented higher
gpx gill transcript levels than fish fed the other dietary treatments. Interestingly, our previous studies indicated a similar delayed pattern of response against the stressor in other mucosal tissues, inducing GMOS supplementation as a controlled and prolonged intestinal-mucus secretion in response to the CI challenge, reducing gut bacterial-translocation rates and thus increasing pathogen resistance and survival rates [
41]. Similarly, in the present study both functional diets also attenuated fish antioxidant-related gill gene response against the CI challenge, with fish fed the functional diets presenting lower values of
sod and
gpx gene expression than fish fed the control diet in the early hours after bacterial gut inoculation. Nevertheless, 24 h after the CI challenge the functional-diet supplementation induced an up-regulation of
sod,
cat, and
gpx gill gene expression. This increase in the expression of antioxidant-related genes might suggest a response against the stress associated with the pathogen gut inoculation and with an organism arrangement to cope with future infection. In particular, dietary PHYTO increased fish
gpx gene expression and kept it up-regulated throughout the entire stress challenge. Similarly, Mansour et al. (2020) observed a higher antioxidant capacity in gilthead sea-bream (
Sparus aurata) gills and skin, with an increased gene expression of
sod and
cat after feeding diets supplemented with
Moringa oleifera leaf extracts [
59].
The gills, skin, and intestine are the first barrier interacting with the external environment, playing a fundamental role in maintaining tissue structure and integrity, regulating solute trafficking across the gill epithelium and thus facilitating or limiting paracellular-ion movement [
60]. For this reason, cell-junction complexes play a fundamental role in maintaining gill-epithelium integrity and functioning. Damage to fish-gill structural integrity like that originated by oxidative-stress processes may lead to degenerative processes such as gas-exchange disturbances [
61] and impairments to immune functions [
62]. In our previous studies, Torrecillas et al. (2021) observed the ability of GMOS and PHYTO diets to induce higher gill gene expression of
zo-1 and
ocln, respectively, in basal conditions [
42]. In the present study, the use of different dietary treatments did not affect fish gill
ocln and
zo-1 relative gene expression in the early hours after both stress challenges. However, 24 h after confinement, fish fed all the dietary treatments presented an increase in
zo-1 gill gene expression despite it only being significant in fish fed the control diet, with fish fed PHYTO diet at 24 h post confinement presenting the highest expression levels. In concordance with these results, Zhao et al. (2020) described an up-regulated gene expression of
ocln and
zo-1 genes in the gills of
Ctenopharyngodon idella-fed diets supplemented with
Allium monoglicum Regel flavonoids (AMRF), alleviating the oxidative stress and toxicity derived from chromium exposure [
56]. In the same way, Trujillo et al. (2015), described the ability of curcumin to prevent cisplatin-induced fibrosis and decreased tight-junction proteins in rat kidney [
63]. These protective effects of phytogenic feed additives could be related to the ability of those plant-origin compounds to interact with MAPK receptors, preventing H
2O
2-induced tight-junction disruptions [
64]. On the contrary, at 24 h after challenge when the fish were subjected to the CI challenge and at 24 h after stress all the dietary treatments presented a down-regulation of
ocln gill gene expression. Moreover, the CI challenge induced a down-regulation of
zo-1 compared to the C challenge. Acute inflammatory processes are characterized by the hyper-permeabilization of tissues, allowing inflammatory mediators and immune cells to infiltrate the damaged tissues [
65]. In this sense, a down-regulation of genes related to tight-junction structure maintenance could be related to a preparation process to facilitate the response against a future infection. The inflammatory response acquires a critical importance in the gills, considering the high number of permanent-resident lymphocytes and immune cells associated with the gill-associated lymphoid tissue (GIALT) [
66]. In the present study, both the C and CI challenge induced an acute response of
nfκβ2 gill gene expression after stress challenge in fish fed the control diet, whereas fish fed the GMOS and PHYTO diets presented an attenuated pro-inflammatory response against the stressors, with the highest
nfκβ2 gill gene-expression levels at 24 h after being subjected to the C and CI challenge. Previous studies have reported on the ability of phytogenic compounds derived from oregano, curcumin, and thymol to modulate pro-inflammatory response in fish [
67,
68,
