
Avian Alarm Calls Do Not Induce Anti-Predator Response in Three Anuran Species


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
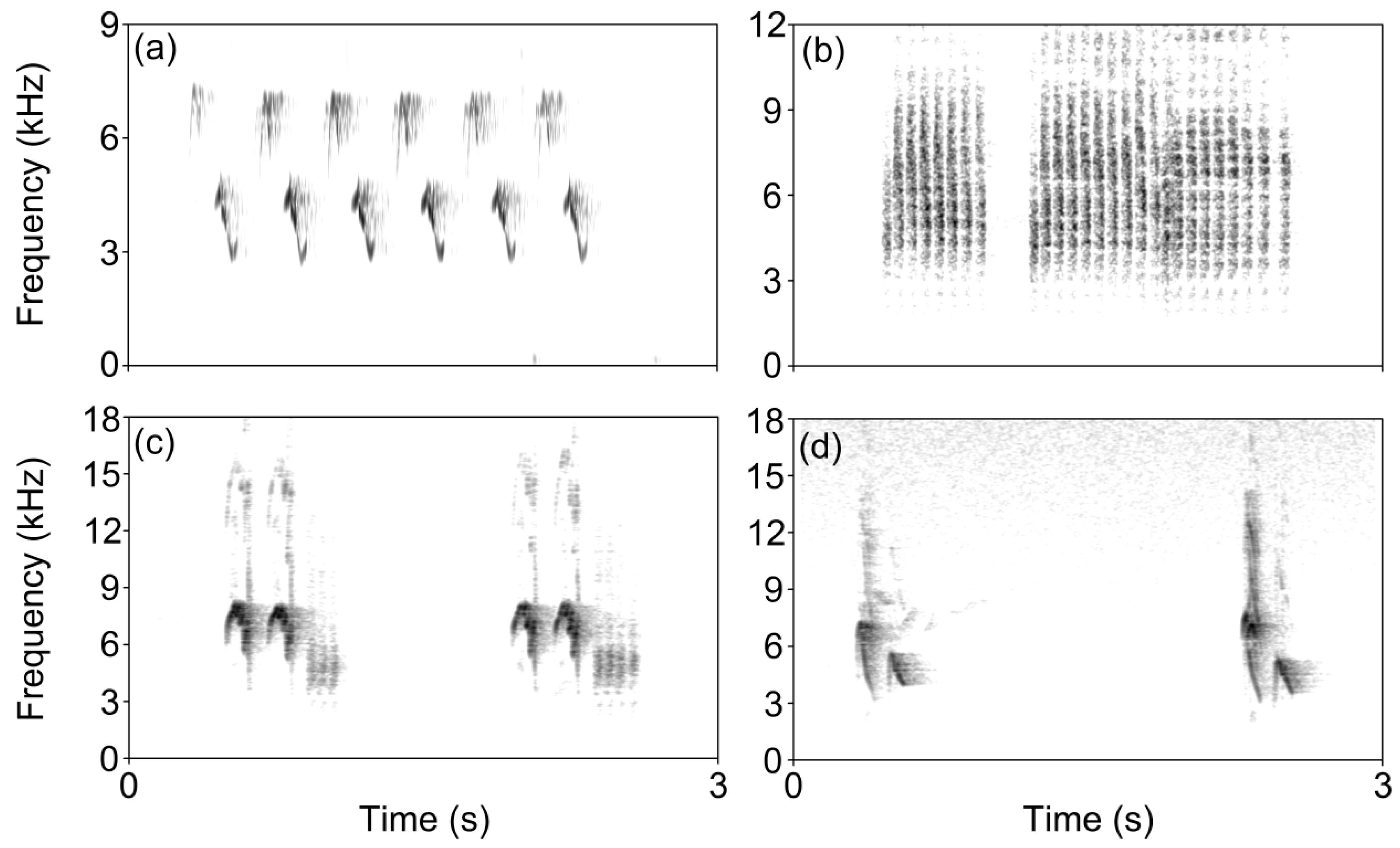
2.2. Sound Designs
2.3. Sound Playback Experiments
2.4. Data Analyses
3. Results
3.1. Responses of Little Torrent Frogs to Sound Playbacks
3.2. Responses of Ornamented Pygmy Frogs and Spot-Legged Treefrogs to Sound Playbacks
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dall, S.R.X.; Giraldeau, L.-A.; Olsson, O.; McNamara, J.M.; Stephens, D. Information and its use by animals in evolutionary ecology. Trends Ecol. Evol. 2005, 20, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.H.; Danchin, E. A taxonomy of biological information. Oikos 2010, 119, 203–209. [Google Scholar] [CrossRef]
- Magrath, R.D.; Haff, T.M.; Fallow, P.M.; Radford, A.N. Eavesdropping on heterospecific alarm calls: From mechanisms to consequences. Biol. Rev. 2015, 90, 560–586. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, J.W.; Vehrencamp, S.L. Principles of Animal Communication, 2nd ed.; Sinauer: Sunderland, UK, 2011. [Google Scholar]
- Klump, G.M.; Shalter, M.D. Acoustic behavior of birds and mammals in the predator context. 1. Factors affecting the structure of alarm signals. 2. The functional-significance and evolution of alarm signals. Z. Tierpsychol. 1984, 66, 189–226. [Google Scholar] [CrossRef]
- Zuberbühler, K. Survivor signals: The biology and psychology of animal alarm calling. Adv. Stud. Behav. 2009, 40, 277–322. [Google Scholar]
- Suzuki, T.N. Communication about predator type by a bird using discrete, graded and combinatorial variation in alarm calls. Anim. Behav. 2014, 87, 59–65. [Google Scholar] [CrossRef]
- Yu, J.; Xing, X.; Jiang, Y.; Liang, W.; Wang, H.; Møller, A.P. Alarm call-based discrimination between common cuckoo and Eurasian sparrowhawk in a Chinese population of great tits. Ethology 2017, 123, 542–550. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yu, J.; Shen, C.; Yin, D.; Jin, L.; Liang, W.; Wang, H. Geographic Variation in Note Types of Alarm Calls in Japanese Tits (Parus minor). Animals 2022, 12, 2342. [Google Scholar] [CrossRef]
- Suzuki, T.N. Parental alarm calls warn nestlings about different predatory threats. Curr. Biol. 2011, 21, R15–R16. [Google Scholar] [CrossRef] [Green Version]
- Templeton, C.N.; Greene, E.; Davis, K. Allometry of Alarm Calls: Black-Capped Chickadees Encode Information about Predator Size. Science 2005, 308, 1934–1937. [Google Scholar] [CrossRef] [Green Version]
- Leavesley, A.J.; Magrath, R.D. Communicating about danger: Urgency alarm calling in a bird. Anim. Behav. 2005, 70, 365–373. [Google Scholar] [CrossRef]
- Dutour, M.; Walsh, S.L.; Ridley, A. Australian magpies adjust their alarm calls according to predator distance. Bioacoustics 2021, 30, 458–468. [Google Scholar] [CrossRef]
- Blumstein, D.T. Alarm calling in three species of marmot. Behaviour 1999, 136, 731–757. [Google Scholar] [CrossRef]
