
Molecular Characterization, Tissue Distribution and Differential Nutritional Regulation of Three n-3 LC-PUFA Biosynthesis-Related Genes in Hybrid Grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Sampling
2.2. Gene Cloning
2.2.1. Primers Design
2.2.2. RNA Extraction and cDNA Synthesis
2.2.3. RACE Amplification
2.2.4. Sequence and Phylogenetic Analysis
2.3. Quantitative Real-Time PCR
2.4. Statistical Analysis
3. Results
3.1. Gene Cloning and Sequence Analysis of elovl5, elovl8 and fads2
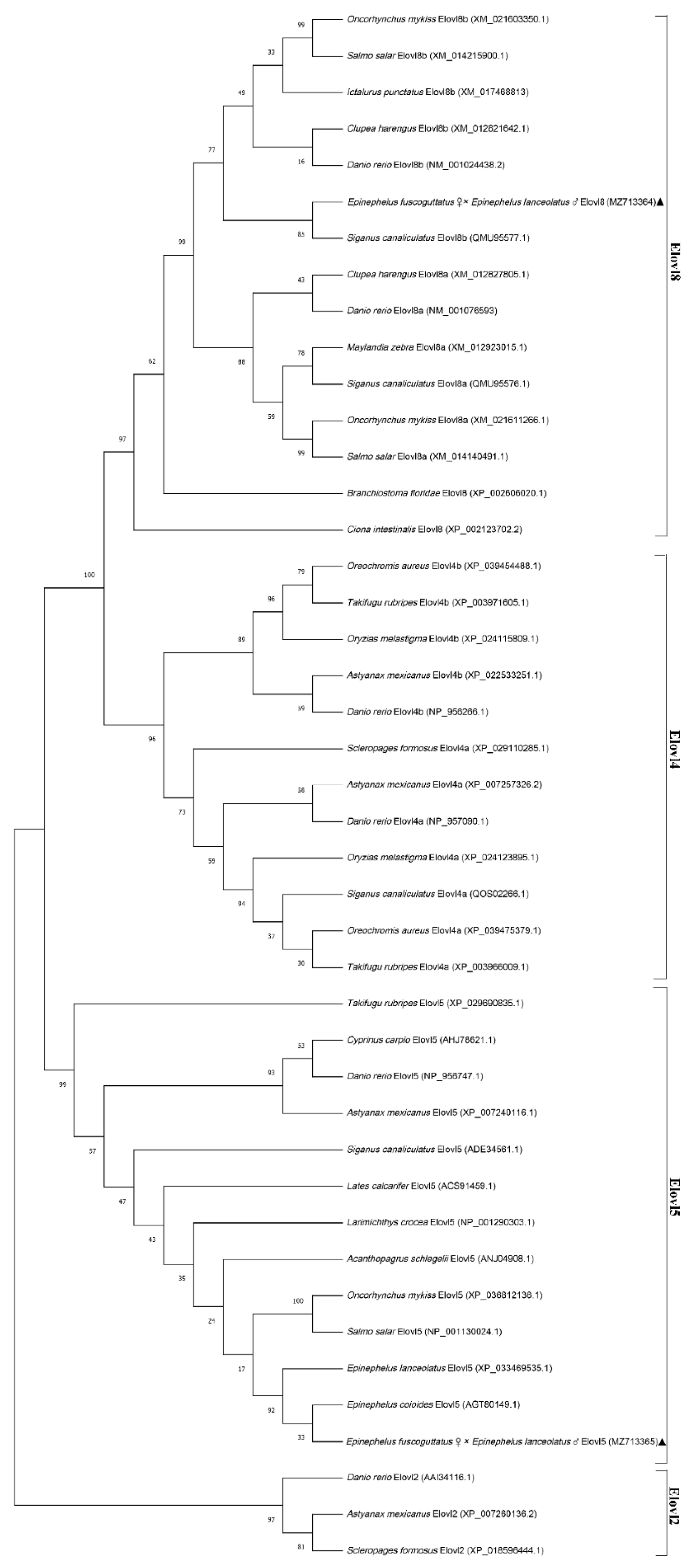
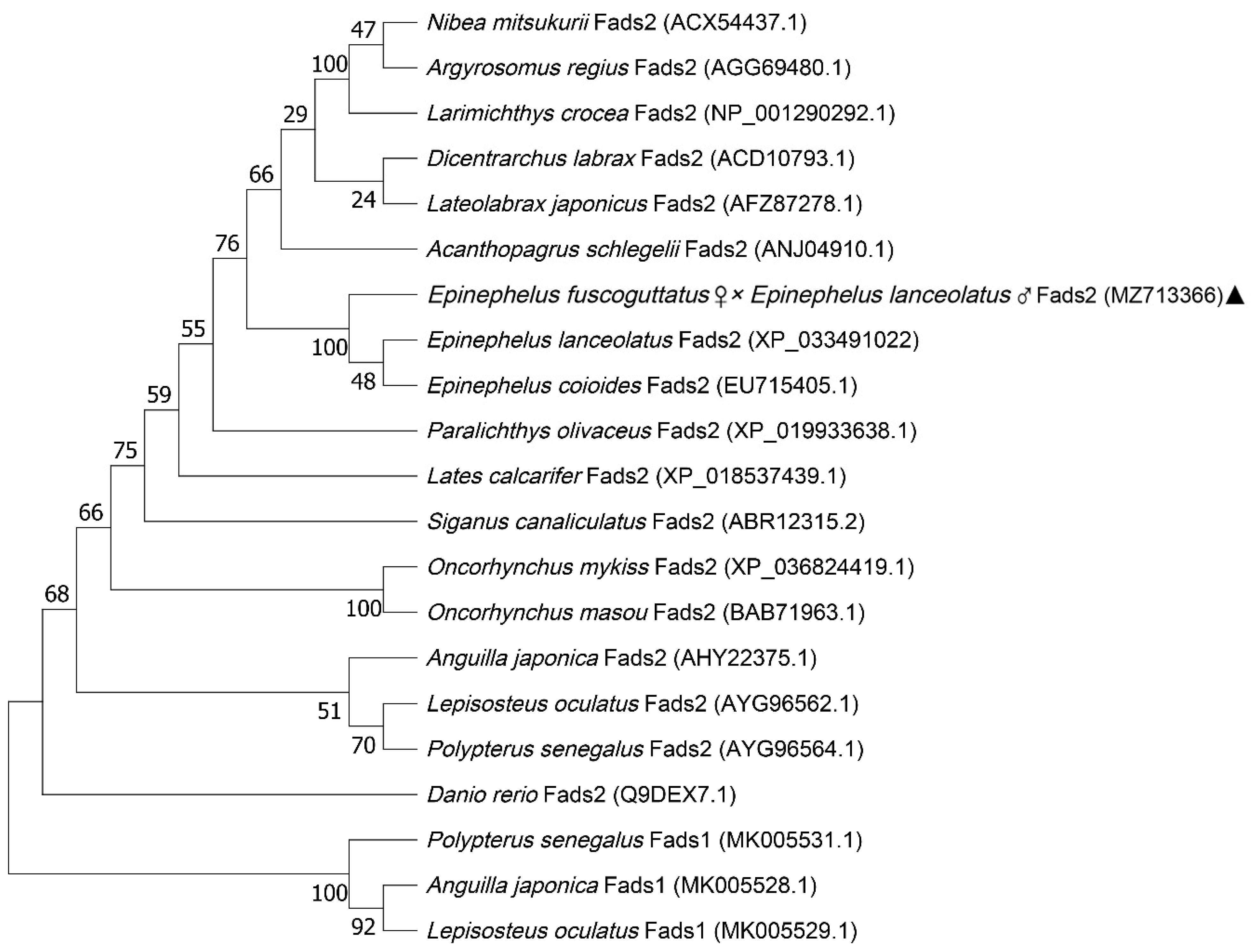
3.2. Multiple Sequence Alignment and Phylogenetic Analysis
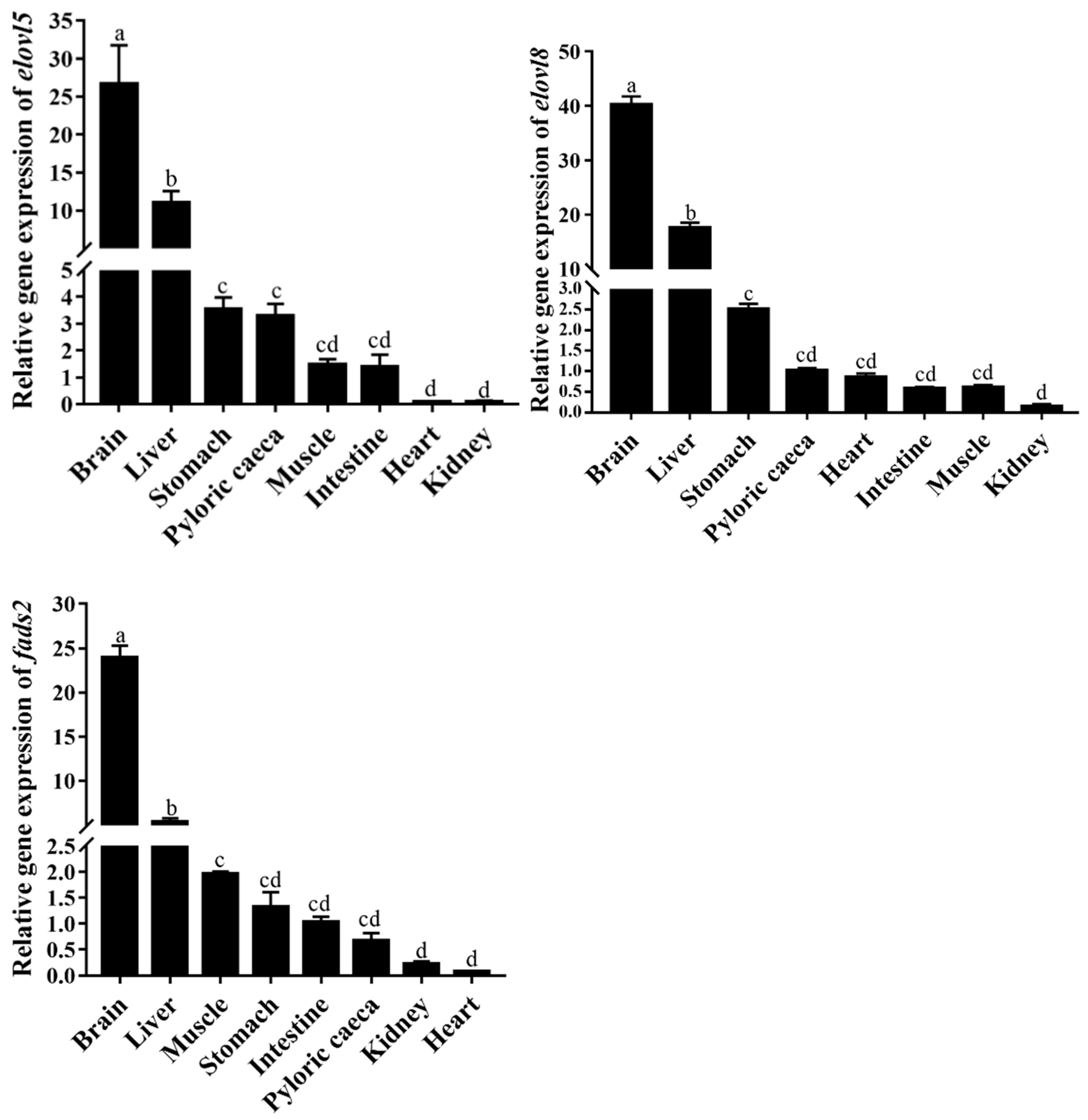
3.3. Tissue Distribution of elovl5, elovl8 and fads2 mRNA Hybrid Grouper
3.4. Nutritional Regulation of Hybrid Grouper elovl5, elovl8 and fads2 Genes Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, T.T.; Xu, J.; Wang, Y.M.; Xue, C.H. Health benefits of dietary marine DHA/EPA-enriched glycerophospholipids. Prog. Lipid Res. 2019, 75, 100997. [Google Scholar] [CrossRef] [PubMed]
- Zárate, R.; Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.; Rodríguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abedi, E.; Sahari, M.A. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef] [PubMed]
- Auchterlonie, N. The continuing importance of fishmeal and fish oil in aquafeeds. Presented at the Aquafarm Conference, Pordenone, Italy, 15–16 February 2018. [Google Scholar]
- Bell, J.G.; Tocher, D.R.; Henderson, R.J.; Dick, J.R.; Crampton, V.O. Altered fatty acid compositions in Atlantic salmon (Salmo salar) fed diets containing linseed and rapeseed oils can be partially restored by a subsequent fish oil finishing diet. J. Nutr. 2003, 133, 2793–2801. [Google Scholar] [CrossRef] [PubMed]
- Menoyo, D.; López-Bote, C.J.; Obach, A.; Bautista, J.M. Effect of dietary fish oil substitution with linseed oil on the performance, tissue fatty acid profile, metabolism, and oxidative stability of Atlantic salmon. J. Anim. Sci. 2005, 83, 2853–2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monroig, Ó.; Kabeya, N. Desaturases and elongases involved in polyunsaturated fatty acid biosynthesis in aquatic invertebrates: A comprehensive review. Fish. Sci. 2018, 84, 911–928. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhang, M.; Li, Y.; Wang, S.; Xie, D.; Wen, X.; Monroig, Ó. Identification of miR-145 as a Key Regulator Involved in LC-PUFA Biosynthesis by Targeting hnf4 α in the Marine Teleost Siganus canaliculatus. J. Agric. Food Chem. 2020, 68, 15123–15133. [Google Scholar] [CrossRef]
