
Genetic Evaluation of Body Weights and Egg Production Traits Using a Multi-Trait Animal Model and Selection Index in Thai Native Synthetic Chickens (Kaimook e-san2)

 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
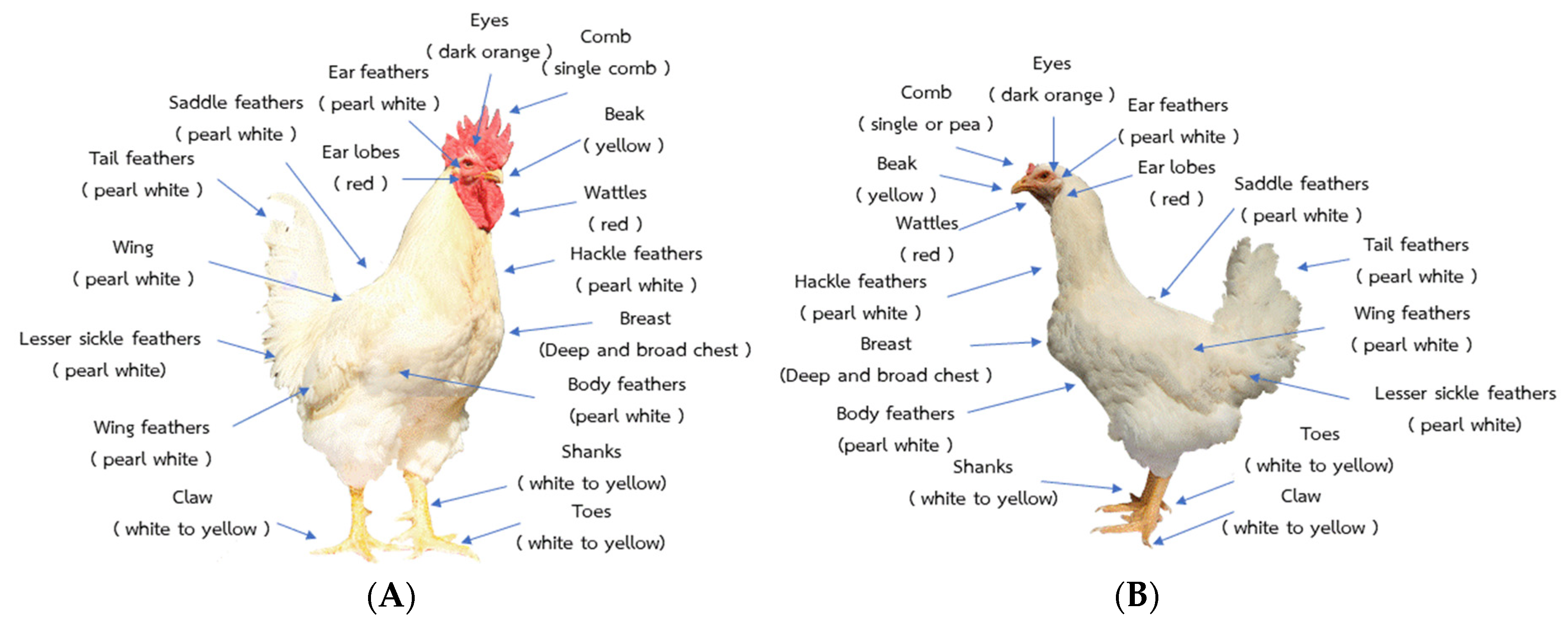
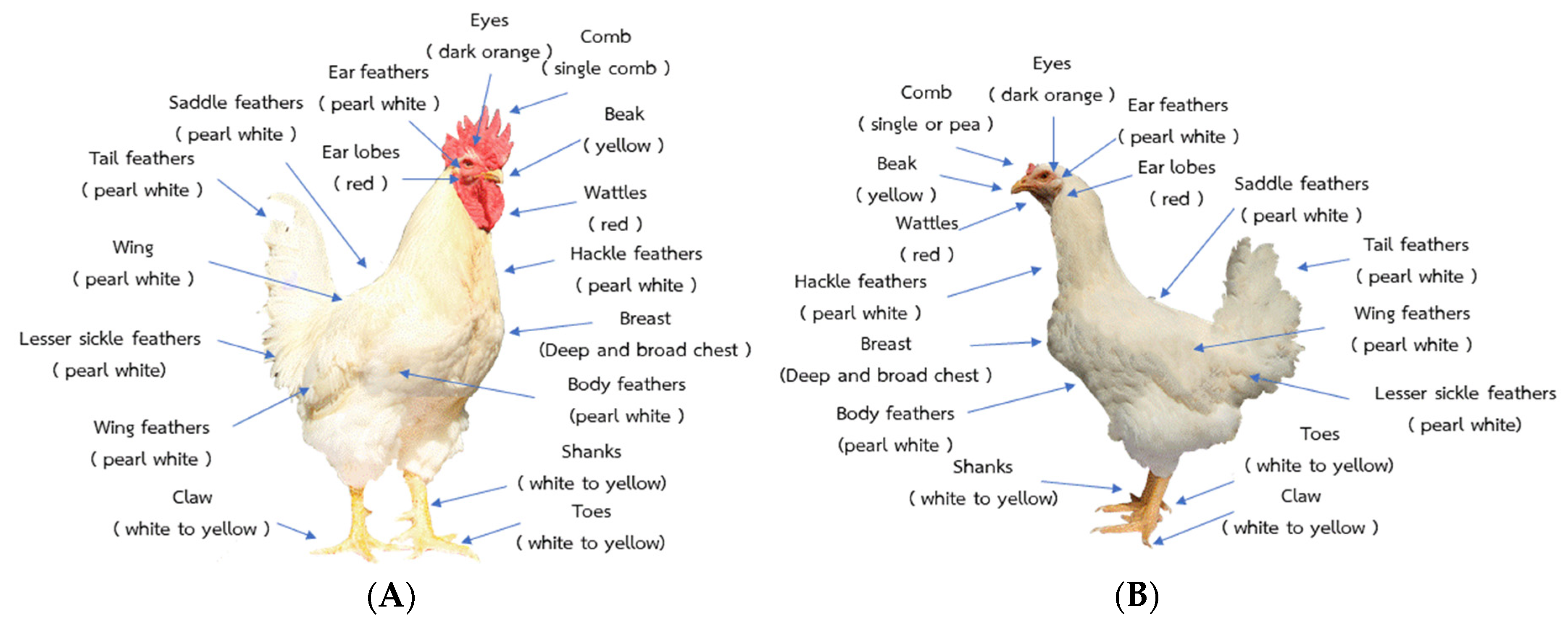
2.1. Animals, Breeding Plan, and Morphology
2.2. Parent Stock Management
2.3. Data Collection
2.4. Genetic Model and Statistical Analysis
2.5. Genetic Progress and Selection Index
3. Results
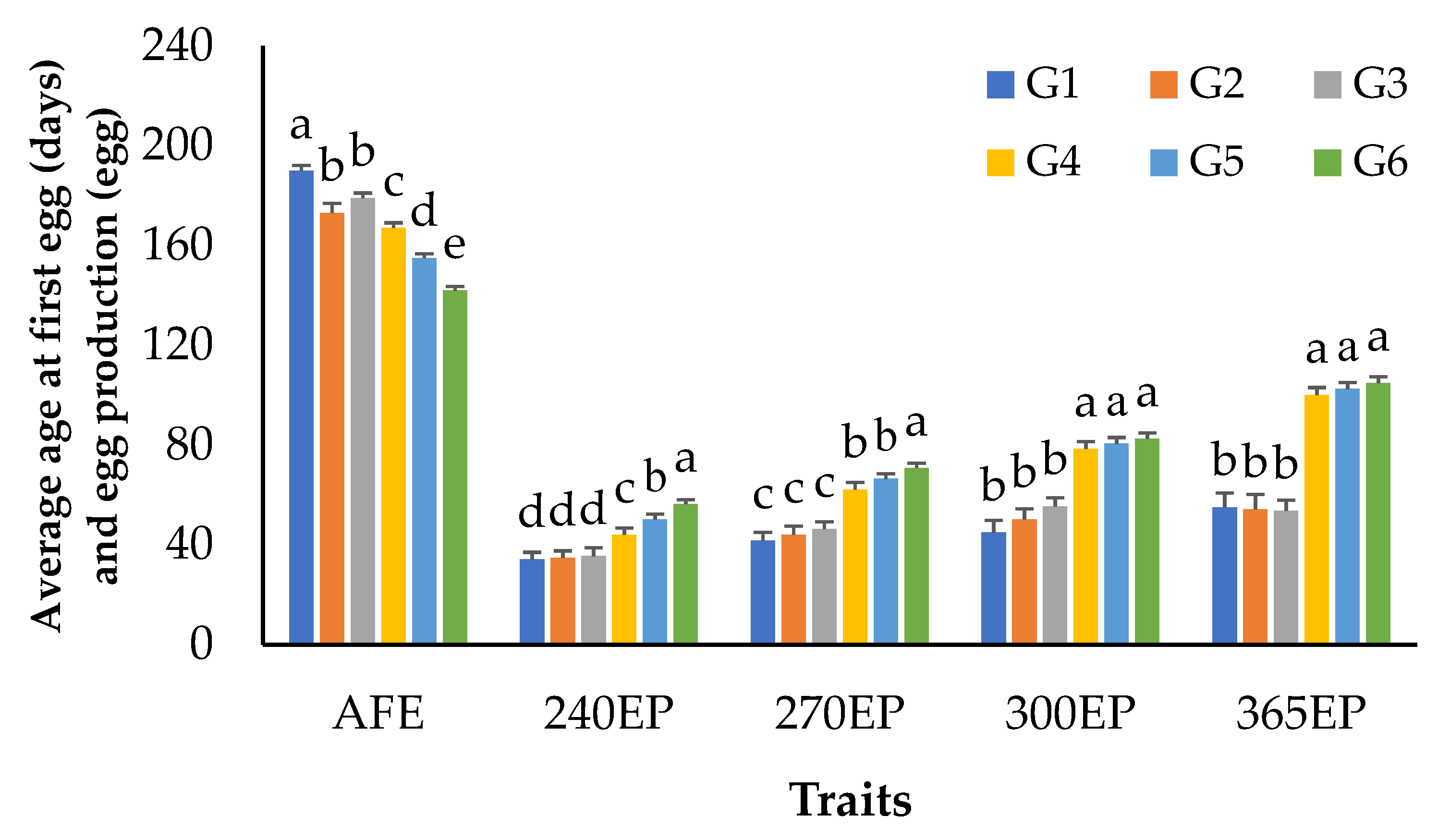