69]. These compounds have been shown to be able to directly regulate the NFKB- and mitogen-activated protein kinase (MAPK)-signaling pathways, attenuating the inflammatory response [
27]. In the case of GMOS, the mechanism that can modulate European sea-bass immune and stress response differs from the phytogenic compounds, as the animal does not directly harness the prebiotics. The by-products from prebiotic fermentation generated by the host microbiome may produce short-chain fatty acids (SCFAs), which can modulate fish innate immune response and inflammatory cells [
70] by interacting with immune-cell pattern-recognition receptors [
71]. Inflammatory processes are characterized by an increased leukocyte infiltration [
5], which may lead to hypoxia conditions due to the high amount of O
2 consumed by the increased phagocytic activity [
72]. In response to the hypoxia, the
nfκβ2 triggers the activation of the
hif-1α, inducing a metabolic swift into a glycolytic strategy, facilitating leukocyte survival in a hypoxic medium [
73]. In the present study, fish fed the functional diets presented lower
hif-1α gill gene-expression levels than those fed the control diet, supporting the idea of an attenuated pro-inflammatory response in the early hours after the stress. Nevertheless, at 24 h after stress challenge, and in parallel with an increased transcription of the
nfκβ2, both functional diets induced an up-regulation of
hif-1α gill gene expression. This could suggest a better protection of the immune-cell populations, leading to a better ability to cope with the deleterious effects derived from a prolonged inflammatory response against a stressor, which may also be related with the lower infection rates and higher survival observed in CI fish fed the functional diets [
41]. Indeed, the fish fed the control diet presented no significant changes in
hif-1α gill gene-expression levels regardless of the variations in the
nfκβ2 gene transcripts during the different stress challenges. A similar response was observed in previous reports, in which the same functional diets protected head-kidney leukocyte populations against apoptotic processes by attenuating head-kidney pro-inflammatory response and increasing
hif-1α head-kidney relative gene expression after CI challenge [
30].
Another mechanism promoting tissue integrity in response to a stressor is the activation of the heat-shock proteins, which are overexpressed to act as molecular chaperones associated with the GR avoiding protein denaturation, refolding denatured proteins and promoting misfolded-protein degradation [
74,
75,
76]. In response to an acute stress process, the
hsp70 and
hsp90 gene expression is increased, activating the necessary mechanisms to respond to the stressor [
77]. In the present study, in fish fed the control diet both C and CI challenges induced an overexpression of gill
hsp70 in the first hours after the challenge, followed by a strong down-regulation until the end of the stress challenges. Meanwhile, the fish fed the GMOS diet presented a delayed
hsp70 and
hsp90 gene-expression pattern, with the highest expression levels at 24 h after stress, indicating an attenuated response to the stress. In concordance, in previous studies dietary supplementation with fructo-oligosaccharides in blunt-snout bream (
Megalobrama amblycephala) induced an increase in
hsp70 and
hsp90 at 24 hours after confinement stress [
78].
Na
+/K
+ ATPase is an ATP-dependent transmembrane enzyme that plays a fundamental role in maintaining cell ionic homeostasis. This protein is highly represented in the gills and confers an important osmoregulatory role to the tissue [
79]. In the present study, at 2 h after C and CI challenges, the fish fed the control diet presented a strong up-regulation of
NKA α1a gill gene expression, indicating an acute response to the stressor. At the end of the confinement stress challenge, these fish presented a down-regulated gene expression of the
NKA α1a. Alterations to cellular ionic balance may lead to the entrance of sodium (Na
+) [
23,
80] and thus disturb the osmotic balance, leading to membrane ruptures [
43]. When fish were subjected to the CI challenge, the control group presented the same pattern of response but the
NKA α1a gene expression remained unchanged throughout the stress challenge, being highest at the end of the CI challenge. Meanwhile, and regardless of the stress challenge, fish fed the functional diets presented the higher values of relative gene expression of
NKA α1a at 24 h after stress, indicating an attenuated response to the stressor. The fish fed the functional dies did not show a down-regulation of
NKA α1a, which could suggest a more prolonged activity of the Na
+/K
+ ATPase and thus a better capacity to cope with the imbalances originated during the stress process, which in turn may also be related to the lower infection rates observed in supplemented fish.