- Sun, S.; Wang, Y.; Goh, P.T.; Lopes-Marques, M.; Castro, L.F.C.; Monroig, Ó.; Gao, J. Evolution and Functional Characteristics of the Novel elovl8 That Play Pivotal Roles in Fatty Acid Biosynthesis. Genes 2021, 12, 1287. [Google Scholar] [CrossRef]
- Monroig, O.; Rotllant, J.; Sanchez, E.; Cerda-Reverter, J.M.; Tocher, D.R. Expression of long-chain polyunsaturated fatty acid (LC-PUFA) biosynthesis genes during zebrafish Danio rerio early embryogenesis. BBA Mol. Cell Biol. Lipids 2009, 1791, 1093–1101. [Google Scholar] [CrossRef] [Green Version]
- Morais, S.; Monroig, O.; Zheng, X.; Leaver, M.; Tocher, D. Highly unsaturated fatty acid synthesis in Atlantic Salmon: Characterization of ELOVL5- and ELOVL2-like elongases. Mar. Biotechnol. 2009, 11, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Gregory, M.K.; James, M.J. Rainbow trout (Oncorhynchus mykiss) Elovl5 and Elovl2 differ in selectivity for elongation of omega-3 docosapentaenoic acid. BBA Mol. Cell Biol. Lipids 2014, 1841, 1656–1660. [Google Scholar] [CrossRef] [PubMed]
- Oboh, A.; Betancor, M.B.; Tocher, D.R.; Monroig, O. Biosynthesis of longchain polyunsaturated fatty acids in the African catfish Clarias gariepinus: Molecular cloning and functional characterisation of fatty acyl desaturase (fads2) and elongase (elovl2) cDNAs. Aquaculture 2016, 462, 70–79. [Google Scholar] [CrossRef]
- Monroig, O.; Lopes-Marques, M.; Navarro, J.C. Evolutionary functional elaboration of the Elovl2/5 gene family in chordates. Sci. Rep. 2016, 6, 20510. [Google Scholar] [CrossRef]
- Monroig, Ó.; Navarro, J.C.; Tocher, D.R. Long-chain polyunsaturated fatty acids in fish: Recent advances on desaturases and elongases involved in their biosynthesis. In Proceedings of the Avances en Nutrición Acuícola XI—Memorias del Décimo Primer Simposio Internacional de Nutrición Acuícola, San Nicolás de los Garza, Mexico, 23–25 November 2011; pp. 257–283. [Google Scholar]