- Ridley, A.R.; Raihani, N.J. Facultative response to a kleptoparasite by the cooperatively breeding pied babbler. Behav. Ecol. 2007, 18, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Flower, T. Fork-tailed drongos use deceptive mimicked alarm calls to steal food. Proc. R. Soc. B Boil. Sci. 2011, 278, 1548–1555. [Google Scholar] [CrossRef]
- Ridley, A.R.; Wiley, E.M.; Thompson, A. The ecological benefits of interceptive eavesdropping. Funct. Ecol. 2014, 28, 197–205. [Google Scholar] [CrossRef]
- Dutour, M.; Danel, S. Wild great tits’ alarm calls prompt vigilant behaviours in free-range chickens. Anim. Cogn. 2020, 24, 213–216. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, C.; Liu, J.; Liang, W. Female cuckoo calls elicit vigilance and escape responses from wild free-range chickens. Ethol. Ecol. Evol. 2020, 33, 37–48. [Google Scholar] [CrossRef]
- Zhang, C.; Jiang, X.; Li, M.; Liang, J.; Liu, J.; Liang, W. Female cuckoo calls elicit anti-predatory behavior in birds. J. Ethol. 2021, 39, 393–398. [Google Scholar] [CrossRef]
- Shriner, W.M. Yellow-bellied marmot and golden-mantled ground squirrel responses to heterospecific alarm calls. Anim. Behav. 1998, 55, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, U.; Coss, R.G. Recognition of heterospecific alarm vocalizations by bonnet macaques (Macaca radiata). J. Com. Psychol. 2000, 114, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Magrath, R.D.; Haff, T.M.; McLachlan, J.; Igic, B. Wild Birds Learn to Eavesdrop on Heterospecific Alarm Calls. Curr. Biol. 2015, 25, 2047–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potvin, D.A.; Ratnayake, C.P.; Radford, A.N.; Magrath, R.D. Birds Learn Socially to Recognize Heterospecific Alarm Calls by Acoustic Association. Curr. Biol. 2018, 28, 2632–2637.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, K.D. The Ecology and Behavior of Amphibians; The University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Phelps, S.M.; Rand, A.S.; Ryan, M.J. The mixed-species chorus as public information: Túngara frogs eavesdrop on a heterospecific. Behav. Ecol. 2007, 18, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Yin, D.; Yu, J.; Zhang, L.; Han, Z.; Jin, L.; Liang, W.; Wang, H. Geographical variation in nest defense among cinereous tit populations in China. Curr. Zool. 2022; in press. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, J.; Zhang, H.; Wang, T.; Yang, Y.; Tang, Y.; Halfwerk, W.; Cui, J. Parasite defensive limb movements enhance acoustic signal attraction in male little torrent frogs. eLife 2022, 11, e76083. [Google Scholar] [CrossRef]
- Zhou, B.; Liang, W. Avian escape responses to observers wearing clothing of different colors: A comparison of urban and rural populations. Glob. Ecol. Conserv. 2020, 22, e00921. [Google Scholar] [CrossRef]
- Zhao, L.; Zhu, B.; Wang, J.; Brauth, S.E.; Tang, Y.; Cui, J. Sometimes noise is beneficial: Stream noise informs vocal communication in the little torrent frog Amolops torrentis. J. Ethol. 2017, 35, 259–267. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing. 2021. Available online: http://www.r-project.org/ (accessed on 19 May 2021).