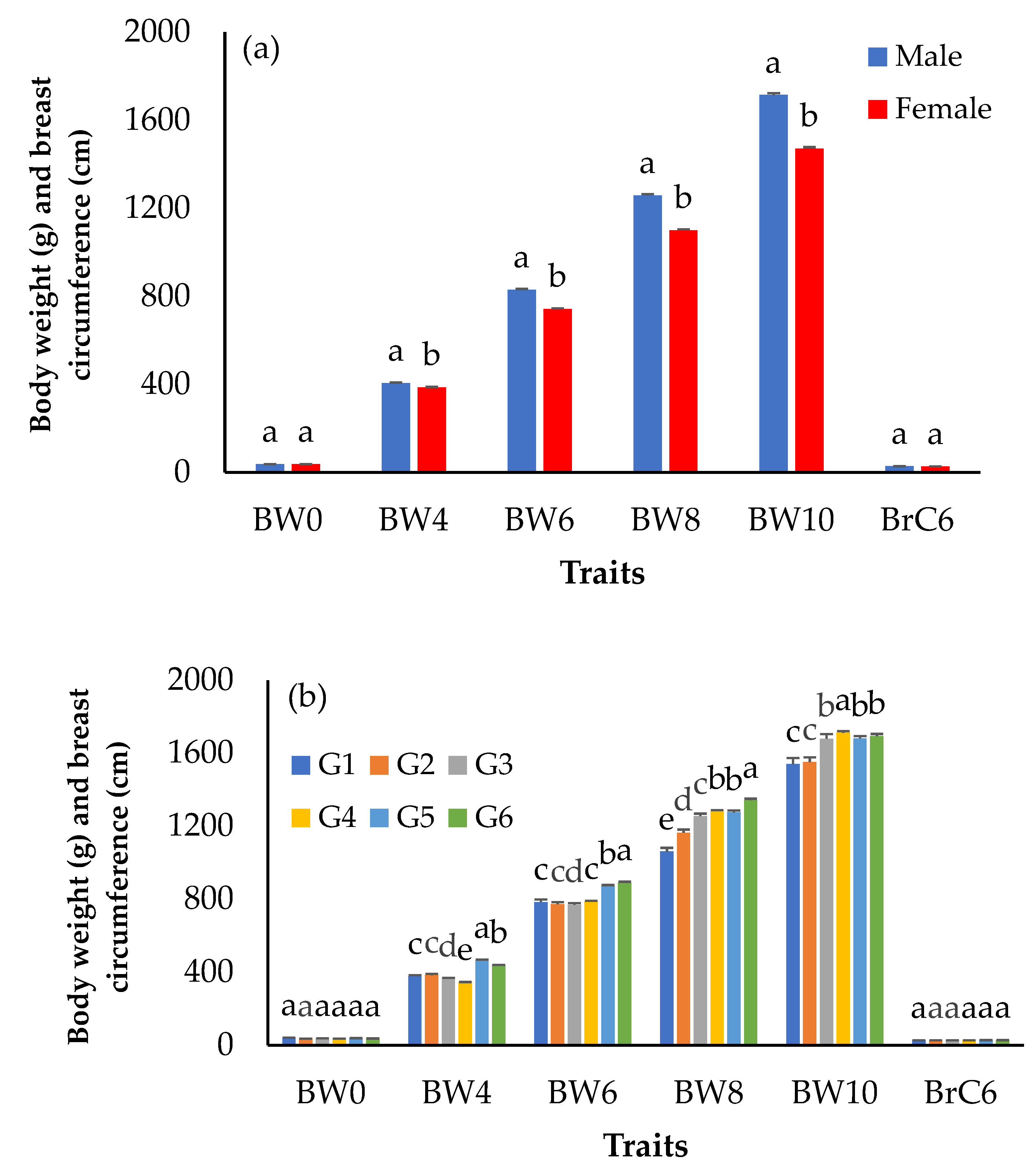
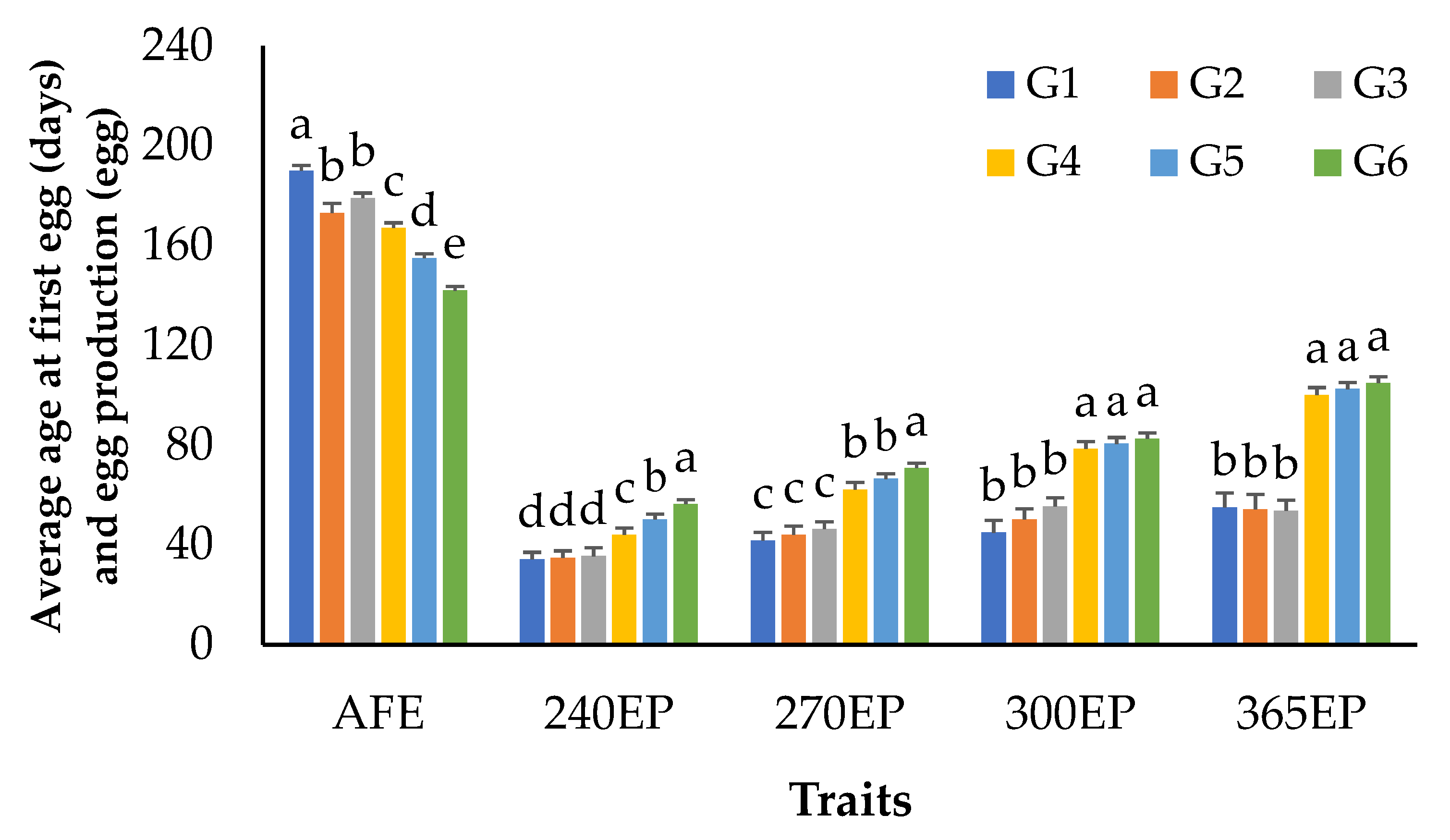
3.1. Growth and Egg Production Performance
3.2. Estimated Variance Components and Heritability
3.3. Genetic and Phenotypic Correlation Estimates
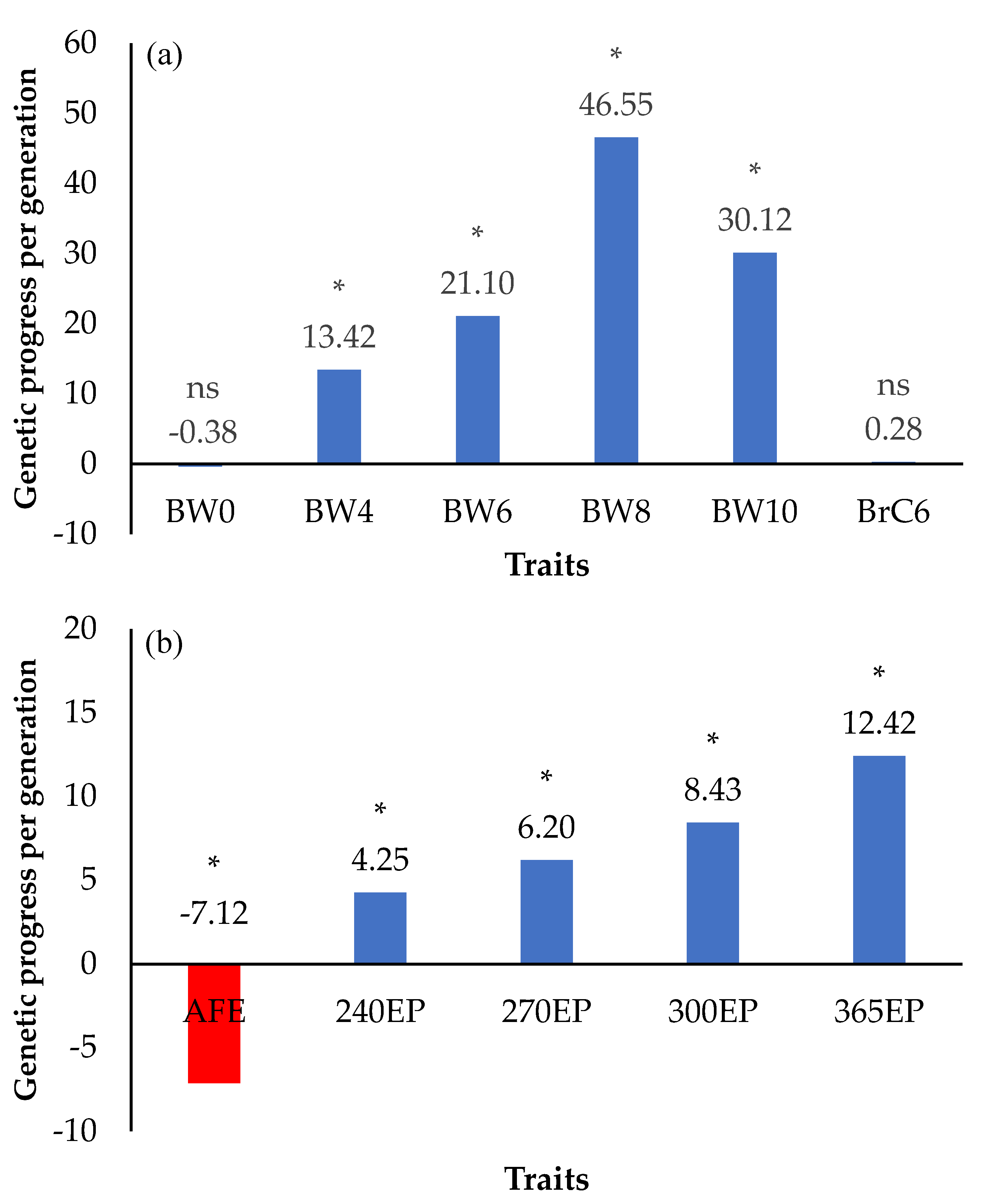
3.4. Genetic Progress (ΔG/Generation)
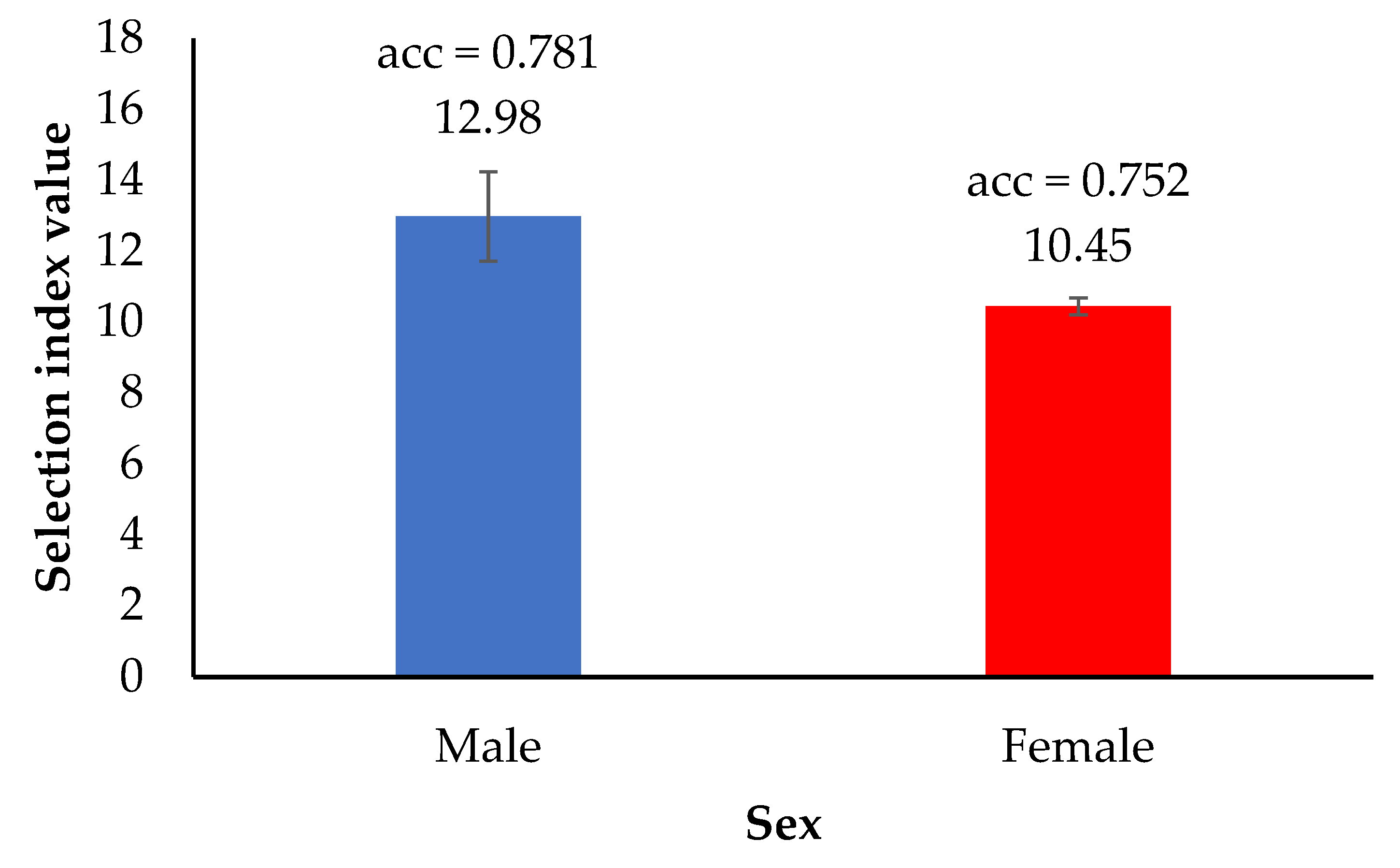
3.5. Selection Index
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tunim, S.; Phasuk, Y.; Aggrey, S.E.; Duangjinda, M. Gene expression of fatty acid binding protein genes and its relationship with fat deposition of Thai native crossbreed chickens. Anim. Biosci. 