- Castro, L.F.C.; Tocher, D.R.; Monroig, O. Long-chain polyunsaturated fatty acid biosynthesis in chordates: Insights into the evolution of Fads and Elovl gene repertoire. Prog. Lipid Res. 2016, 62, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.E.; Kelder, B.; Bobik, E.G.; Chuang, L.T.; Lewis, C.J.; Kopchick, J.J.; Mukerji, P.; Huang, Y.S. Identification and expression of mammalian long-chain PUFA elongation enzymes. Lipids 2002, 37, 733–740. [Google Scholar] [CrossRef]
- Monroig, O.; Wang, S.; Zhang, L.; You, C.; Tocher, D.R.; Li, Y. Elongation of long-chain fatty acids in rabbitfish Siganus canaliculatus: Cloning, functional characterisation and tissue distribution of Elovl5-and Elovl4-like elongases. Aquaculture 2012, 350, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wen, Z.; You, C.; Xie, Z.; Li, Y. Genome wide identification and functional characterization of two LC-PUFA biosynthesis elongase (elovl8) genes in rabbitfish (Siganus canaliculatus). Aquaculture 2020, 522, 735127. [Google Scholar]
- Sprecher, H.; Luthria, D.L.; Mohammed, B.S.; Baykousheva, S.P. Reevaluation of the pathways for the biosynthesis of polyunsaturated fatty acids. J. Lipid Res. 1995, 36, 2471–2477. [Google Scholar] [CrossRef]
- Marquardt, A.; Stöhr, H.; White, K.; Weber, B.H. cDNA cloning, genomic structure, and chromosomal localization of three members of the human fatty acid desaturase family. Genomics 2000, 66, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Kabeya, N.; Yamamoto, Y.; Cummins, S.F.; Elizur, A.; Yazawa, R.; Takeuchi, Y.; Yoshizaki, G. Polyunsaturated fatty acid metabolism in a marine teleost, Nibe croaker Nibea mitsukurii: Functional characterization of Fads2 desaturase and Elovl5 and Elovl4 elongases. Comp. Biochem. Physiol. Part B Biochem. 2015, 188, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Abdul Hamid, N.K.; Carmona-Antonanzas, G.; Monroig, O.; Tocher, D.R.; Turchini, G.M.; Donald, J.A. Isolation and functional characterisation of a fads2 in rainbow trout (Oncorhynchus mykiss) with Δ5 desaturase activity. PLoS ONE 2016, 11, 0150770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Monroig, O.; Zhang, L.; Wang, S.; Zheng, X.; Dick, J.R. Vertebrate fatty acyl desaturase with Δ4 activity. Proc. Natl. Acad. Sci. USA 2010, 107, 16840–16845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morais, S.; Castanheira, F.; Martínez-Rubio, L.; Conceição, L.E.C.; Tocher, D.R. Long-chain polyunsaturated fatty acid synthesis in a marine vertebrate: Ontogenetic and nutritional regulation of a fatty acyl desaturase with Δ4 activity. Biochim. Biophys. Acta 2012, 1821, 660–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yearbook, C.F.S. China Fishery Statistics Yearbook; Bureau of Fisheries, Ministry of Agriculture: Beijing, China, 2021. [Google Scholar]