- Vitousek, M.N.; Adelman, J.; Gregory, N.C.; Clair, J.S. Heterospecific alarm call recognition in a non-vocal reptile. Biol. Lett. 2007, 3, 632–634. [Google Scholar] [CrossRef] [Green Version]
- Ito, R.; Mori, A. Vigilance against predators induced by eavesdropping on heterospecific alarm calls in a non-vocal lizard Oplurus cuvieri cuvieri (Reptilia: Iguania). Proc. R. Soc. B 2010, 277, 1275–1280. [Google Scholar] [CrossRef] [Green Version]
- Ito, R.; Ikeuchi, I.; Mori, A. A day gecko darkens its body color in response to avian alarm calls. Curr. Herpetol. 2013, 32, 26–33. [Google Scholar] [CrossRef]
- Wang, J. Wild Vertebrate in Diaoluoshan, Hainan, China; China Forestry Publishing House: Beijing, China, 2014. [Google Scholar]
- Hödl, W.; Amézquita, A. Visual signaling in anuran amphibians. In Anuran Communication; Ryan, M.J., Ed.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2001; pp. 121–141. [Google Scholar]
- Zhao, L.; Santos, J.C.; Wang, J.; Ran, J.; Tang, Y.; Cui, J. Noise constrains the evolution of call frequency contours in flowing water frogs: A comparative analysis in two clades. Front. Zool. 2021, 18, 37. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, J.; Cai, Y.; Ran, J.; Brauth, S.E.; Tang, Y.; Cui, J. Behavioral and neurogenomic responses to acoustic and visual sexual cues are correlated in female torrent frogs. Asian Herpetol. Res. 2021, 12, 88–99. [Google Scholar]
- Merilaita, S.; Tuomi, J.; Jormalainen, V. Optimization of cryptic coloration in heterogeneous habitats. Biol. J. Linn. Soc. 1999, 67, 151–161. [Google Scholar] [CrossRef]
- Leclercq, E.; Taylor, J.; Migaud, H. Morphological skin colour changes in teleosts. Fish Fish. 2010, 11, 159–193. [Google Scholar] [CrossRef]
- Sköld, H.N.; Aspengren, S.; Wallin, M. Rapid color change in fish and amphibians-function, regulation, and emerging applications. Pigm. Cell. Melanoma. R. 2013, 26, 29–38. [Google Scholar] [CrossRef]
- Stevens, M. Color change, phenotypic plasticity, and camouflage. Front. Ecol. Evol. 2016, 4, 51. [Google Scholar]

{kind=link}
| Species | Sample Size | Number Escaped/Not Escaped | Number Silent/Not Silent |
|---|---|---|---|
| Little Torrent Frog | 13 | 1/12 | 0/13 |
| Ornamented Pygmy Frog | 15 | 0/15 | 0/15 |
| Spot-legged Treefrog | 13 | 0/13 | 0/13 |
| Species | Call Type | Number Not Escaped | Number Escaped | p |
|---|---|---|---|---|
| Little Torrent Frog | TS | 30 | 1 | <0.001 |
| SCEC | 31 | 0 | <0.001 | |
| SEC | 31 | 0 | <0.001 | |
| SHEC | 31 | 0 | <0.001 | |
| Ornamented Pygmy Frog | TS | 20 | 1 | <0.001 |
| SCEC | 21 | 0 | <0.001 | |
| SEC | 21 | 0 | <0.001 | |
| SHEC | 21 | 0 | <0.001 | |
| Spot-legged Treefrog | TS | 24 | 0 | <0.001 |
| SCEC | 23 | 0 | <0.001 | |
| SEC | 23 | 1 | <0.001 | |
| SHEC | 23 | 1 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Qin, Y.; Wang, J.; Liang, W. Avian Alarm Calls Do Not Induce Anti-Predator Response in Three Anuran Species. Animals 2022, 12, 3537. https://doi.org/10.3390/ani12243537
Zhao L, Qin Y, Wang J, Liang W. Avian Alarm Calls Do Not Induce Anti-Predator Response in Three Anuran Species. Animals. 2022; 12(24):3537. https://doi.org/10.3390/ani12243537
Chicago/Turabian StyleZhao, Longhui, Yuanyu Qin, Jichao Wang, and Wei Liang. 2022. "Avian Alarm Calls Do Not Induce Anti-Predator Response in Three Anuran Species" Animals 12, no. 24: 3537. https://doi.org/10.3390/ani12243537
APA StyleZhao, L., Qin, Y., Wang, J., & Liang, W. (2022). Avian Alarm Calls Do Not Induce Anti-Predator Response in Three Anuran Species. Animals, 12(24), 3537. https://doi.org/10.3390/ani12243537