2021, 34, 751–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tunim, S.; Phasuk, Y.; Aggrey, S.E.; Duangjinda, M. Increasing fat deposition via upregulates the transcription of peroxisome proliferator-activated receptor gamma in native crossbred chickens. Animals 2021, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Charoensin, S.; Laopaiboon, B.; Boonkum, W.; Phetcharaburanin, J.; Villareal, M.O.; Isoda, H.; Duangjinda, M. Thai native chicken as a potential functional meat source rich in anserine, anserine/carnosine, and antioxidant substances. Animals 2021, 11, 902. [Google Scholar] [CrossRef]
- Grashorn, M.A. Functionality of poultry meat. J. Appl. Poult. Res. 2007, 16, 99–106. [Google Scholar] [CrossRef]
- Perić, L.; Rodić, V.; Milošević, N. Production of poultry meat and eggs as functional food—Challenges and opportunities. Biot. Anim. Husb. 2011, 27, 511–520. [Google Scholar] [CrossRef]
- Liu, X.D.; Jayasena, D.D.; Jung, Y.; Kang, B.S.; Heo, K.N.; Lee, J.H.; Jo, C. Differential proteome analysis of breast and thigh muscles between Korean native chickens and commercial broilers. Asian-Aust. J. Anim. Sci. 2012, 25, 895–902. [Google Scholar] [CrossRef]
- Yousif, I.A.; Binda, B.D.; Elamin, K.M.; Malik, H.E.E.; Babiker, M.S. Evaluation of carcass characteristics and meat quality of indigenous fowl ecotypes and exotic broiler strains raised under hot climate. Global J. Anim. Sci. Res. 2014, 2, 365–371. [Google Scholar]
- Information and Communication Technology Center. Department of Livestock Development. Available online: https://dld.go.th/th/index.php/th/ (accessed on 16 January 2021).