 ,
, 





{kind=link}
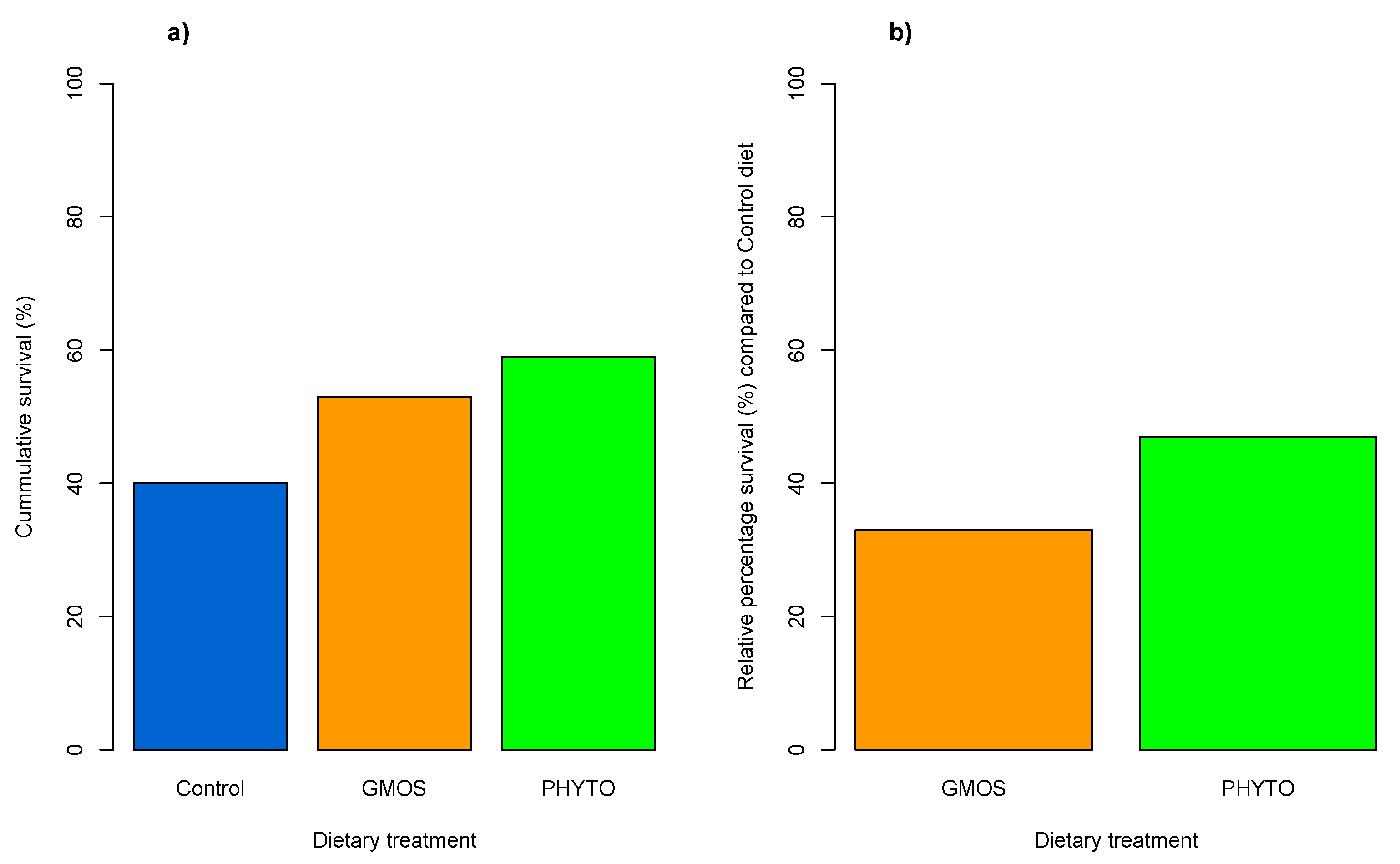{kind=link}
{kind=link}
{kind=link}
{kind=link}