- Sun, Y.; Guo, C.Y.; Wang, D.D.; Li, X.F.; Xiao, L.; Zhang, X.; Zhang, Y. Transcriptome analysis reveals the molecular mechanisms underlying growth superiority in a novel grouper hybrid (Epinephelus fuscogutatus♀ × E. lanceolatus♂). BMC Genet. 2016, 17, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Mai, K.; Xu, W.; Yuan, Y.; Zhang, Y.; Ai, Q. Characterization, mRNA expression and regulation of Δ6 fatty acyl desaturase (FADS2) by dietary n-3 long chain polyunsaturated fatty acid (LC-PUFA) levels in grouper larvae (Epinephelus coioides). Aquaculture 2014, 434, 212–219. [Google Scholar] [CrossRef]
- Li, S.; Monroig, Ó.; Navarro, J.C.; Yuan, Y.; Xu, W.; Mai, K.; Ai, Q. Molecular cloning and functional characterization of a putative Elovl4 gene and its expression in response to dietary fatty acid profiles in orange-spotted grouper Epinephelus coioides. Aquacult. Res. 2017, 48, 537–552. [Google Scholar] [CrossRef]
- Baoshan, L.; Jiying, W.; Yu, H.; Tiantian, H.; Shixin, W.; BingShan, H.; Yongzhi, S. Effects of replacing fish oil with wheat germ oil on growth, fat deposition, serum biochemical indices and lipid metabolic enzyme of juvenile hybrid grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂). Aquaculture 2019, 505, 54–62. [Google Scholar] [CrossRef]
- Yong, A.S.K.; Mubarak, N.S.S.; Shapawi, R. Effects of partial replacement of fish oil with different vegetable oils on growth, feed utilization and fatty acid profile of hybrid grouper juvenile (Epinephelus fuscoguttatus x Epinephelus lanceolatus). J. Oil Palm Res. 2019, 31, 110–121. [Google Scholar]
- An, W.; Dong, X.; Tan, B.; Wu, M.; Zhang, S.; Chi, S.; Yang, Y. Effects of dietary vegetable oil on growth performance, digestive capacity, antioxidant capacity and expression of immune-related genes in the hybrid grouper (Epinephelus fuscoguttatus♀ × E. lanceolatus♂). Aquacult. Nutr. 2020, 26, 2086–2101. [Google Scholar] [CrossRef]
- Yan, X.; Dong, X.; Tan, B.; Zhang, S.; Chi, S.; Liu, H.; Yang, Y. Effects of alternative dietary oils on lipid metabolism and related gene expression in hybrid grouper (Epinephelus fuscoguttatus♀ × ♂ E. lanceolatu). Aquacult. Nutr. 2021, 27, 105–115. [Google Scholar] [CrossRef]