- Jaturasitha, S.; Srikanchai, T.; Kreuzer, M.; Wicke, M. Differences in carcass and meat characteristics between chicken indigenous to northern Thailand (black-boned and Thai native) and imported extensive breeds (Bresse and Rhode Island Red). Poult. Sci. 2008, 87, 160–169. [Google Scholar] [CrossRef]
- Adeleke, M.A.; Peters, S.O.; Ozoje, M.O.; Ikeobi, C.O.N.; Bamgbose, A.M.; Adebambo, O. Genetic parameter estimates for body weight and linear body measurements in pure and crossbred progenies of Nigerian indigenous chickens. Livest. Res. Rural. Dev. 2011, 23, 1–7. [Google Scholar]
- Padhi, M.K.; Chatterjee, R.N.; Rajkumar, U.; Bhattacharya, T.K.; Bhanja, S.K. Genetic and phenotypic parameters estimates for body weight, conformation, production and reproduction traits of PD1 (Vanaraja male line) during different periods. Indian J. Anim. Sci. 2015, 85, 883–888. [Google Scholar]
- Anh, N.T.L.; Kunhareang, S.; Duangjinda, M. Association of chicken growth hormones and insulin-like growth factor gene polymorphisms with growth performance and carcass traits in Thai broilers. Asian-Aust. J. Anim. Sci. 2015, 18, 1686–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssao, I.A.K.; Senou, M.; Dahouda, M.; Kpodékon, M.T.; Djenontin, J.; Idrissou, N.D.; Bonou, G.A.; Tougan, U.P.; Ahounou, S.; Assogba, H.M.; et al. Genetic improvement of local chickens by crossing with the label rouge (T55XSA51): Growth performances and heterosis effects. Int. J. Poult. Sci. 2009, 8, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Duangjinda, M.; Tunim, S.; Duangdaen, C.; Boonkum, W. Hsp70 genotypes and heat tolerance of commercial and native chickens reared in hot and humid conditions. Rev. Bras. Cienc. Avic. 2016, 19, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Misztal, I.; Tsuruta, S.; Lourenco, D.; Aguilar, I.; Legarra, A.; Vitezica, Z. Manual for BLUPF90 Family of Programs. Available online: Indexof/html/projects/programs/Windows/64bit(uga.edu) (accessed on 9 August 2019).
- Duangjinda, M.; Misztal, I.; Tsuruta, S. BLUPF90-ChickenPAK v 2.5 Program: User’s Manual; The University of Georgia: Athens, GA, USA; Khon Kaen University: Khon Kaen, Thailand, 2005. [Google Scholar]
- Promwatee, N.; Loapaiboon, B.; Vongpralub, T.; Phasuk, Y.; Kunhareang, S.; Boonkum, W.; Duangjinda, M. Insulin-like growth factor I gene polymorphism associated with growth and carcass traits in Thai synthetic chickens. Genet. Mol. Res. 2013, 12, 4332–4341. [Google Scholar] [CrossRef]
- Tongsiri, S.; Jeyaruban, G.M.; Hermesch, S.; van der Werf, J.H.J.; Chormai, T. Genetic parameters and inbreeding effects for production traits of Thai native chickens. Asian-Aust. J. Anim. Sci. 2019, 32, 930–938. [Google Scholar] [CrossRef]
- Molee, A.; Kuadsantia, P.; Kaewnakian, P. Gene effects on body weight, carcass yield, and meat quality of Thai indigenous chicken. J. Poult. Sci. 2018, 55, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Dana, N.; vander Waaij, E.H.; van Arendonk, J.A.M. Genetic and phenotypic parameter estimates for body weights and egg production in Horro chicken of Ethiopia. Trop. Anim. Health. Prod. 2011, 43, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Mookprom, S.; Boonkum, W.; Kunhareang, S.; Siripanya, S.; Duangjinda, M. Genetic evaluation of egg production curve in Thai native chickens by random regression and spline models. Poult. Sci. 2017, 96, 274–281. [Google Scholar] [CrossRef]
- Niknafs, S.; Nejati-Javaremi, A.; Mehrabani-Yeganeh, H.; Fatemi, S.A. Estimation of genetic parameters for body weight and egg production traits in Mazandaran native chicken. Trop. Anim. Health Prod. 2012, 44, 1437–1443. [Google Scholar] [CrossRef]
- Oleforuh-Okoleh, V.U. Estimation of genetic parameters and selection for egg production traits in a Nigerian local chicken ecotype. J. Agri. Biol. Sci. 2011, 6, 54–57. [Google Scholar]
- Han, W.; Zhu, Y.; Su, Y.; Li, G.; Qu, L.; Zhang, H.; Wang, K.; Zou, J.; Liu, H. High-throughput sequencing reveals circulating mirnas as potential biomarkers for measuring puberty onset in chicken (Gallus gallus). PLoS ONE 2016, 5, e0154958. [Google Scholar] [CrossRef] [PubMed]
- Norris, D.; Ngambi, J.W. Genetic parameter estimates for body weight in local Venda chickens. Trop. Anim. Health. Prod. 2006, 38, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Saatci, M.; Omed, H.; Dewi, I.A. Genetic parameters from univariate and bivariate analyses of egg and weight traits in Japanese quail. Poult. Sci. 2006, 85, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Manjula, P.; Park, H.B.; Seo, D.; Choi, N.; Jin, S.; Ahn, S.J.; Heo, K.N.; Kang, B.S.; Lee, J.H. Estimation of heritability and genetic correlation of body weight gain and growth curve parameters in Korean native chicken. Asian-Aust. J. Anim. Sci. 2018, 31, 26–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamali, M.A.; Ghorbani, S.H.; Sharbabak, M.M.; Zamiri, M.J. Heritabilities and genetic correlations of economic traits in Iranian native fowl and estimated genetic trend and inbreeding coefficients. Br. Poult. Sci. 2007, 48, 443–448. [Google Scholar] [CrossRef]
- Sang, B.D.; Kong, H.S.; Kim, H.K.; Choi, C.H.; Kim, S.D.; Cho, Y.M.; Sang, B.C.; Lee, J.H.; Jeon, G.J.; Lee, H.K. Estimation of genetic parameters for economic traits in Korean native chickens. Asian-Aust. J. Anim. Sci. 2006, 19, 319–323. [Google Scholar] [CrossRef]
- Zonuz, A.Y.; Alijani, S.; Mohammadi, H.; Rafat, A.; Kia, H.D. Estimation of genetic parameters for productive and reproductive traits in Esfahan native chickens. J. Livest. Sci. Tech. 2013, 1, 34–38. [Google Scholar]
- Firozjah, G.N.; Atashi, H.; Zare, A. Estimation of genetic parameters for economic traits in Mazandaran native chickens. J. Anim. Poult. Sci. 2015, 4, 20–26. [Google Scholar]
- Sungkhapreecha, P.; Duangjinda, M.; Laopaiboon, B.; Laopaiboon, T.; Boonkum, W. Comparison of growth performances in crossbred black-bone and Thai native chickens (Pradu-Hang dam KKU-55) by diallel crossing system. Khon Kaen Agr. J. 2017, 45, 333–340. [Google Scholar]
- Lwelamira, J.; Kifaro, G.C.; Gwakisa, P.S. Genetic parameters for body weights, egg traits and antibody response against Newcastle Disease Virus (NDV) vaccine among two Tanzania chicken ecotypes. Trop. Anim. Health. Prod. 2009, 41, 51–59. [Google Scholar] [CrossRef]
- Jafarnejad, A.; Kamali, M.A.; Fatemi, S.J.; Aminafshar, M. Genetic evaluation of laying traits in Iranian indigenous hens using univariate and bivariate animal models. J. Anim. Plant Sci. 2017, 27, 20–27. [Google Scholar]
- Li, H.F.; Zhu, W.Q.; Chen, K.W.; Wu, X.; Tang, Q.P.; Gao, Y.S. Associations between GHR and IGF-1 gene polymorphisms, and reproductive traits in Wenchang chickens. Turk. J. Vet. Anim. Sci. 2008, 32, 281–285. [Google Scholar]
- Feng, X.P.; Kuhnlein, U.; Fairfull, W.R.; Aggrey, S.E.; Yao, J.; Zadworny, D. A genetic marker in the growth hormone receptor gene associated with body weight in chickens. J. Hered. 1998, 89, 355–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhnlein, U.; Ni, L.; Weigend, S.; Gavora, J.S.; Fairfull, W.; Zadworny, D. DNA polymorphisms in the chicken growth hormone gene: Response to selection for disease resistance and association with egg production. Anim. Genet. 1997, 28, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Hayes, B.J.; Goddard, M.E. Technical note: Prediction of breeding values using marker-derived relationship matrices. J. Anim. Sci. 2008, 86, 2089–2092. [Google Scholar] [CrossRef] [Green Version]
- Pszczola, M.; Strabel, T.; Mulder, H.A.; Calus, M.P.L. Reliability of direct genomic values for animals with different relationships within and to the reference population. J. Diary Sci. 2012, 95, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Pal, A.; Chakravarty, A.K. Genetics and Breeding for Disease Resistance of Livestock; Chatlotte Cockle: Oxford, UK, 2020; pp. 245–258. [Google Scholar]