- Xie, D.; Chen, C.; Dong, Y.; You, C.; Wang, S.; Monroig, Ó.; Li, Y. Regulation of long-chain polyunsaturated fatty acid biosynthesis in teleost fish. Prog. Lipid Res. 2021, 36, 2471–2477. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.; Kirsch, H.; Domergue, F.; Abbadi, A.; Sperling, P.; Bauer, J.; Cirpus, P.; Zank, T.K.; Moreau, H.; Roscoe, T.J. Novel fatty acid elongases and their use for the reconstitution of docosahexaenoic acid biosynthesis. J. Lipid Res. 2004, 45, 1899–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsson, A.; Westerberg, R.; Jacobsson, A. Fatty acid elongases in mammals: Their regulation and roles in metabolism. Prog. Lipid Res. 2006, 45, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Tocher, D.R.; Zheng, X.; Schlechtriem, C.; Hastings, N.; Dick, J.R.; Teale, A.J. Highly unsaturated fatty acid synthesis in marine fish: Cloning, functional characterization, and nutritional regulation of fatty acyl Δ6 desaturase of Atlantic cod (Gadus morhua L.). Lipids 2006, 41, 1003–1016. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Marques, M.; Kabeya, N.; Qian, Y.; Ruivo, R.; Santos, M.M.; Venkatesh, B.; Tocher, D.R.; Castro, L.F.C.; Monroig, Ó. Retention of fatty acyl desaturase 1 (fads1) in Elopomorpha and Cyclostomata provides novel insights into the evolution of long-chain polyunsaturated fatty acid biosynthesis in vertebrates. BMC. Evol. Biol. 2018, 18, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, D.; Fu, Z.; Wang, S.; You, C.; Monroig, O.; Tocher, D.R.; Li, Y. Characteristics of the fads2 gene promoter in marine teleost Epinephelus coioides and role of Sp1-binding site in determining promoter activity. Sci. Rep. 2018, 8, 5305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; King, Z.; Xu, Y.; Monroig, O.; Morais, S.; Tocher, D.R. Physiological roles of fatty acyl desaturases and elongases in marine fish: Characterisation of cDNAs of fatty acyl Δ6desaturase and Elovl5 elongase of cobia (Rachycentron canadum). Aquaculture 2009, 290, 122–131. [Google Scholar] [CrossRef]
- Mohd-Yusof, N.Y.; Monroig, Ó.; Mohd-Adnan, A.; Wan, K.-L.; Tocher, D.R. Investigation of highly unsaturated fatty acid metabolism in the Asian sea bass, Lates calcarifer. Fish Physiol. Biochem. 2010, 3, 827–843. [Google Scholar] [CrossRef]
- Carmona-Antonanzas, G.; Tocher, D.R.; Taggart, J.B.; Leaver, M.J. An evolutionary perspective on Elovl5 fatty acid elongase: Comparison of Northern pike and duplicated paralogs from Atlantic salmon. BMC Evol. Biol. 2013, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.V.; Dick, J.R.; Porter, A.E. Pyloric ceca are significant site of of newly synthesized 22:6n3 in rainbow trout (Oncorhynchus mykiss). Lipids 2003, 39, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yuan, Y.; Wang, T.; Xu, W.; Li, M.; Mai, K.; Ai, Q. Molecular cloning, functional characterization and nutritional regulation of the putative elongase Elovl5 in the orange-spotted grouper (Epinephelus coioides). PLoS ONE 2016, 11, 0150544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, D.; Ye, J.; Lu, M.; Wang, S.; You, C.; Li, Y. Comparsion of activities of fatty acyl desaturases and elongases among six teleosts with different feeding and ecological habits. Front. Mar. Sci. 2020, 7, 117. [Google Scholar] [CrossRef] [Green Version]
- Zuo, R.; Mai, K.; Xu, W.; Dong, X.; Ai, Q. Molecular cloning, tissue distribution and nutritional regulation of a fatty acyl elovl5-like elongase in large yellow croaker, Larimichthys crocea. Aquacult. Res. 2016, 47, 2393–2406. [Google Scholar] [CrossRef]
- Minghetti, M.; Leaver, M.J.; Tocher, D.R. Transcriptional control mechanisms of genes of lipid and fatty acid metabolism in the Atlantic salmon (Salmo salar l.) established cell line, shk-1. Biochim. Biophys. Acta 2011, 1811, 194–202. [Google Scholar] [CrossRef]
- Geay, F.; Zambonino-Infante, J.; Reinhardt, R.; Kuhl, H.; Santigosa, E.; Cahu, C. Characteristics of Fads2 gene expression and putative promoter in European sea bass (Dicentrarchus labrax): Comparison with salmonid species and analysis of CpG methylation. Mar. Genom. 2012, 5, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | Usage | TM | Expected Amplicon Size |
|---|---|---|---|---|
| Elovl5 F | ACCTTCTAATCGTGTGGATG | Partial sequence | 51.9 °C | 648 bp |
| Elovl5 R | TGTGCTTCTTGTAAGTCTGA | Partial sequence | 51.3 °C | |
| Elovl5 5F | AAGCAGTGGTATCAACGCAGAGT | 5’-RACE | 59 °C | 383 bp |
| Elovl5 5R | TCCACTTCCTGTGCACTGTGAGTGTC | 5’-RACE | 62.8 °C | |
| Elovl5 3F | CACACTCATCTTCCTCTTCTCAAACTT | 3’-RACE | 57.3 °C | 466 bp |
| Elovl5 3R1 | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | 3’-RACE | 68 °C | |