- Hayes, B.J.; Visscher, P.M.; Goddard, M.E. Increased accuracy of artificial selection by using the realized relationship matrix. Genet. Res. 2009, 91, 47–60. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | No. of Records | Mean | SD | Min | Max | CV (%) |
|---|---|---|---|---|---|---|
| BW0 (g) | 2713 | 37.9 | 4.2 | 26.0 | 50.0 | 11.1 |
| BW4 (g) | 2713 | 395.4 | 104.7 | 150.0 | 650.0 | 26.5 |
| BW6 (g) | 2713 | 812.3 | 138.7 | 457.0 | 1400.0 | 17.1 |
| BW8 (g) | 2713 | 1228.8 | 220.2 | 700.0 | 1895.0 | 17.9 |
| BW10 (g) | 2713 | 1642.4 | 261.4 | 900.0 | 2450.0 | 15.9 |
| BrC6 (cm) | 2713 | 27.6 | 2.3 | 20.3 | 36.0 | 8.3 |
| AFE (day) | 2713 | 168.0 | 21.0 | 120.0 | 199.0 | 12.5 |
| 240EP (egg) | 2710 | 43.0 | 19.4 | 12.0 | 96.0 | 45.1 |
| 270EP (egg) | 2710 | 55.0 | 24.2 | 15.0 | 123.0 | 44.0 |
| 300EP (egg) | 2708 | 65.0 | 28.1 | 23.0 | 146.0 | 43.2 |
| 365EP (egg) | 2700 | 84.0 | 28.0 | 44.0 | 151.0 | 33.3 |
| Traits/Parameters | ||||
|---|---|---|---|---|
| BW0 | 8.77 | 4.89 | 13.66 | 0.642 (0.10) |
| BW4 | 1412 | 2835 | 4247 | 0.332 (0.05) |
| BW6 | 5782 | 7687 | 13,469 | 0.429 (0.04) |
| BW8 | 11,485 | 21,059 | 32,544 | 0.353 (0.04) |
| BW10 | 12,602 | 32,364 | 44,966 | 0.280 (0.02) |
| BrC6 | 1.10 | 2.77 | 3.87 | 0.284 (0.03) |
| AFE | 86.50 | 234.80 | 321.30 | 0.269 (0.03) |
| 240EP | 138.10 | 185.40 | 323.50 | 0.427 (0.03) |
| 270EP | 2.50 | 330.20 | 552.70 | 0.403 (0.02) |
| 300EP | 303.40 | 448.20 | 751.60 | 0.404 (0.02) |
| 365EP | 489.20 | 660.40 | 1149.60 | 0.426 (0.02) |
| Traits | BW0 | BW4 | BW6 | BW8 | BW10 | BrC6 | AFE | 240EP | 270EP | 300EP | 365EP |
|---|---|---|---|---|---|---|---|---|---|---|---|
| BW0 | - | 0.22 | 0.15 | 0.10 | 0.08 | 0.12 | −0.03 | −0.15 | −0.03 | −0.28 | −0.29 |
| BW4 | 0.16 | - | 0.61 * | 0.51 * | 0.40 * | 0.30 * | −0.10 * | −0.24 * | −0.32 * | −0.43 * | −0.48 * |
| BW6 | 0.14 | 0.58 * | - | 0.91 * | 0.79 * | 0.80 * | −0.18 * | 0.08 * | −0.46 * | −0.49 * | −0.40 * |
| BW8 | 0.10 | 0.48 * | 0.85 * | - | 0.86 * | 0.84 * | −0.20 * | 0.14 * | −0.40 * | −0.44 * | −0.32 * |
| BW10 | 0.06 | 0.36 * | 0.72 * | 0.82 * | - | 0.93 * | −0.25 * | 0.15 * | −0.38 * | −0.42 * | −0.35 * |
| BrC6 | 0.12 | 0.24 * | 0.72 * | 0.80 * | 0.90 * | - | −0.27 * | 0.08 * | −0.32 * | −0.35 * | −0.30 * |
| AFE | 0.01 | −0.08 * | −0.16 * | −0.21 * | −0.22 * | −0.24 * | - | −0.14 * | −0.29 * | −0.21 * | −0.16 * |
| 240EP | −0.08 | −0.14 * | −0.01 * | 0.10 * | 0.12 * | 0.07 * | −0.14 * | - | 0.34 * | 0.22 * | 0.04 * |
| 270EP | −0.11 | −0.25 * | −0.40 * | −0.38 * | −0.36 * | −0.33 * | −0.23 * | 0.33 * | - | 0.60 * | 0.40 * |
| 300EP | −0.20 | −0.40 * | −0.42 * | −0.40 * | −0.40 * | −0.28 * | −0.18 * | 0.19 * | 0.54 * | - | 0.86 * |
| 365EP | −0.24 | −0.42 * | −0.38 * | −0.28 * | −0.32 * | −0.26 * | −0.12 * | 0.09 * | 0.38 * | 0.85 * | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chomchuen, K.; Tuntiyasawasdikul, V.; Chankitisakul, V.; Boonkum, W. Genetic Evaluation of Body Weights and Egg Production Traits Using a Multi-Trait Animal Model and Selection Index in Thai Native Synthetic Chickens (Kaimook e-san2). Animals 2022, 12, 335. https://doi.org/10.3390/ani12030335
Chomchuen K, Tuntiyasawasdikul V, Chankitisakul V, Boonkum W. Genetic Evaluation of Body Weights and Egg Production Traits Using a Multi-Trait Animal Model and Selection Index in Thai Native Synthetic Chickens (Kaimook e-san2). Animals. 2022; 12(3):335. https://doi.org/10.3390/ani12030335
Chicago/Turabian StyleChomchuen, Kitsadee, Veeraya Tuntiyasawasdikul, Vibuntita Chankitisakul, and Wuttigrai Boonkum. 2022. "Genetic Evaluation of Body Weights and Egg Production Traits Using a Multi-Trait Animal Model and Selection Index in Thai Native Synthetic Chickens (Kaimook e-san2)" Animals 12, no. 3: 335. https://doi.org/10.3390/ani12030335
APA StyleChomchuen, K., Tuntiyasawasdikul, V., Chankitisakul, V., & Boonkum, W. (2022). Genetic Evaluation of Body Weights and Egg Production Traits Using a Multi-Trait Animal Model and Selection Index in Thai Native Synthetic Chickens (Kaimook e-san2). Animals, 12(3), 335. https://doi.org/10.3390/ani12030335