| Elovl5 3R2 | CTAATACGACTCACTATAGGGC | 3’-RACE | 51.8 °C | |
| Elovl8 F | ATGGCTGCTGGTCTACTC | Partial sequence | 53.3 °C | 677 bp |
| Elovl8 R | AGTTACTGAAGAGGATGATGAG | Partial sequence | 51.4 °C | |
| Elovl8 5F | AAGCAGTGGTATCAACGCAGAGT | 5’-RACE | 59 °C | 608 bp |
| Elovl8 5R | GGTGCCGTGGTGGTAAACATGAAGG | 5’-RACE | 62.5 °C | |
| Elovl8 3F | ATGCAGAAGTACCTGTGGTGGAAGAGA | 3’-RACE | 61.4 °C | 538 bp |
| Elovl8 3R1 | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | 3’-RACE | 68 °C | |
| Elovl8 3R2 | CTAATACGACTCACTATAGGGC | 3’-RACE | 51.8 °C | |
| Fads2 F | CACTACGCTGGAGAGGATG | Partial sequence | 55.2 °C | 608 bp |
| Fads2 R | TGGTGTTGGTGATGATAGG | Partial sequence | 51.4 °C | |
| Fads2 5F | AAGCAGTGGTATCAACGCAGAGT | 5’-RACE | 59 °C | 476 bp |
| Fads2 5R | CTCCGTGGCATCCTCTCCAGCGTAGT | 5’-RACE | 65.6 °C | |
| Fads2 3F | CATTTCCAGCATCACGCTAAACCCAAC | 3’-RACE | 60 °C | 1117 bp |
| Fads2 3R1 | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | 3’-RACE | 68 °C | |
| Fads2 3R2 | CTAATACGACTCACTATAGGGC | 3’-RACE | 51.8 °C | |
| elongation factor 1 alpha F | AGGGATGGAAGATTGAGCGC | Internal control | 57.2 °C | 74 bp |
| elongation factor 1 alphaR | CGTACCGGGCTTCAGGATAC | Internal control | 57 °C | |
| Elovl8 RT-F | CAGATGATCCAGTTCCACGTCA | qRT-PCR | 56.8 °C | 525 bp |
| Elovl8 RT-R | GCGGTAGGTCTGGTAGTAGAAG | qRT-PCR | 56.1 °C | |
| Elovl5 RT-F | CTACTGCCAGGACACTCACA | qRT-PCR | 56 °C | 415 bp |
| Elovl5 RT-R | GAGGCGCCAAAGTATGAGTG | qRT-PCR | 56 °C | |
| Fads2 RT-F | CCAGGTGGAGGCAGAAGAACA | qRT-PCR | 58.8 °C | 184 bp |
| Fads2 RT-R | AGCCACTATGCTGGAGAGGATG | qRT-PCR | 58.2 °C | |
| Elovl8 AF | AGCGTGCTCACTCACTTTTAACGG | ORF validation | 57 °C | 933 bp |
| Elovl8 AR | GTGTGCTTCTGCCTTCTCCATCCTT | ORF validation | 59 °C | |
| Elovl5 AF | GTCGCTTTCTCTCCCCCGCCTCTCA | ORF validation | 64 °C | 953 bp |
| Elovl5 AR | CTACAGTGAGAATTGGGTGGCGGTTT | ORF validation | 60 °C | |
| Fads2 AF | GCCAAAATCTGGATACTGTGTCAAA | ORF validation | 54 °C | 1628 bp |
| Fads2 AR | AAGACACTGTAAGGCAACCAGAGAAA | ORF validation | 56 °C |
| Ingredients (g/100 g Diet) | Dietary n-3 PUFA Levels | ||
|---|---|---|---|
| 0.53 | 1.19 | 2.69 | |
| Fish meal 1 | 22 | 22 | 22 |
| Pork blood meal 2 | 6.0 | 6.0 | 6.0 |
| Chicken meal 3 | 18 | 18 | 18 |
| Casein 4 | 17.5 | 17.5 | 17.5 |
| Palm oil 5 | 3.41 | 2.53 | 0.66 |
| DHA purified oil 6 | 0.2 | 0.73 | 1.87 |
| EPA purified oil 7 | 0 | 0.35 | 1.08 |
| Vitamin premix 8 | 1 | 1 | 1 |
| Mineral Premix 9 | 0.5 | 0.5 | 0.5 |
| Starch | 16 | 16 | 16 |
| Cellulose | 13.4 | 13.4 | 13.4 |
| Crude protein | 53.3 | 53.4 | 53.6 |
| Crude lipid | 7.12 | 6.93 | 7.1 |
| n-3 PUFA | 0.53 | 1.19 | 2.69 |
| Fatty Acid | Dietary n-3 PUFA Levels | ||
|---|---|---|---|
| 0.53 | 1.19 | 2.69 | |
| 14:0 | 1.01 | 1.27 | 2.04 |
| 16:0 | 28.18 | 23.55 | 10.16 |
| 18:0 | 10.26 | 8.75 | 9.40 |
| 16:1n-9 | 0.97 | 2.12 | 3.18 |
| 18:1n-7 | 0.80 | 0.87 | 0.91 |
| 18:1n-9 | 20.75 | 17.33 | 19.48 |
| 20:1n-9 | 0.11 | 0.11 | 0.21 |
| 22:1n-9 | 0.11 | 0.10 | 0.10 |
| 18:2n-6 (LA) | 18.13 | 13.06 | 5.99 |
| 18:3n-6 | 0.41 | 0.32 | 0.30 |
| 20:2n-6 | 0.10 | 0.20 | 0.31 |
| 20:4n-6 (ARA) | 1.06 | 1.07 | 0.99 |
| 18:3n-3 (ALA) | 1.07 | 1.00 | 1.01 |
| 20:5n-3 (EPA) | 2.99 | 6.69 | 15.12 |
| 22:6n-3 (DHA) | 4.61 | 10.28 | 23.29 |
| ∑SFA | 39.45 | 33.57 | 21.60 |
| ∑MUFA | 22.73 | 20.54 | 23.87 |
| ∑n-3 PUFA | 7.60 | 16.97 | 38.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Zheng, Z.; Wang, C.; Wang, Y.; Sun, Y.; Gao, Y. Molecular Characterization, Tissue Distribution and Differential Nutritional Regulation of Three n-3 LC-PUFA Biosynthesis-Related Genes in Hybrid Grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Animals 2022, 12, 234. https://doi.org/10.3390/ani12030234
Wu Q, Zheng Z, Wang C, Wang Y, Sun Y, Gao Y. Molecular Characterization, Tissue Distribution and Differential Nutritional Regulation of Three n-3 LC-PUFA Biosynthesis-Related Genes in Hybrid Grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Animals. 2022; 12(3):234. https://doi.org/10.3390/ani12030234
Chicago/Turabian StyleWu, Qingjun, Zhi Zheng, Chuijin Wang, Yao Wang, Yuejia Sun, and Yujie Gao. 2022. "Molecular Characterization, Tissue Distribution and Differential Nutritional Regulation of Three n-3 LC-PUFA Biosynthesis-Related Genes in Hybrid Grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂)" Animals 12, no. 3: 234. https://doi.org/10.3390/ani12030234
APA StyleWu, Q., Zheng, Z., Wang, C., Wang, Y., Sun, Y., & Gao, Y. (2022). Molecular Characterization, Tissue Distribution and Differential Nutritional Regulation of Three n-3 LC-PUFA Biosynthesis-Related Genes in Hybrid Grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Animals, 12(3), 234. https://doi.org/10.3390/ani12030234